
miR-125a/TRAF6调控通路在破骨细胞分化中的 作用研究
2014-08-10 12:28杨丽赵新兰雷丹丹廖斌秦爱平
疑难病杂志 2014年11期
杨丽,赵新兰,雷丹丹,廖斌,秦爱平
论著·基础
miR-125a/TRAF6调控通路在破骨细胞分化中的 作用研究
杨丽,赵新兰,雷丹丹,廖斌,秦爱平
目的研究miR-125a在破骨细胞分化过程中的作用及其作用机制,方法单核细胞集落刺激因子(M-CSF)和核因子-κB配体的受体激活剂(RANKL)诱导CD+14PBMCs向破骨细胞分化。生物信息学运算预测miR-125a的靶基因以及结合于miR-125a启动子区域的转录因子。实时定量PCR法检测miR-125a以及其他相关基因的表达。过表达实验和抑制试验用于研究miR-125a在破骨细胞分化中的作用及其与靶基因之间的相互关系。荧光素酶报告基因实验验证miR-125a与其靶基因之间的结合。结果(1)miR-125a表达在CD+14PBMCs前体细胞高表达,随着诱导时间的延续逐渐下降,在诱导第5天开始明显下降并在第15天达到最低值,表明miR-125a的表达在破骨细胞分化过程中呈现明显下降趋势。(2)miR-125a参与调控RANKL和M-CSF诱导的破骨细胞分化,过表达miR-125a明显抑制了TRAP和NFATc1的mRNA表达和CD+14PBMCs向破骨细胞的分化。(3)miR-125a直接作用于靶基因肿瘤坏死因子相关因子6 (TRAF6),TRAF6是RANKL/RANK/NFATc1信号转导通路的转录因子,过表达miR-125a降低了TRAF6的蛋白表达水平,而TRAF6的mRNA水平无明显改变。结论miR-125a通过作用于新的TRAF6/NFATC1/miR-125a负反馈调控环路在破骨细胞的分化中发挥了重要的调控作用,调控miR-125a的表达可能成为骨代谢疾病新的治疗靶点。
microRNA; 破骨细胞分化; 转录因子;靶基因;负反馈调节

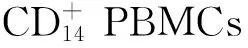
1 材料与方法
1.1 材料
1.1.2 主要仪器: Nikon 300型自动摄像倒置显微镜(日本Nikon公司生产),电泳仪、转印仪、紫外交联仪、凝胶成像系统(美国Bio.Rad公司生产),流式细胞仪(美国Coulter公司)。
1.1.3 主要试剂: 无酚红a-MEM培养液、Trizol、及胎牛血清(FBS)、0.25%胰酶-EDTA、胶原酶(美国Gibico公司生产),RANKL(美国Invitrogen公司生产),M-CSF(美国Sigma公司),TRAP染色试剂盒(美国Sigma公司), 染色质免疫沉淀实验试剂盒(Upstate, Charlottesville, VA), anti-miR-125a(GenePharma Co., Ltd), NFATc1 siRNA(OriGene Technologies Inc)。
1.2 方法
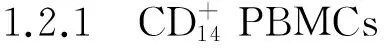
诱导液(25 ng/ml重组人类巨噬细胞集落刺激因子+30 ng/ml RANKL)诱导破骨细胞分化。抗酒石酸酸性磷酸酶(TRAP)染色法鉴定破骨细胞的成熟,同时检测组织蛋白酶免疫染色并检测破骨细胞分化标志性基因,如RAP和活化T细胞核因子磷酸酶依赖因子1的mRNA的表达水平。
1.2.2 实时定量PCR分析: 利用罗氏实时荧光定量PCR仪,方法按参考文献[10]。 miR-125a、U6、TRAF6、NFATc1、TRAP和β-actin的引物见表1、表2, U6和β-actin作为内参。

表1 mRNA实时定量RT-PCR引物序列

表2 mRNA实时定量RT-PCR引物序列
注: F. 正向引物; R.反向引物; Acc. No. Genbank公布的准入号
1.2.3 TRAP染色: 利用TRAP染色试剂盒进行染色。在显微光镜下(× 200)随机挑选10个视野观察TRAP染色阳性的细胞(细胞核≥3),每个视野计数重复3次,取平均值。
1.2.4 免疫组化: PBMCs的培养方法同前。多聚甲醛固定诱导液下培养于二孔板中的破骨细胞。破骨细胞免疫染色方法按参考文献 [11]。
1.2.5 miR-125a靶基因预测: 目前有大量miRNA靶基因预测软件,其各有特点。TargetScan软件是一个常用的预测软件。在本研究中利用TargetScan预测并使用RNA22-HSA进行验证。
1.2.6 质粒构建与转染: 质粒构建方法按参考文献[12],构建包含所预测miR-125a结合位点的WT-pGL3-TRAF6-3′UTR。引物如下,F:5′-GGCTCTAGACTAATAGGGAGATGATTT-3′,R:5′-GGCCGGCCGCATTACCAGACAACTTTA-3′。MUT-pGL3-TRAF6-3′UTR. 突变引物如下,F:5′-TATCGTGGAATCTAGTTCTCCG CGAGACCCGCAACTAGTATAAG-3′,R:5′-GCTTATACTAGTTGCGGGTCTCGCGGAGAACTAGATTCCA CGATA-3′miR-125a 特异性抑制剂2′-O-甲基反义寡核苷酸(anti-miR-125a)和脂质体复合体严格按说明书直接和细胞混合。
1.2.7 荧光素酶报告基因实验: 外周血单核细胞与野生型或突变型 pGL3-TRAF6-3′UTR和miR-125a 或 miR-C共培养48 h。空脂质体作为阴性对照。
双荧光素酶报告基因系统(Promega)和光度计用以荧光信号定量。每个荧光素酶值通过海肾荧光素酶值进行校正。

2 结 果


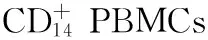
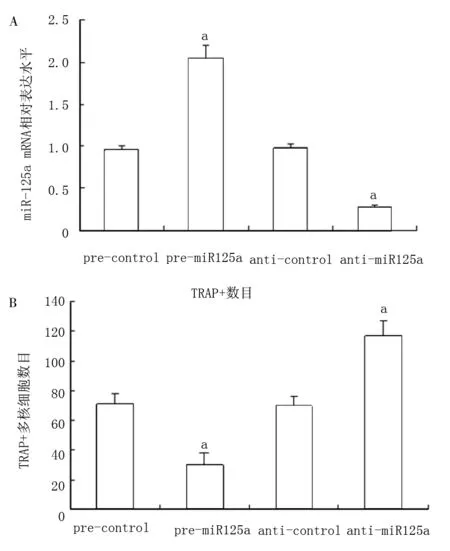



图3 miR-125a抑制破骨细胞分化的标志性基因NFATC1的表达
2.3 miR-125a直接作用于靶基因肿瘤坏死因子相关因子6 (TRAF6) 利用TargetScan软件预测miR-125a的靶基因。在TRAF6的3′UTR区存在miR-125a的碱基互补序列(图4A)。TRAF6是破骨细胞分化中的重要调控因子。
我们检测了调控破骨细胞分化的一些重要基因如c-Fos、Mitf和NF-κB基因在转染pre-miR-125a或miR-C以及anti-miR-125a或anti-C前后的表达情况。结果示,miR-125对c-Fos和Mitf基因的表达没有影响,过表达miR-125a促进NF-κB的蛋白表达,反之则抑制其表达(图4E)。而NF-κB不是miR-125a的靶基因。
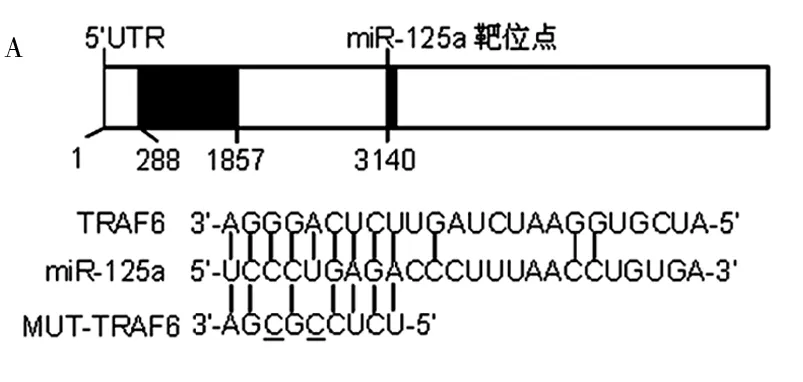



注:A.图示标明了在TRAF6的3’UTR端存在有miR-125a的结合位点。并且标注了与miR-125a结合序列结合的野生型(WT)TRAF6- 3’UTR以及突变型(MUT)TRAF6- 3’UTR
B,C.miR-125a在转录后水平抑制TRAF6的表达。过表达或抑制miR-125a的表达可以降低或提高TRAF6的蛋白表达水平而不影响其mRNA表达水平。与对照组比较,aP<0.05
D.荧光素酶报告基因实验示:共转染pre-miR-125a可以降低野生型(WT)TRAF6- 3’UTR的荧光素酶活性而不影响突变型(MUT)TRAF6- 3’UTR的荧光素酶活性。与对照组比较,aP<0.05
E.过表达或下调miR-125a的表达可抑制或升高NF-κB蛋白水平却对破骨细胞分化相关的其他一些关键基因无直接作用
图4 miR-125a直接作用于其靶基因TRAF6
3 讨 论
本研究发现过表达miR-125a抑制了TRAP+多核巨细胞的生成同时也抑制了TRAP 和NFATc1的基因表达。反之则结果相反。表明miR-125a在破骨细胞的分化过程中起到了重要的调控作用。
研究已发现miRNAs通过与靶基因的3’UTR结合来抑制靶基因的转录或降低靶基因mRNA的稳定性。我们利用TargetScan软件预测发现RANKL信号转导因子肿瘤坏死因子相关因子6 (TRAF6)是我们所预测的靶基因之一。结果提示miR-125a通过与TRAF6 mRNA的3’UTR结合来调控破骨细胞的分化。
TRAF6是NF-κB受体活化因子(RANK)的重要的衔接分子,在破骨细胞膜与RANK结合并激发了TRAF6的募集并进一步激活NF- κB和NFATc1促使破骨细胞的形成[15,16]。TRAF6是TRAF超家族和白介素-1/Toll样受体超家族的重要信号转导因子。研究报道TRAF6和RANK的耦联对于破骨细胞的骨吸收功能的维持至关重要。因此,TRAF6在破骨细胞分化和成熟过程中其重要的调控作用[17]。
本研究证实miR-125a通过作用于靶基因TRAF6调控破骨细胞分化。荧光素酶报告基因实验显示过表达miR-125a可抑制野生型TRAF6荧光素酶报告基因活性,而突变型TRAF6荧光素酶报告基因载体可取消该作用。另外,过表达和抑制试验表明miR-125a在转录后水平抑制TRAF6的表达水平。而对其他一些可能对破骨细胞分化起调控作用的基因例如:c-Fos、小眼畸形相关转录因子(Mitf)和NF-κB的表达没有影响。这些结果提示miR-125a在转录后水平抑制TRAF6基因的表达。
本研究数据可以得出miR-125a的作用模式。破骨细胞分化信号在破骨细胞前体细胞激活TRAF6/NFATc1转导通路,miR-125a表达的下降维持了TRAF6的蛋白表达并维持了破骨细胞分化过程。因此,一个新的miR-125a/TRAF6调控通路被挖掘出来。
综上所述,本研究证实了miR-125a通过一个新发现的调控通路参与破骨细胞的分化调节。我们的研究为miRNA在破骨细胞分化中的作用提供了新的认识,揭示了miR-125a在破骨细胞分化过程中的重要作用并使miR-125a表达的调控,可能成为治疗骨代谢疾病的新的手段。
1 Taylor A,Rogers MJ,Tosh D,et al.A novel method for efficient Generation of transfected human osteoclasts[J].Calcif Tissue Int,2007,80(2):132-136.
2 Graves P1,Zeng Y. Biogenesis of mammalian microRNAs: a global view Genomics[J].Proteomics Bioinformatics,2012,10(5):239-245.
3 Ambros V.The functions of animal microRNAs[J].Nature,2004,431:350-355.
4 Kim SW,Ramasamy K,Bouamar H,et al.MicroRNAs miR-125a and miR-125b constitutively activate the NF-κB pathway by targeting the tumor necrosis factor alpha-induced protein 3 (TNFAIP3, A20)[J].Proc Natl Acad Sci U S A,2012,109(20):7865-7870.
5 Li Z,Hassan MQ,Volinia S,et al.A microRNA signature for a BMP2-induced osteoblast lineage commitment program[J].Proc Natl Acad Sci U S A,2008,105(37):13906-13911.
6 Mizuno Y,Yagi K,Tokuzawa Y,et al.miR-125b inhibits osteoblastic differentiation by down-regulation of cell proliferation[J].Biochem Biophys Res Commun,2008,368(2):267-272.
7 Hu R,Liu W,Li H,et al.A Runx2/miR-3960/miR-2861 regulatory feedback loop during mouse osteoblast differentiation[J].J Biol Chem,2011,286(14):12328-12339.
8 Yang L,Cheng P,Chen C,et al.miR-93/Sp7 function loop mediates osteoblast mineralization[J].J Bone Miner Res,2012,27(7):1598-1606.
9 Feige U,Overwien B,Sorg C.Purificmion of human blood monocytes by hypotonic density gradient centrifugation in Percoll[J].J Immunol Methods,1982,54(3):309-315.
10 Li H,Xie H,Liu W,et al.A novel microRNA targeting HDAC5 regulates osteoblast differentiation in mice and contributes to primary osteoporosis in humans[J].J Clin Invest,2009,119(12):3666-3677.
11 Wang L,Zhao R,Shi X,et al.Substance P stimulates bone marrow stromal cell osteogenic activity, osteoclast differentiation, and resorption activity in vitro[J].Bone,2009,45(2):309-320.
12 Nakamura T,Scorilas A,Stephan C,et al.Quantitative analysis of macrophage inhibitory cytokine-1(MIC-1)gene expression in human prostatic tissues[J].Br J cancer,2003,88(7):1101-1104.
13 Marson A,Levine SS,Cole MF,et al.Connecting microRNA genes to the core transcriptional regulatory circuitry of embryonic stem cells[J].Cell,2008,734(3):521-533.
14 Cheng P,Chen C,He HB,et al.miR-148a regulates osteoclastogenesis by targeting v-maf musculoaponeurotic fibrosarcoma oncogene homolog B[J].J Bone Miner Res,2013,28(5):1180-1190.
15 Zhou XY,Zhou ZG,Ding JL,et al.TRAF6 as the key adaptor of TLR4 signaling pathway is involved in acute pancreatitis[J].Pancreas,2010,39(3):359-366.
16 Chen H,Wu Y,Zhang Y,et al.Hsp70 inhibits lipopolysaccharide-induced NF-kappaB activation by interacting with TRAF6 and inhibiting its ubiquitination[J].FEBS Lett,2006,580(13):3145-3152.
17 Chen H,Li M,Campbell RA,et al.Interference with nuclear factor kappa B and c-Jun NH2-terminal kinase signaling by TRAF6C small interfering RNA inhibits myeloma cell proliferation and enhances apoptosis[J].Oncogene,2006,25(49):6520-6527.
EffectofmiR-125a/TRAF6/NFATC1loopmediatesonosteoclastogenesis
YANGLi*,ZHAOXinlan,LEIDandan,LIAOBin,QINAiping.
*DepartmentofEndocrinology,HunanProvinceMawangduiGeriatricHospital,Changsha410016,China
QINAiping,E-mail:qinaip@163.com
ObjectiveTo investigate the effect of miR-125a in the process of osteoclast.MethodsMonocyte colony stimulating factor (M-CSF) and nuclear factor kappa B ligand / receptor activator (RANKL) induced CD+14PBMCs to osteoclast. The target gene bioinformatics arithmetic predictive miR-125a and miR-125a binding to initiate transcription factor sub region. To detect the expression of miR-125a using real-time quantitative PCR and other related genes. Overexpression experiments and inhibition test were used to investigate the relationship between effect of miR-125a in osteoclast differentiation and target gene. Used the luciferase reporter gene experiments to verify the combination between miR-125a and its target gene.Results(1) miR-125a expression in CD+14PBMCs precursor cells showed high expression level, with a continuation of the induction time, the expression decreased gradually, and reached the lowest value at fifteenth days, which showed that the expression of miR-125a revealed a clear downward trend in osteoclast differentiation process. (2) miR-125a are involved in the regulation of RANKL and M / CSF induced osteoclast differentiation, over expression of miR-125a significantly inhibited the expression of mRNA and CD+14PBMCs TRAP and NFATc1 to differentiate into osteoclasts. (3) miR-125a direct effect on target gene expression of tumor necrosis factor related factor 6 (TRAF6), TRAF6 is a transcription factor of the RANKL/RANK/NFATc1 signaling pathway, overexpression of miR-125a decreased the protein expression level of TRAF6, but no significant change of TRAF6 mRNA level were found.ConclusionIt proved that the miR-125a played an important role in the regulation of osteoclast differentiation by acting on the new TRAF6/NFATC1/miR-125a negative feedback control loop, expression and
regulation of miR-125a may become a new therapeutic target for metabolic bone diseases.
microRNA; Osteoclast; Transcription factor; Target gene; Negative feedback
湖南省自然科学基金(No.13JJ4119)
410016 长沙,湖南省马王堆医院老年内分泌科(杨丽、赵新兰、廖斌、秦爱平); 410010 中南大学湘雅二医院 内分泌科(雷丹丹)
秦爱平,E-mail:qinaip@163.com
10.3969 / j.issn.1671-6450.2014.11.019
2014-08-12)
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21
中日友好医院学报(2021年1期)2021-04-14
山东医药(2020年36期)2020-12-31
山东医药(2020年9期)2020-05-20
中国临床医学(2019年3期)2019-01-04
中国骨质疏松杂志(2016年1期)2016-01-29
中国药理学通报(2015年2期)2016-01-12
热带农业科学(2015年9期)2015-10-14
中国医学科学院学报(2015年5期)2015-03-01
中国医药导报(2015年27期)2015-02-28
