植物初级三体的产生途径、鉴定方法及在遗传学研究中的应用概述
2019-01-22 03:59祝海燕
生物学教学 2019年1期
祝海燕
(山东潍坊科技学院/山东省高校设施园艺实验室 寿光 262700)
自然界中正常植物都是整倍体植物(2n),其体细胞中染色体组(x)是整倍存在的,如二倍体植物(2n=2x)、三倍体植物(2n=3x)及四倍体植物(2n=4x)等。而植物三体(2n+1)是指在正常植物(2n)的染色体组中多一条额外染色体的植物个体,属非整倍体植物。由于额外染色体的性质不同,植物三体又可分为初级三体、次级三体、三级三体和端三体四种类型,其中以初级三体应用最广。
植物初级三体是重要的遗传工具材料。利用初级三体可进行目标染色体的分离,可将基因、连锁群等定位到特定染色体上,从而构建基因的物理图谱。因此,对植物初级三体的创建和研究国内外都十分重视。本文就植物三体的概念、类型,初级三体产生途径、鉴定方法及在遗传学研究中的应用做一概述。
1 植物三体的类型
根据额外染色体的来源和性质,可将三体分为以下四种类型:
1.1 初级三体 在正常(2n)染色体组基础上,某对同源染色体增加了一条额外的完整的染色体(图1)。
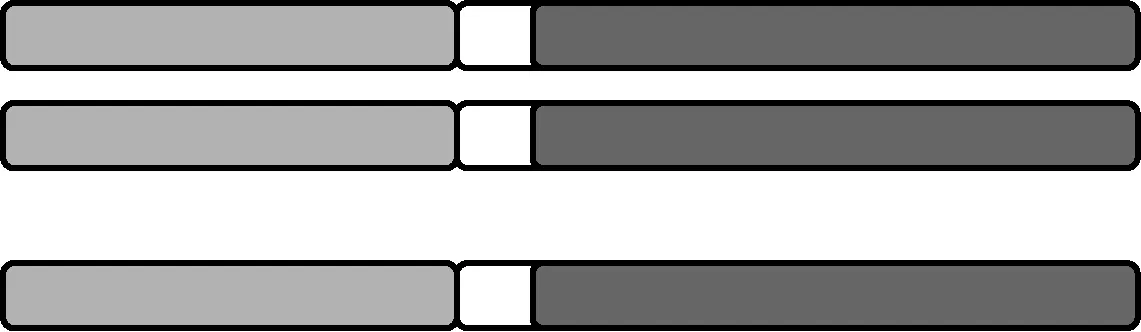
图1 初级三体示意图
1.2 次级三体 在正常(2n)染色体组基础上,某对同源染色体增加了一条由自己一条染色体臂构成的等臂染色体(图2)。
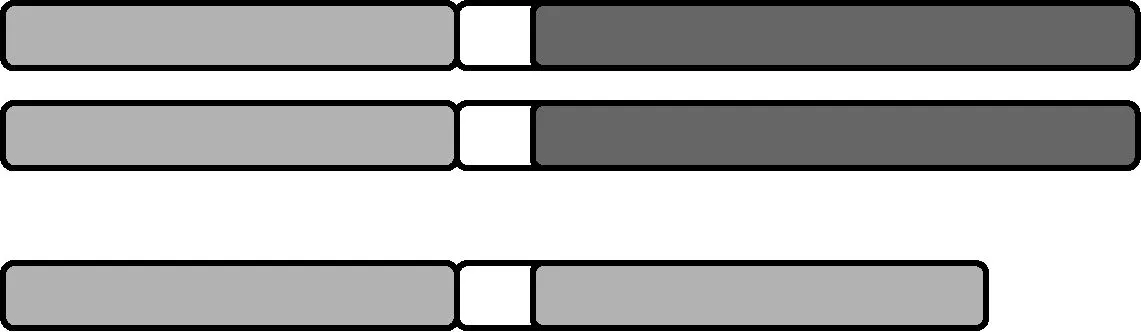
图2 次级三体示意图
1.3 三级三体 在正常(2n)染色体组基础上,某对同源染色体增加了一条自己的易位染色体(图3)。
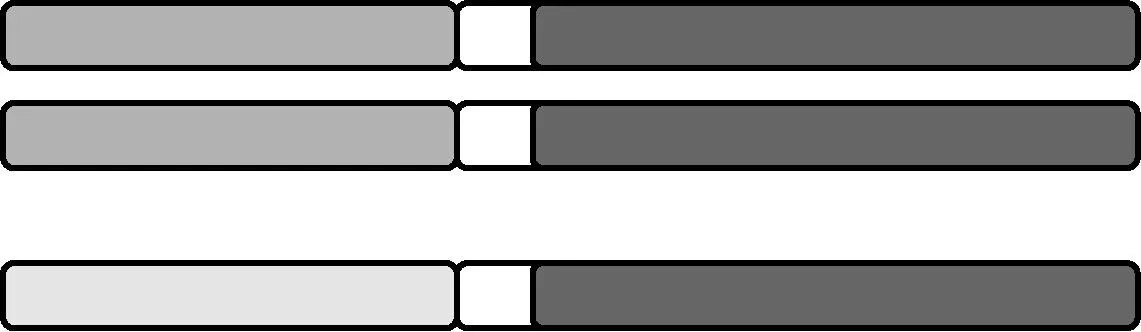
图3 三级三体示意图
1.4 端三体 在正常(2n)染色体组基础上,某对同源染色体增加了一条端着丝点染色体(图4)。
图4 端三体示意图
在植物的四种三体类型中,初级三体是最常见、研究最多的,在植物遗传学研究中应用最广的三体类型。
2 植物初级三体产生的途径
2.1 自然发生 三体植物在自然界可自然产生,如Blakeslee最早从曼陀罗中分离出的全部12个染色体的初级三体都是自然产生的。三体植株的自然发生主要来源于减数分裂中的错分离,倘若某对同源染色体在后期Ⅰ不分离,其结果就会产生n+1和n-1两种配子,进而与正常(n)配子受精结合产生三体和单体植株。但这种不分离现象并不常见,自然发生的频率较低,平均不到千分之一。因此,依靠自然途径获得一套初级三体是十分困难的。
2.2 人工创建 目前,植物的三体系一般都是通过人工选育获得。主要是通过三倍体自交或回交分离非整倍体植株获得,通过组织培养和物理途径也可获得一些三体。
2.2.1 杂交途径 利用同源三倍体减数分裂染色体的无规律分配,从3x×2x或2x×3x的杂交子代中筛选三体植株,是人工创建植物三体系的主要途径。迄今为止,一些重要农作物(如番茄、玉米、水稻、甜菜、大麦、油菜和谷子等)的初级三体系均是通过该途径创建而成的。该途径的优点是适用性比较广泛、可靠性高,一般能获得三倍体的植物,均能筛选出初级三体系。
2.2.2 射线诱发 利用各种射线(如χ和γ射线)诱发,使染色体分裂异常,从而提高三体等非整倍体的发生频率,如胡保民等[1]用γ射线处理陆地棉种子,其自交后代群体内的非整倍体频率显著提高,并从中分离出了3个三体和2个四体。
2.2.3 同源四倍体小孢子培养 刘宗贤等[2]从同源四倍体水稻的花粉植株中鉴定出了三体植株。申书兴等[3]利用同源四倍体大白菜游离小孢子培养获得了139个非整倍体小孢子植株系,初步建成了全套的白菜初级三体系,为植物初级三体的创建开辟了一条新途径。
3 植物初级三体的鉴定
3.1 形态学方法 植物的初级三体由于是在植物正常染色体组(2n)的基础上额外增加了一条正常的染色体,从而使植物所涉及的相关染色体上的基因剂量由原来的2份增加为3份。这种附加效应,使得植物初级三体植株在形态上与正常植株相比表现出一定的差异。例如,直果曼陀罗的各三体植株与正常植株相比其球形朔果出现不同程度的形态变异。另外,水稻、玉米、番茄和甘蓝等植物部分三体植株在形态学上均有不同程度的差异。但通过形态学差异鉴定植物三体准确性不高,只能作为初步筛选的依据。
3.2 有丝分裂染色体核型分析 对生物细胞核内全部染色体的形态特征(如染色体的数目、形态、长度、带型及着丝粒位置等)所进行的分析称为染色体的核型分析。不同物种的染色体组具有不同的核型,且具有很高的稳定性和再现性。因此,通过植物染色体核型分析可对植物初级三体进行鉴定。张成合等[4]通过染色体核型分析,鉴定了甘蓝的全套初级三体。但染色体核型分析鉴定植物初级三体对染色体数目较少,或染色体较大且细胞内染色体差别较大的作物准确性较高,而对染色体较小且染色体差别较小的作物用核型分析,就不能准确鉴定,故需采用其他方法鉴定。
3.3 染色体分带核型分析 染色体分带技术是通过一定的碱、酸、盐和温度等因素对染色体进行处理,借助Giemsa等染料染色,使染色体在一定部位呈现深浅程度不同的染色区段而形成特定的带纹,由于其带纹在各染色体上显示的位置、宽窄、大小、数目、浓淡等具有相对的稳定性,故可用来进行染色体的鉴别和分析。姚青等[5]利用染色体分带G带技术,成功地进行了水稻三体的鉴定。盛茂银[6]利用染色体分带C带技术对甜荞整套三体材料进行了鉴定。随着染色体分带技术的不断发展,染色体分带核型分析已成为三体鉴定的一个重要方法。
3.4 荧光原位杂交技术鉴定 20世纪90年代以后,原位杂交技术等现代手段使染色体显带方法又有进一步的改进和完善,形成了荧光原位杂交显带技术,已经被用于染色体的鉴别和分析。轩淑欣等[7]以5s rDNA为探针,用荧光原位杂交技术对大白菜3个三体材料额外染色体的归属进行了鉴定。此外,有研究用染色体分带C-带带型分析技术和荧光原位杂交技术对普通荞麦的12个三体进行了鉴定。
4 植物初级三体在植物遗传学研究中的应用
4.1 利用三体分离特点进行标志基因定位 由于三体自交后代的分离比例与二倍体不同,并且三体由父母方传递频率不同,可对三体由父母方进行传递的总体平均数加以修正,从而以F2分离比例为标准判定基因的染色体归属。例如,Dong YJ等[8]利用三体将水稻的香味基因定位在水稻的第8条染色体上。矮杆基因、光敏核不育基因也相继被定位到水稻的染色体上。
4.2 对三体植物进行同功酶分析,把同功酶基因定位到特定染色体上 用“三体系”的每个三体为母本,分别与2n个体杂交,选杂种F1的三体进行酶谱分析。如果F1表达了双体的酶谱型,即呈现了相对染色强度1∶1两条酶带(单体酶),表明该同功酶基因与测定的三体无关。如果F1表达了相对染色强度为2∶1的两条酶带(单体酶),即呈现了三体酶谱型,表明该同功酶基因与三体有关。WuKS等[9]利用水稻三体的酶谱分析,成功地把10个同功酶基因分别定位到不同的染色体上。此外,利用三体分析法,对玉米、番茄和大麦等植物的某些同功酶基因也成功进行了染色体定位。
4.3 利用分子杂交技术,对连锁群进行染色体定位 由于初级三体增加一条同源的额外染色体,位于该染色体上的DNA含量亦以等比例增加,使得位于所涉及染色体上的DNA剂量也由原来的2份增加至3份。因此通过制作不同初级三体的等量DNA膜,并以待测DNA序列作探针进行Southern杂交,其杂交信号的强度与基因组中标记DNA的剂量成正比。所以,根据杂交信号的强弱便可测知这些序列所在的染色体。Young等[10]用这一原理通过制备番茄不同初级三体等量DNA膜,并与基因组克隆进行Southern杂交,从而将52个克隆定位在不同染色体上。
4.4 利用初级三体进行目标染色体的分离 利用初级三体减数分裂终变期及中期Ⅰ染色体分裂行为的特点,对于一些染色体较小,通过核型分析较难将各条染色体区分开的作物,可较准确地辨别目标染色体从而实现目标染色体的分离。魏建华等[11]以粳稻第12号初级三体为材料,采用玻璃针染色体微分离法,成功地分离到了水稻第12号染色体,为水稻特定染色体的微分离、体外扩增找到了新途径,也为以后的水稻目标染色体特异性文库的建立奠定了基础。
4.5 以三体为母本与未知易位纯合体杂交来鉴别易位所涉及的染色体 以各初级三体为母本分别与未知易位纯合体杂交,然后对杂交子代植株的减数分裂行为进行显微观察,如果下代植株染色体在减数分裂前期Ⅰ同源染色体联会时出现有一个四体环和一个三价体或一个四体环和一个二价体加单价体,则该三体染色体不涉及易位染色体。如果下代染色体联会时出现一个五价体(环或链),表明易位染色体与该三体染色体有关。
猜你喜欢
现代妇产科进展(2022年9期)2022-10-07
基层中医药(2022年4期)2022-07-22
云南医药(2021年6期)2022-01-08
临床检验杂志(2021年10期)2021-11-24
汉字汉语研究(2021年2期)2021-08-30
川北医学院学报(2021年6期)2021-07-13
中国产前诊断杂志(电子版)(2021年1期)2021-04-23
实用临床医药杂志(2020年1期)2020-03-13
中国妇幼健康研究(2019年12期)2019-12-31
汉字汉语研究(2019年2期)2019-08-27