
电刺激微生物调控人工湿地中CH4和N2O气体排放研究*
2023-11-24 09:18肖东彩薛鸿普梁杨杨
环境污染与防治 2023年11期
肖东彩 薛鸿普 梁杨杨
(1.银川能源学院化学与生物工程学院,宁夏 银川 750150;2.重庆大学环境与生态学院,重庆 400044;3.宁夏宝丰能源集团股份有限公司,宁夏 银川 753200)
针对温室气体(GHGs,包括CO2、CH4及N2O)排放导致的全球变暖问题,我国提出“2030年前达到碳排放峰值、2060年前实现碳中和”的目标愿景[1-2]。现阶段碳减排技术仍处于研发阶段,想要大规模应用仍存在许多制约。因此,亟需寻找一个节能减排、协同增效的GHGs调控技术。
人工湿地(CW)耦合微生物燃料电池(MFC)是一种新型的低碳型污水处理技术,YADAV等[3]首次将MFC嵌入到CW,构建的CW-MFC系统既可以强化CW处理污水的效率,也能回收产生的电能。LIU等[4]通过生物电化学方法来控制CH4的排放,发现电力生成和CH4排放是同时发生的,这进一步说明电化学细菌和产甲烷菌是和谐共存的。
本实验通过搭建新型升流式人工湿地(UCW)型MFC(UCW-MFC),选择菖蒲为湿地植物并将其嵌入到MFC的阳极构建根际阳极UCW-MFC系统,研究微生物产电活动对湿地CH4排放通量和N2O排放通量的影响规律及作用机制,为GHGs调控提供新的思路。
1 实验与方法
1.1 UCW-MFC的搭建与启动
1.1.1 植物的选择
植物(如美人蕉、灯心草、菖蒲、茭白、水葱、芦苇、水鬼蕉、再力花等)是CW系统的重要影响因素,本研究选取缺氧忍耐力强的根茎型湿地植物菖蒲种植于阳极层来构建系统。对购置的菖蒲根系进行清洗,使用人工配水驯化约一周并挑选生命力旺盛的植物进行实验。
1.1.2 污泥的采集及驯化
生物阳极接种某污水处理厂厌氧段污泥,驯化期间,阴/阳极两端接入自动电压收集装置并间歇启动。
1.1.3 电极材料的选择
考虑到生物的附着性能和快速收集电子的需求,选择中间夹有不锈钢丝网(SSM,丝径2 mm,孔径0.5 mm)的碳纤维毡(CAF)作为阳极材料。
1.1.4 装置的搭建与启动
反应主体装置采用高55 cm、直径15 cm的有机玻璃制成,采用高28 cm、直径14 cm的静态箱收集GHGs。如图1所示,装置底部留有锥形进水口,底部承托层高度为5 cm,中间层高度为10 cm,均填充砾石。生物阳极和空气阴极层厚度约为3~4 cm,阴/阳极分别用钛丝引出并连接一个1 000 Ω的外电阻形成通路。
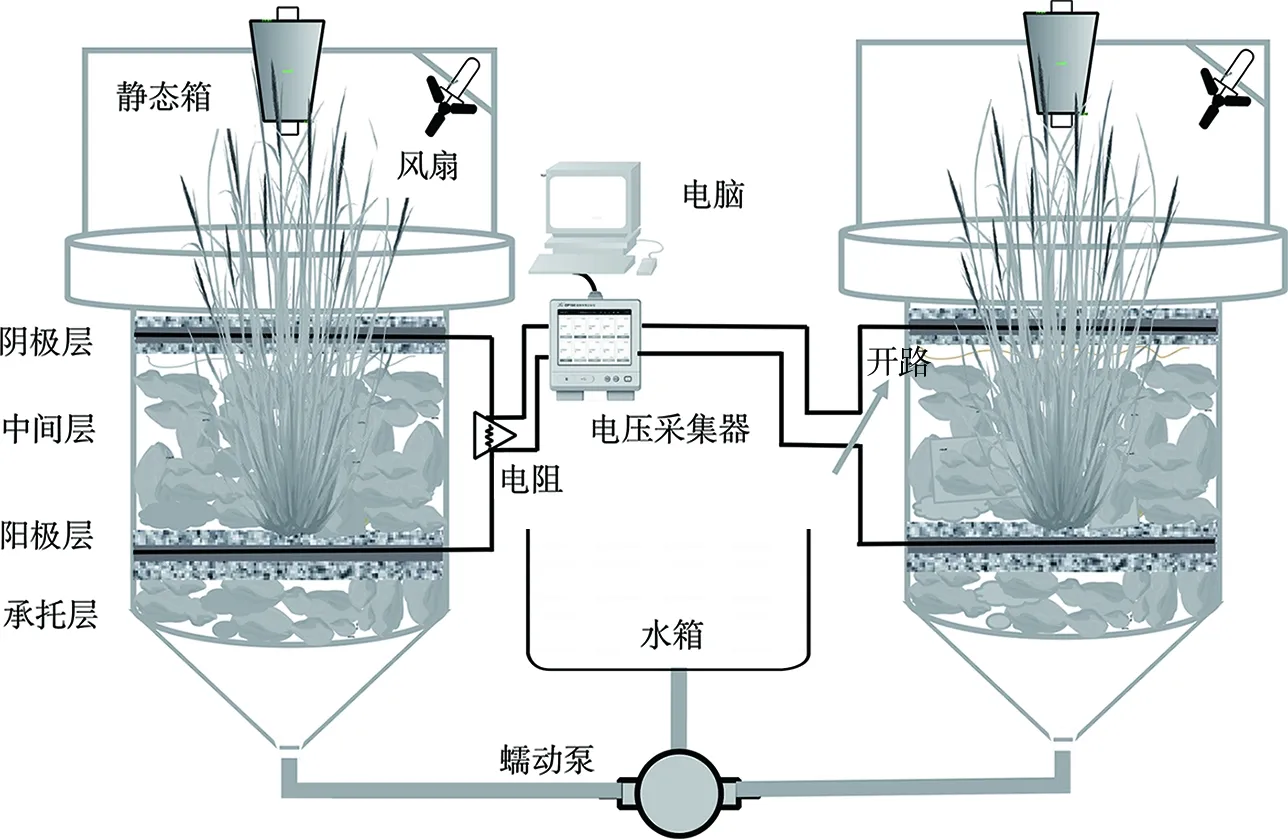
图1 UCW-MFC装置示意图Fig.1 Schematic diagram of UCW-MFC
1.2 实验设计
菖蒲对GHGs排放的影响实验中,水箱中人工配水成分:磷酸缓冲溶液5 mmol/L,葡萄糖 0.2 g/L,NH4Cl 0.20 g/L,KCl 0.13 g/L,NaHCO33.13 g/L。种植5株菖蒲(平均高度为48.92 cm)的系统作为实验组(启动成功时的电压约为0.66 V),未种植菖蒲的系统作为对照组(启动成功时的电压约为0.38 V),实验组同时考虑了开路、闭路对系统影响。
基质共代谢对GHGs排放的影响实验中,各系统均种植5株菖蒲(平均高度为48.92 cm),均为闭路,水箱中人工配水成分:磷酸缓冲溶液5 mmol/L,NH4Cl 0.20 g/L,KCl 0.13 g/L,NaHCO33.13 g/L;葡萄糖质量浓度分别设为0、300、500 mg/L。
1.3 实验测试方法
1.3.1 GHGs的收集与检测
每日8:00—10:00采用静态箱收集气体,箱体内部设有采气孔和风扇,以便收集和混合气体。采用配有氢火焰离子化检测器(HFID)和热导检测器(TCD)的气相色谱仪(Agilent 6890B)来检测CH4和N2O,其载气为氮气,色谱柱为Agilent HP-PLOT-Q柱(30 m×530 μm×40 μm)。
1.3.2 电压及功率密度曲线的测定
采用万用表(FLUKE 17B+)检测闭合电压和开路电压。极化曲线及功率密度曲线使用变阻箱分别检测9 000、8 000、7 000、6 000、5 000、4 000、3 000、2 000、1 000、800、500、100、50 Ω时的电压,并计算功率密度。
1.3.3 微生物高通量测序
实验选用V3~V4片段的细菌来扩增16S rRNA基因,其引物分别为338F(5’-ACTCCTACGGGAGGCAGCA-3’)和806R(5’-GGACTACHVGGGTWTCTAAT-3’)。实验具体流程为阳极污泥宏基因组提取、功能区域细菌扩增、产物定量和混样、文库构建和文库质检与测序。样品由上海某生物公司进行测序。
2 结果与分析
2.1 菖蒲对GHGs排放的影响
如图2(a)所示,有菖蒲和无菖蒲的系统在闭路条件下CH4排放通量分别为(0.35±0.01)、(0.17±0.01) mg/(m2·h),N2O排放通量分别为(122.85±3.68)、(91.18±2.78) μg/(m2·h),显然菖蒲加入提高了GHGs的排放。这可能是因为菖蒲自身存在通气组织,根系周围的气体被茎部维管传输至大气中;其次微生物易附着在菖蒲根系且在厌氧条件下利用极少量的腐殖质(纤维素)作为自身的营养物质,导致呼吸速率加快,产出更多的CH4和N2O。如图2(b)可知,菖蒲对系统产电影响较大,有菖蒲和无菖蒲的系统最大功率密度分别为(1.62±0.05)、(0.76±0.06) W/m3,系统内阻分别为154.27、237.55 Ω,可见电化学细菌可利用菖蒲增加系统产电并降低其内阻以加速氧化还原反应的进行。
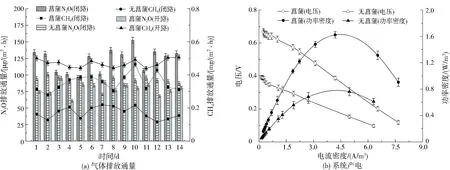
图2 有无菖蒲对系统气体排放通量和产电的影响Fig.2 Effect of calamus or non-calamus on gas emission flux and electricity generation in the system
UCW-MFC在开路下相当于CW,如图2(a)所示,有菖蒲的系统CH4和N2O排放通量在开路条件下分别约为(0.47±0.02) mg/(m2·h)、(79.24±2.38) μg/(m2·h),相比闭路条件下,CH4排放通量增加约0.12 mg/(m2·h)。而阳极产生的电子在开路条件下不能传输至阴极,遏制还原产生N2O,N2O排放通量降低约43.61 μg/(m2·h)。系统开路下,阳极产甲烷菌的菌群丰富度多于产电菌,导致细菌呼吸作用产生更多CH4,这进一步验证产电微生物和产甲烷微生物之间存在竞争且可以共存,而N2O被抑制是中断电子传递导致的。
2.2 基质共代谢对GHGs排放的影响
共基质(根系腐殖质和外加葡萄糖)是影响系统产电和GHGs排放的重要因素。如图3(a)所示,根系+0 mg/L葡萄糖、根系+300 mg/L葡萄糖和根系+500 mg/L葡萄糖的CH4排放通量分别为 (0.12±0.01)、(0.73±0.02)、(1.38±0.04) mg/(m2·h),N2O排放通量分别为0、(63.85±1.98)、(12.41±0.38) μg/(m2·h),可以看出,系统在高浓度基质共代谢下易产生CH4,主要原因是充足的COD刺激产酸菌产生乙酸等小分子有机物,进一步被产甲烷菌转化为CH4[5]。相反,N2O排放通量随着共基质浓度增大而降低,可能因为共基质浓度增加使得电化学细菌活性增加,刺激自身胞外电子的传递以加速反硝化反应的进行,进而减少N2O的积累和排放。本研究GHGs排放趋势与文献[6]的研究结果相似。系统中电化学细菌和产甲烷菌存在竞争状态,产甲烷菌和电化学细菌均适合在较高COD的情况下生存和产电且CH4排放通量较高。
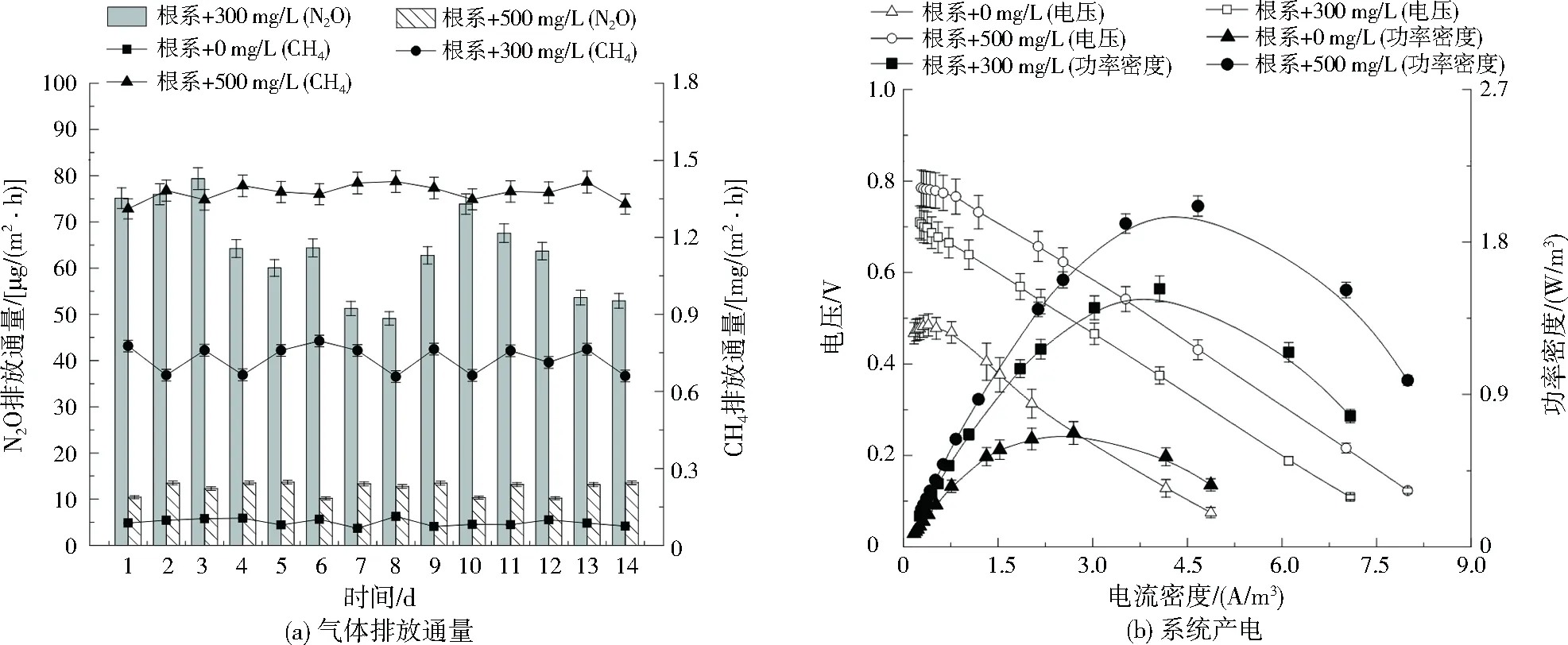
注:根系+300 mg/L是指种植菖蒲并外加300 mg/L葡萄糖,其余类推。根系+0 mg/L条件下N2O排放通量未检测出。图3 共基质对系统气体排放通量和产电的影响Fig.3 Effects of comatrix on gas emission flux and electricity generation in the system
电刺激作用下,大量的产甲烷菌繁殖使得呼吸产生的CH4排放通量增加。如图3(b)所示,根系+300 mg/L葡萄糖和根系+500 mg/L葡萄糖的最大功率密度分别为(1.52±0.04)、(2.01±0.06) W/m3,内阻分别为279.33、285.52 Ω,由此可知共基质浓度与产电、CH4排放通量正相关,并与N2O排放通量负相关。根系+0 mg/L葡萄糖时,系统最大功率密度为(0.67±0.03) W/m3,内阻为292.18 Ω,可能的原因是生物阳极的电化学细菌(如脱硫叶菌属(Desulfobulbus)、假单胞菌属(Pseudomonas)等)利用菖蒲根系腐烂物(有机酸、聚合碳水化合物以及死细胞物质等)来供自身消耗并产电[7]。而表观内阻在根系+0 mg/L葡萄糖下比根系+300 mg/L葡萄糖和根系+500 mg/L葡萄糖都要大,可能原因是微生物不能充分得到营养物质导致传递质子和产生电子的能力变弱。NI等[8]使用MFC处理猪场废水,结果发现适宜COD浓度可有效增强系统产电,而高负荷COD抑制猪场废水中污染物的降解,进一步研究发现充足的底物在生物阳极未降解完全,并游离到阴极消耗溶解氧使其形成一层好氧生物膜阻挡氧气渗入到阴极。
2.3 微生物高通量测序结果分析
为进一步分析有无菖蒲及开闭路的系统微生物群落,对生物阳极共基质(根系+500 mg/L葡萄糖)进行属水平微生物丰富度分析。如图4所示,属水平的物种丰富度表明,有菖蒲的系统在开路条件下,微生物包括不动杆菌属(约78.77%)、变形菌属(约0.31%)、厚壁菌属(约0.56%)、假单胞菌属(约0.42%)、硝化螺旋菌属(约0.40%)、拟杆菌属(约0.89%)、地杆菌属(约0.09%)、甲烷丝状菌属(约3.29%)、类芽胞八叠球菌属(约3.51%)及毛球菌属(约0.25%)等。有菖蒲的系统在闭路条件下,微生物包括不动杆菌属(约1.85%)、变形菌属(约1.43%)、厚壁菌属(约2.77%)、假单胞菌属(约18.77%)、硝化螺旋菌属(约3.41%)、拟杆菌属(约7.68%)、地杆菌属(约0.91%)、甲烷丝状菌属(约7.28%)、类芽胞八叠球菌属(约0.10%)及毛球菌属(约2.62%)等。而无菖蒲的系统在闭路条件下,微生物包括不动杆菌属(约1.26%)、变形菌属(约1.31%)、厚壁菌属(约1.92%)、假单胞菌属(约6.16%)、硝化螺旋菌属(约2.34%)、拟杆菌属(约7.79%)、地杆菌属(约1.73%)、甲烷丝状菌属(约6.94%)、类芽胞八叠球菌属(约3.91%)及毛球菌属(约0.45%)等。其中电化学细菌有变形菌属[9]、厚壁菌属[10]、不动杆菌属[11]、拟杆菌属[12]及地杆菌属[13]等,产甲烷菌有甲烷丝状菌属[14],反硝化细菌有硝化螺旋菌属[15]。实验条件改变会使微生物群落发生变化,最终使系统产电电压及GHGs排放通量发生改变。
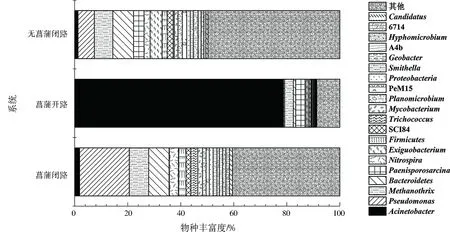
注:Acinetobacter为不动杆菌属,Methanothrix为甲烷丝状菌属,Bacteroidetes为拟杆菌属,Paenisporosarcina为类芽胞八叠球菌属,Nitrospira为硝化螺旋菌属,Exiguobacterium为微小杆菌属,Firmicutes为厚壁菌属,Trichococcus为毛球菌属,Mycobacterium为分枝杆菌属,Planomicrobium为游动球菌属,Proteobacteria为变形菌属,Smithella为史密斯氏菌属,Geobacter为地杆菌属,Hyphomicrobium为生丝微菌属,Candidatus为假丝酵母菌属。图4 属水平微生物丰富度Fig.4 Microbial abundance at genus level
不同系统共基质(根系+500 mg/L葡萄糖)条件下的物种多样性指标见表1,闭路条件下的有菖蒲系统,其Chao1、Shannon及Simpson指数都优于开路条件下的有菖蒲系统和闭路条件下的无菖蒲系统,说明系统闭合及菖蒲存在更有助于电化学细菌产生电子,也丰富了阳极的物种丰富度。

表1 多样性指数Table 1 Diversity index
3 GHGs排放的机制分析
CH4排放分为发酵阶段、产酸阶段和产气阶段,在厌氧发酵中,产酸菌将大分子有机物降解为小分子有机物,并通过产甲烷菌和(反)硝化细菌转化为GHGs,当然系统的阴极也会产生CH4,这是因为阳极产生质子(H+)传输至阴极发生还原反应形成H2,阴极在好氧条件下产生CH4[16]。文献[17]报道,沼泽湿地或者稻田地采用同位素示踪法追踪CH4足迹,结果发现共基质阳极发酵产生小分子有机物(CH3COOH、HCOOH、CH3OH)生成的CH4约占70%,而空气阴极还原(CO2)产生的CH4仅占30%。
N2O的产生较为复杂,分为硝化阶段和反硝化阶段。硝化反应是在好氧状态下进行的,首先在好氧状态下发生氨氧化反应继而发生硝化反应。MILOSLAV等[18]通过实验发现反硝化作用在厌氧条件下发生,而N2O主要是反硝化细菌在厌氧下产生的。ULRIKE等[19]通过研究发现,反硝化细菌产生的N2O不是最终产物,这是因为湿地表面的浮水层阻挡氧气进入内部进一步会加速反硝化反应的进行并导致反硝化的最终产物是N2而不是N2O。
如图5所示,电信号刺激下微生物丰富度增加,导致对有机质的生物降解加快,并直接刺激电化学细菌在阳极的积累,从而加速阳极厌氧降解的过程,提高了系统氧化还原的能力。为深入探讨根系+500 mg/L葡萄糖条件下C、N元素对菖蒲生长的影响,测量菖蒲的高度及C含量。如表2所示,结果表明少量C、N养分被菖蒲吸收以供自身生长,实验后菖蒲高度增加13.42 cm,菖蒲的通气组织会储存微量的营养元素。

表2 系统运行前后菖蒲的变化特征Table 2 Change characteristics of calamus before and after operation

注:共基质是指根系+500 mg/L葡萄糖。图5 共基质生物阳极表面微生物扫描电镜图Fig.5 Bio-anode scanning electron microscope images
4 结 语
(1) 有菖蒲的系统在闭路条件下CH4排放通量为(0.35±0.01) mg/(m2·h),最大功率密度为(1.62±0.05) W/m3;开路的系统不适合调节GHGs,CH4排放通量增加0.12 mg/(m2·h),N2O排放通量降低43.61 μg/(m2·h)。共基质浓度增加会加大CH4排放通量,降低N2O排放通量。
(2) 甲烷丝状菌属和硝化螺旋菌属是产生CH4和N2O的主要微生物且在系统中和电化学细菌之间存在竞争关系。
(3) MFC嵌入到CW为控制GHGs的排放提供了绿色思路。
猜你喜欢
河南医学研究(2022年19期)2022-10-19
选煤技术(2022年3期)2022-08-20
大电机技术(2021年5期)2021-11-04
基层中医药(2020年6期)2020-09-11
环球时报(2020-06-30)2020-06-30
生态学报(2019年11期)2019-07-08
中国交通信息化(2019年3期)2019-06-18
中国钼业(2018年4期)2018-08-24
旅游(2018年6期)2018-06-08
中华手工(2016年9期)2016-11-10
