三七总皂苷在氧化应激的PC12细胞中抑制Aβ蛋白生成的作用
2014-04-02 00:33廖冬燕肖颖梅屈泽强
中成药 2014年4期
廖冬燕, 肖颖梅, 屈泽强, 夏 星*
(1.广州中医药大学中药学院,广东 广州 510405;2.广西中医药大学药学院,广西 南宁 530001;3.广西中医药大学基础医学院,广西 南宁 530001)
阿尔茨海默氏病(Alzheimer’s disease)为伴随着机体老龄化而出现的神经退行性病变,其发病率随年龄增长呈显著增加,Aβ蛋白(amyloid β peptide,Aβ)的大量积累导致大脑老年斑的形成及大量的神经元损失是阿尔茨海默氏病的关键病理特征[1]。氧化应激常在生命活动的能量代谢过程中出现,当机体的抗氧化系统不能及时清除能量代谢所产生的大量氧自由基和活性氧,就会造成质膜细胞器及细胞中的各种蛋白质和核酸受到氧化损伤,引发氧化应激,此外各种环境污染物、紫外线照射等也导致氧自由基大量增加而引发氧化应激[2]。老龄机体内氧化应激造成的损伤也随年龄呈进行性增强,这也是引发阿尔茨海默氏病患者神经退行性病变的重要原因。三七Panaxnotoginseng(Burk.)F.H.Chen为我国常用中药,有散瘀止血,消肿定痛的功效,中医常于止血、补血及治疗各种血瘀证。三七总皂苷(Panaxnotoginsengsaponins, PNS)为三七中主要的活性成分,当前临床上常用于脑血管疾病后遗症及各种血栓性疾病的治疗。经多方研究发现,PNS对痴呆症有明显的治疗作用[3-4]。本课题组前期研究也已证实,PNS能显著缓解体外培养的PC12细胞的氧化应激程度,增强细胞的抗氧化能力[5]。本研究进一步关注于PNS对氧化应激的PC12细胞中Aβ生成的影响,为阐明其治疗阿尔茨海默氏病的作用机制提供实验依据。
1 实验材料
1.1 细胞 大鼠肾上腺髓质嗜铬细胞瘤细胞株PC12细胞购买于北京北纳创联生物技术研究院。
1.2 实验试剂 三七总皂苷由云南玉溪维和制药厂生产(批号 20100701)。DMEM/high glucose购买于赛默飞世尔生物化学制品有限公司(批号NXBO566);无支原体胎牛血清购买于杭州四季清生物工程材料有限公司(批号090603); 淀粉样前体蛋白(APP,批号 201205),α-分泌酶(批号 201205),β-分泌酶(批号 201205)及γ-分泌酶(批号 201205)的ELISA检测试剂盒均为Raybio 公司产品(进口分装);Western blot用一抗APP(批号 297543),早老素-1(PS-1,批号 Y-AB3-07E02A)均购买武汉博士德生物工程有限公司。Western blot所用其它试剂(批号 20120228)均购于碧云天生物技术研究所。Taq酶(批号 20111201)为BioFlux公司产品;DNA ladder(批号25911J)为北京康为世纪生物科技有限公司产品;TIANScript cDNA 第一链合成试剂盒(批号 K0208),dNTP(批号K0103)为天根生化科技(北京)有限公司产品;琼脂糖(批号 111860)为西班牙BIOWEST产品;PCR引物由生工生物工程(上海)股份有限公司合成。
1.3 仪器 DLE560超净工作台(荷兰clearAir公司);311型二氧化碳培养箱(美国Thermo Forma);Sorvall ST16R超低温离心机(Thermo Fisher);DML 3000B倒置荧光显微镜(莱卡仪器有限公司);EPOCH 酶标仪(美国BIOTEK公司),JY92超声波粉碎机(宁波新芝生物科技股份有限公司);Criterion蛋白印迹系统及Universal通用型电源,均为美国Bio-rad产品;Gel Logic 212 PRO凝胶成像系统及 Image station system 4000MM分子影像系统,均为美国Carestream产品。
2 实验方法
2.1 PC12细胞培养 用含10%胎牛血清和5%马血清的DMEM培养液, 37 ℃、5% CO2条件下培养细胞,每2天传代1次。待细胞增长至80%融合时,用0.25%胰酶消化细胞,调整细胞数至1×105个/mL后传代或接种于细胞培养板。前期实验已经确定PNS增强细胞抗氧化能力的最佳浓度,本研究即在该浓度下考察PNS对细胞Aβ生成的影响,故实验分别设正常组(不加药物正常培养的细胞),模型组(不经药物处理,用H2O2400 μmol/L处理),维生素E组(Vit-E,1 μg/mL),PNS 1、0.1 mg/mL组(同模型组,并分别用PNS 1、0.1 mg/mL处理),同时设无细胞的空白对照组,每组设3个复孔。细胞于接种18 h后,向培养液中加入相应药物处理24 h。之后按照文献方法诱导细胞的氧化应激[6],除正常组外,均给予终浓度为400 μmol/mL的H2O2损伤1 h造成氧化应激模型,收集细胞供各项指标的检测。
2.2 ELISA检测APP,α-分泌酶、β-分泌酶及γ-分泌酶水平 取对数生长期细胞,调整细胞数至1×105个/mL接种于6孔板。分组及药物处理同“2.2”项。完成给药及H2O2处理后,采用细胞刮彻底收集细胞,超声破碎细胞,4 ℃下3 000 r/min离心10 min,收集上清液按照试剂盒方法测定细胞匀浆中APP,α-分泌酶、β-分泌酶及γ-分泌酶水平。
2.3 RT-PCR检测Aβ生成相关基因转录水平 取对数生长期细胞,调整细胞数至1×105个/mL接种于培养瓶。分组及药物处理同“2.2”项。细胞培养结束后,以TRIzol裂解细胞,提取总RNA。1%琼脂糖凝胶中,于150 V电泳30 min,同时将RNA适度稀释后测定OD260及OD280。观察18 s和25 s条带清晰,亮度比约为1∶2,无明显降解,且OD260/OD280在1.8~2.0,即认为提取的RNA 质量合格。根据测定的OD260将RNA稀释至0.2 μg/μL,按照TIANScript cDNA 第一链合成试剂盒方法将RNA转录为cDNA。取6 μL稀释后的cDNA,加入Reaction Buffer 2.5 μL,dNTP 4 μL, 上、下游引物各0.8 μL,Taq酶0.3 μL,加ddH2O补充体积至25 μL,PCR引物分别为:GAPDH上游引物F:GGCACAGTCAAGGCTGAGAATG,下游引物R:ATGGTGGTGAAGACGCCAGTA;PS-1 F:ACAGAGACTTGACAACCCTG,下游引物R:AGGCATGGATGACCTTGTAG;β-分泌酶(BACE)上游引物F:TGCCATCACTGAATCGGACAA,下游引物R:AAGATGTTCGGAATGTGGGTCTG;金属蛋白酶结构蛋白-10(ADAM10)上游引物F:ACTACTGCTTGGCCTATGTC,下游引物R:GTAGCCACAGTCACACTCTT。每个样本设3个复孔。PCR仪扩增程序为:95 ℃,2 min预变性,94 ℃,35 s变性,56 ℃左右,40 s退火,72 ℃,45 s延伸,扩增25~40个循环后72 ℃充分延伸10 min。取PCR产物于琼脂糖凝胶上样,80 V下,电泳30 min,Gel Logic 212 PRO凝胶成像系统拍照,Quantity One 4.40软件对结果图片进行灰度分析,以各目的基因与内参基因(Gapdh)的灰度值之比作为目的基因的相对表达量。
2.4 Western Blotting检测Aβ生成相关蛋白表达 取对数生长期细胞,调整细胞数至1×105个/mL接种于培养瓶。分组及药物处理同“2.2”项。细胞培养结束后,4 ℃裂解细胞30 min,收集裂解物细胞4 ℃ 12 000 r/min离心5 min,稀释蛋白质量浓度至3 mg/mL,PCR仪100 ℃变性6 min。15%和8% SDS-PAGE 110 V分离样品。于低温100 V 1 h将蛋白转至PVDF膜上,室温封闭2 h。分别以适量稀释的APP(1∶100)、PS-1(1∶100)、ADAM10(1∶100)、Gapdh(1∶200)一抗,封闭过夜。TBST 充分洗涤后,将膜与溶于封闭液中的二抗羊抗兔IgG-HRP(1∶3 000)室温孵育2 h,加入化学发光试剂,以Image station system 4000MM对经化学发光处理的PVDF膜进行实时数码曝光,保存数码曝光结果。采用Quantity One 4.40软件对结果图片进行定量分析,以各目的蛋白与内参蛋白(Gapdh)定量灰度值之比作为目的蛋白的表达量。
3 结果
3.1 PNS对氧化应激的PC12细胞中APP蛋白,α-分泌酶、β-分泌酶及γ-分泌酶水平的影响 实验结果见表1。模型组 PC12 细胞的APP蛋白水平有增加的趋势,但与正常组没有显著差异(P>0.05),1 μg/mL的PNS能显著降低APP蛋白水平。模型组细胞α-分泌酶水平较正常组显著降低(P<0.05),而使用0.1 μg/mL PNS显著提高α-分泌酶水平,1 μg/mL PNS仅有提高α-分泌酶水平的趋势。PC12细胞经H2O2刺激后,其β-分泌酶水平没有显著变化,PNS对β-分泌酶水平也没有显著影响。PC12细胞中γ-分泌酶水平在H2O2刺激后显著升高(P<0.001),0.1、1 μg/mL PNS均能显著降低γ-分泌酶水平(P<0.01)。
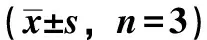
表1 PNS对氧化应激的PC12细胞中APP、α-分泌酶、β-分泌酶及γ-分泌酶水平的影响
3.2 PNS对氧化应激的PC12细胞中ADAM10、BACE及PS-1基因转录的影响 实验结果见表2及图1。经H2O2处理后,模型组细胞ADAM10 mRNA表达显著降低(P<0.01),PNS 1、0.1μg/mL均能显著增加ADAM10 mRNA的表达(P<0.01,P<0.001)。模型组BACE mRNA表达量显著高于正常组(P<0.05),PNS只有较弱的减少BACE mRNA表达的趋势。模型组PS-1 mRNA表达量显著高于正常组(P<0.05),PNS 1、0.1 μg/mL均能显著减少PS-1 mRNA的表达(P<0.05,P<0.01)。

表2 PNS对氧化应激的PC12细胞中ADAM10、BACE及PS-1 mRNA转录的影响
3.3 PNS对氧化应激的PC12细胞中APP,ADAM10及PS-1蛋白表达的影响 实验结果见表3及图2。经H2O2处理的PC12细胞中,APP蛋白表达显著高于正常组(P<0.05),0.1 μg/mL的PNS能显著抑制H2O2诱导的PC12细胞中APP蛋白的生成(P<0.05)。模型组PC12细胞接受H2O2处理后,ADAM10蛋白表达显著减少(P<0.05),1、0.1 μg/mL的PNS均能显著增加ADAM10蛋白表达(P<0.05)。模型组细胞接受PS-1蛋白表达显著增加(P<0.05),1、0.1 μg/mL的PNS均能显著减少PS-1蛋白表达(P<0.05,P<0.01)。

图1 PNS对氧化应激的PC12细胞中Gapdh,ADAM10,BACE及PS-1 mRNA表达的影响

表3 PNS对氧化应激的PC12细胞中APP,ADAM10及PS-1 蛋白表达的影响

图2 PNS对氧化应激的PC12细胞中GAPDH,APP,ADAM10及PS-1蛋白表达的影响
4 讨论
氧化应激能通过激活细胞内腺苷酸环化酶,催化环磷腺苷(cyclic adenosine monophosphate,cAMP)的活化,从而促进APP蛋白的大量生成[7],APP经β-分泌酶及γ-分泌酶的加工后形成的Aβ蛋白,具有很强的寡聚特性。氧化应激还可以通过炎症细胞因子途径,增加Aβ蛋白的生成。如Gu等[8]在过氧化氢刺激的SH-SY5Y细胞中发现,过氧化氢的刺激能使APP及BACE上游启动子去甲基化,促进使DNA与NFr-κB的结合,此外还通过影响组蛋白去乙酰化酶,进一步在基因表达的调控水平增加了Aβ蛋白的生成。此外,氧化应激还可通过激活双链RNA依赖性蛋白激酶,促进真核生物翻译起始因子磷酸化,从而增加BACE1基因的转录后翻译水平,促进Aβ蛋白的积累[9]。而过度积累的Aβ蛋白反过来又进一步引发了新的氧化损伤[10],如APP/PS-1转基因痴呆小鼠模型中就发现了非常显著的免疫系统氧化应激现象[11]。抑制Aβ蛋白加工成熟的治疗手段可以减轻Aβ带来的氧化损伤,如PS-1基因为γ-分泌酶复合物组分之一,Fiorini等的研究发现,快速老化痴呆模型小鼠大脑经PS-1反义沉默处理后,其学习记忆能力有所提高,大脑氧化应激程度显著减轻[12]。Webster等发现[13], PC12细胞在经慢性缺氧诱发的氧化应激条件下,ADAM10表达量显著减少,并且Aβ蛋白生成增加。可见,氧化应激是引起细胞内Aβ蛋白代谢异常的主要因素之一。目前已发现很多中药来源的有效成分,可能通过缓解机体的氧化应激,发挥对痴呆的治疗作用[14]。
在本课题组前期研究中运用外源性的H2O2刺激PC12细胞,导致细胞进入氧化应激状态,PNS处理能减少细胞内ROS水平,增强细胞内的主要抗氧化酶,如SOD、GSH-Px的活力,缓解细胞的氧化应激[5]。本研究在氧化应激的PC12细胞中,观察到了APP蛋白表达的增加,α-分泌酶表达减少以及γ-分泌酶的表达增加,可见,H2O2的刺激改变了细胞中APP蛋白的代谢水平,引发Aβ蛋白的积累从而导致Aβ蛋白介导的细胞退行性病变。更有意思的是,PNS的处理减少了氧化应激引起的PC12细胞中APP蛋白积累,增加了α-分泌酶表达,并减少γ-分泌酶的表达。可见,缓解氧化应激造成的Aβ蛋白代谢异常也是PNS治疗阿尔茨海默氏病的可能途径之一。
参考文献:
[1] Bertram L, Tanzi R. Thirty years of Alzheimer′s disease genetics: the implications of systematic meta-analyses[J].NatRevNeurosci, 2008, 9(10): 768-778.
[2] Wang C H, Wu S B, Wu Y T,etal. Oxidative stress response elicited by mitochondrial dysfunction: Implication in the pathophysiology of aging[J].ExpBiolMed, 2013, 238(5):450-460.
[3] 孙启祥, 陆 阳, 胡雅儿, 等. 三七有效成分对SD大鼠拟痴呆模型的影响[J]. 中国临床康复, 2004, 8(28): 6152-6154.
[4] 郭长杰, 伍杰雄, 李若馨. 三七总皂苷对痴呆模型大鼠大脑皮质内神经递质含量的影响[J]. 中国临床药学杂志, 2004, 13(3): 150-152.
[5] 夏 星,钟振国,冯丹霞. 三七总皂苷保护PC12细胞对抗过氧化氢损伤的作用[J]. 中国实验方剂学杂志, 2013, 19(4):216-219.
[6] Crispo J A, Piché M, Ansell D R,etal. Protective effects of methyl gallate on H2O2-induced apoptosis in PC12 cells[J].BiochemBiophysResCommun, 2010, 393(4):773-778.
[7] Canepa E, Domenicotti C, Marengo B,etal. Cyclic adenosine monophosphate as an endogenous modulator of the amyloid-β precursor protein metabolism[J].IUBMBLife, 2013, 65(2): 127-133.
[8] Gu X, Sun J, Li S,etal. Oxidative stress induces DNA demethylation and histone acetylation in SH-SY5Y cells: potential epigenetic mechanisms in gene transcription in Aβ production[J].NeurobiolAging, 2013, 34(4): 1069-1079.
[9] Mouton-Liger F, Paquet C, Dumurgier J,etal. Oxidative stress increases BACE1 protein levels through activation of the PKR-eIF2α pathway[J].BiochimBiophysActa, 2012, 1822(6): 885-896.
[10] Moreira P I, Santos M S, Oliveira C R,etal. Alzheimer disease and the role of free radicals in the pathogenesis of the disease[J].CNSNeurolDisordDrugTargets, 2008, 7(1): 3-10.
[11] Robinson R A, Cao Z, Williams C. Oxidative stress in CD90+ T-cells of AβPP/PS-1 transgenic mice[J].JAlzheimersDis, 2013, 37(4): 661-666.
[12] Fiorini A, Sultana R, Förster S,etal. Antisense directed against PS-1 gene decreases brain oxidative markers in aged senescence accelerated mice (SAMP8) and reverses learning and memory impairment: A proteomics study[J].FreeRadicBiolMed, 2013, 65:1-14.
[13] Webster N J, Green K N, Peers C,etal. Altered processing of amyloid precursor protein in the human neuroblastoma SH-SY5Y by chronic hypoxia[J].JNeurochem, 2002, 83(6): 1262-1271.
[14] 段小花, 李秀芳, 周宁娜, 等. 天麻提取物对血管性痴呆大鼠学习记忆及海马氧化损伤的作用[J]. 中成药, 2011, 53(7): 1138-1141.
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
航天电子对抗(2022年4期)2022-10-24
世界科学技术-中医药现代化(2020年2期)2020-07-25
生物学教学(2019年3期)2019-03-22
中国科技纵横(2018年2期)2018-11-29
中成药(2018年7期)2018-08-04
中成药(2018年6期)2018-07-11
中成药(2018年5期)2018-06-06
癌变·畸变·突变(2015年3期)2015-02-27