miR-146b-5p靶向ZNRF2影响IL-17a诱导乳腺癌细胞增殖、迁移和侵袭
2021-08-21 05:19:56龚莉莉叶春梅武汉市妇女儿童医疗保健中心乳腺外科武汉430014
中国免疫学杂志 2021年14期
王 娜 龚莉莉 叶春梅(武汉市妇女儿童医疗保健中心乳腺外科,武汉430014)
乳腺癌是世界范围内女性最常见的恶性肿瘤,其异质性和转移性导致患者死亡率居高不下[1]。研究乳腺癌发生和转移的机制,对其精准治疗具有重要意义。
研究表明,IL-17a(interleukin-17a)在部分乳腺浸润性导管癌组织中高表达,体外可促进乳腺癌MCF-7细胞的侵袭[2]。IL-17在乳腺癌血清样本中高表达,可促进乳腺癌进展[3]。miR-146在乳腺癌患者中表达下调并与乳腺癌的发生和恶化有关[4],miR-146b-5可通过靶向抑制IL-6表达抑制乳腺间质成纤维细胞的致癌作用[5]。ZNRF2与甲状腺乳头状癌的发生有关[6],参与miR-100抑制人骨肉瘤细胞增殖和耐药过程[7],且抑制ZNRF2表达可抑制非小细胞肺癌生长[8]。而ZNRF2在乳腺癌中的表达和作用尚不明确,其与miR-146b-5p和IL-17a在乳腺癌中是否存在关系还未可知。
本研究假设miR-146b-5p和ZNRF2参与IL-17a诱导乳腺癌进展的过程,以期为乳腺癌的靶向治疗提供新的研究靶点。
1 资料与方法
1.1 资料 选取本院病理科资料完整、诊断确切的女性乳腺癌手术切除癌症组织20例和正常癌旁组织20例,患者年龄23~68岁,平均(45.4±9.6)岁,术前未进行放疗、化疗及免疫治疗。人乳腺癌细胞株MDA-MB-231购自美国ATCC;DMEM高糖培养基和胎牛血清(fetal bovine serum,FBS)购自美国Gibco公司;胰蛋白酶、二甲基亚砜(dimethyl sulfoxide,DMSO)和四氮唑蓝(thiazolyl blue tetrazolium bromide,MTT)购自美国Sigma-Aldrich公司;IL-17a抗体、ZNRF2抗体、MMP-2抗体、MMP-9抗体、STAT3抗体、p-STAT3抗体和β-actin抗体购自英国Abcam公司;引物、miR-146b-5p模拟物(miR-146b-5p)、miR-146b-5p抑制剂(anti-miR-146b-5p)、ZNRF2抑制物(si-ZNRF2)、阴性对照(miR-con、anti-miR-con、si-con)及双荧光素酶ZNRF2突变和野生型载体购自上海吉玛制药有限公司;双荧光素报告基因检测试剂盒酶购自上海碧云天生物技术有限公司;BCA蛋白检测试剂盒购自江苏凯基生物技术股份有限公司;Lipofectamine 2000转染试剂、Total RNA提取试剂盒、real-time PCR试剂盒、反转录试剂盒(RTPCR)购自美国Invitrogen公司;real-time PCR仪购自美国Bio-Rad公司。
1.2 方法
1.2.1 免疫组化测乳腺癌组织和癌旁组织中的IL-17a将乳腺癌组织和癌旁组织于4%多聚甲醛固定,梯度乙醇脱水,然后石蜡包埋、切片,按常规程序脱蜡和水化,再用3%H2O2灭活内源性过氧化物酶15 min,枸橼酸缓冲液进行抗原修复,10%血清37℃封闭30 min,加入1∶200的IL-17a一抗4℃孵育过夜,以PBS代替一抗作阴性对照,用PBS缓冲液冲洗3次×10 min,加入稀释的二抗,37℃孵育30 min,洗3次,加DAB显色液,避光显色,终止显色。苏木素衬染胞核,梯度乙醇脱水,二甲苯透明,中性树胶封片,37℃干燥48 h,显微镜观察并拍照。
1.2.2 细胞培养和IL-17a处理 细胞培养:配制含10%FBS、100 U/ml青霉素和100 μg/ml链霉素的DMEM高糖培养液,将乳腺癌细胞MDA-MB-231培养于上述配制好的培养液,置于37℃、5%CO2、饱和湿度的培养箱中培养。待细胞成长至对数生长期,用胰蛋白酶消化传代。
IL-17a处理:用培养液稀释对数生长期的MDAMB-231细 胞,以1×104个/孔 接 种于96孔板培 养24 h,然后在培养液中加入终浓度为0、10、50、100及300 μg/L的IL-17a,继续培养24 h、48 h或72 h,进行后续实验。对照组常规培养,培养液不添加IL-17a。
1.2.3 细胞转染 将稀释的MDA-MB-231细胞以2×105个/孔接种于6孔板,培养至细胞融合度为80%~90%时进行转染。根据转染试剂盒说明书操作,将无血清OptiMEM培养液稀释脂质体和各组片段及载体(包括miR-con、miR-146b-5p、anti-miRcon、anti-miR-146b-5p、si-ZNRF2、si-con、双荧光素酶报告系统载体),将稀释的脂质体和各组载体片段等体积轻柔混匀,室温孵育20 min后加入培养好的MDA-MB-231细胞中,混匀后培养6 h后,换成DMEM高糖完全培养基,转染48 h,收集细胞,验证转染效果。
1.2.4 Real-time PCR检 测miR-146b-5p和ZNRF2 mRNA表达 收集IL-17a处理后的对数生长期的MDA-MB-231细胞,根据RNA提取试剂盒提取细胞总RNA,按照反转录PCR试剂盒操作要求进行cDNA合成,再以cDNA为模板按照real-time PCR说明书进行反应,合成miR-146b-5p和ZNRF2 mRNA。反应程序为:95℃2 min;94℃30 s、58℃40 s、72℃40 s,35个循环;72℃10 min。以2-ΔΔCt方法进行数据分析。
1.2.5 Western blot收集各组实验MDA-MB-231细胞,裂解细胞,收集蛋白并检测浓度。进行SDS-PAGE分离蛋白,转PVDF膜,室温封闭2 h,加入稀释的一抗,ZNRF2抗体(1∶2 000)、MMP-2抗体(1∶1 500)、抗MMP-9抗体(1∶1 000)、STAT3抗体(1∶5 000)、p-STAT3抗体(1∶2 000)和抗β-actin抗体(1∶3 000),4℃孵育过夜,PBST洗膜3次,然后加入稀释的二抗,室温孵育1 h,显影。以β-actin为内参照,分析蛋白表达水平。
1.2.6 MTT测定细胞活性 按照1.2.2的方法对MDA-MB-231细胞进行IL-17a处理,分别在培养至24 h、48 h、72 h时,每孔加入20 μl MTT溶液,培养4 h,弃上清,每孔再加入150 μl DMSO,室温振荡5 min,酶标仪测定OD490nm处吸光度(A)值。
1.2.7 Transwell检测细胞迁移和侵袭
1.2.7.1 迁移实验 将MDA-MB-231细胞培养至对数生长期,用无血清DMEM培养液稀释细胞至5×105个/ml。在Transwell上层加入100 μl稀释好的细胞,下层小室加入500 μl含10%FBS和IL-17a终浓度为100 μg/L的DMEM高糖培养基,培养72 h,弃去培养液,用棉签拭去上层小室未迁移的细胞,甲醛固定迁移的细胞30 min,结晶紫染色30 min,显微镜观察并计数迁移细胞。
1.2.7.2 侵袭实验 将Matrigel用4℃无血清培养基以1∶10比例稀释,加入上层小室,37℃烘干3 h,以下步骤同迁移实验,上层小室加入100 μl细胞,下层加入含10%FBS和IL-17a终浓度为100 μg/L的培养基,培养72 h,固定细胞,染色,计数。
1.2.8 双荧光素酶报告实验 将培养好的MDAMB-231细胞按照1.2.3步骤进行转染,将构建好的ZNRF2野生型(WT)和突变型(MUT)双荧光素酶报告载体分别与miR-con或miR-146b-5p共转染至培养好的细胞,转染48 h,收集细胞,验证转染效果,RIPA裂解液室温裂解转染后的细胞20 min,离心收集裂解上清,以海肾荧光素酶活性为内参照,计算萤火虫荧光素相对酶活性。
1.3 统计学处理 采用SPSS19.0统计软件进行数据分析,数据均以±s表示。组间比较采用独立样本t检验和单因素方差分析,以P<0.05为差异有统计学意义。
2 结果
2.1 IL-17a在乳腺癌患者组织中高表达 检测20例乳腺癌肿瘤组织和癌旁组织,结果表明,与癌旁组织相比,在大多乳腺癌肿瘤组织中IL-17a的表达呈阳性(P<0.05),见图1。说明IL-17在乳腺癌组织中高表达。
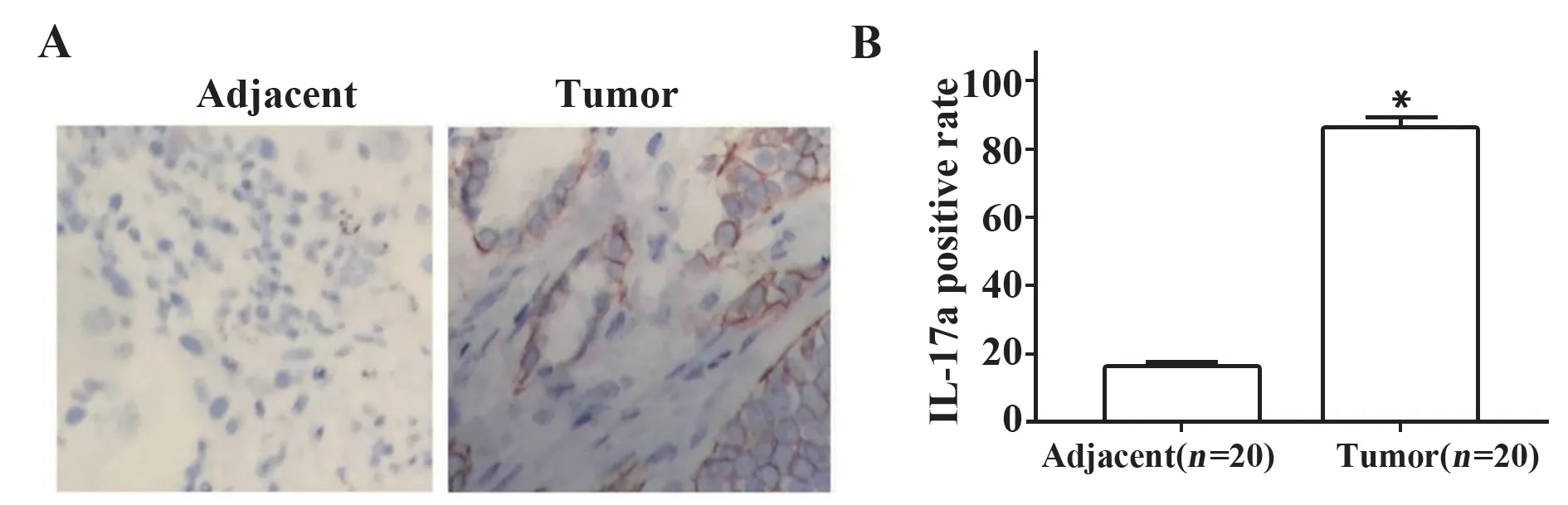
图1 乳腺癌组织及癌旁组织中IL-17的表达(免疫组化染色,×200)Fig.1 Expression of IL-17 in breast carcinoma tissue and adjacent tissues(IHC staining,×200)
2.2 IL-17a处理促进乳腺癌细胞株MDA-MB-231细胞的增殖、迁移和转移 分别用终浓度为0、10、50、100和300 μg/L的IL-17a处理乳腺癌MDA-MB-231细胞株。结果表明,与对照组(0 μg/L)相比,经IL-17a处理后MDA-MB-231细胞增殖活性在48 h和72 h均明显升高(P<0.05),迁移细胞数和侵袭细胞数均明显增多(P<0.05),且呈剂量依赖趋势。说明IL-17a可促进MDA-MB-231细胞增殖、迁移和侵袭。根据实验结果和经济原则,后续实验选择100 μg/L的IL-17a处理细胞72 h(图2)。
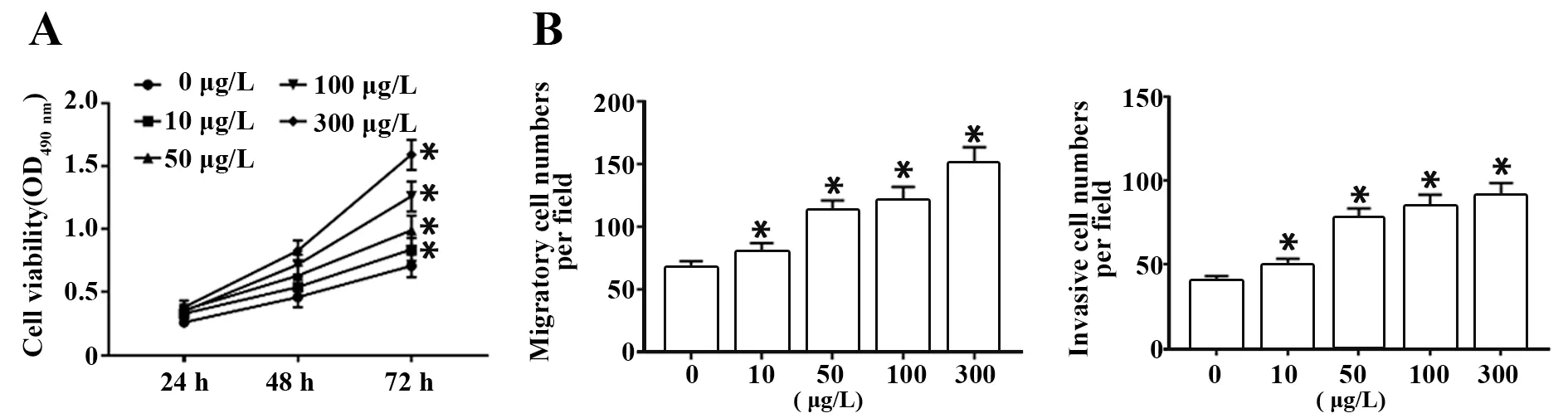
图2 IL-17a处理促进乳腺癌细胞株MDA-MB-231细胞的增殖、迁移和侵袭Fig.2 IL-17a treatment promoted proliferation,migration and invasion of breast cancer cell line MDAMB-231
2.3 IL-17a处理MDA-MB-231细胞抑制miR-146b-5p表达,促进ZNRF2表达 与对照组相比,IL-17a处理后MDA-MB-231细胞中miR-146b-5p的表达量明显降低(P<0.05),ZNRF2蛋白和mRNA表达量均明显升高(P<0.05),迁移侵袭蛋白MMP-2和MMP-9表达量明显升高(P<0.05)。说明IL-17a处理MDAMB-231细胞可抑制细胞中miR-146b-5p表达并促进ZNRF2的表达(图3)。
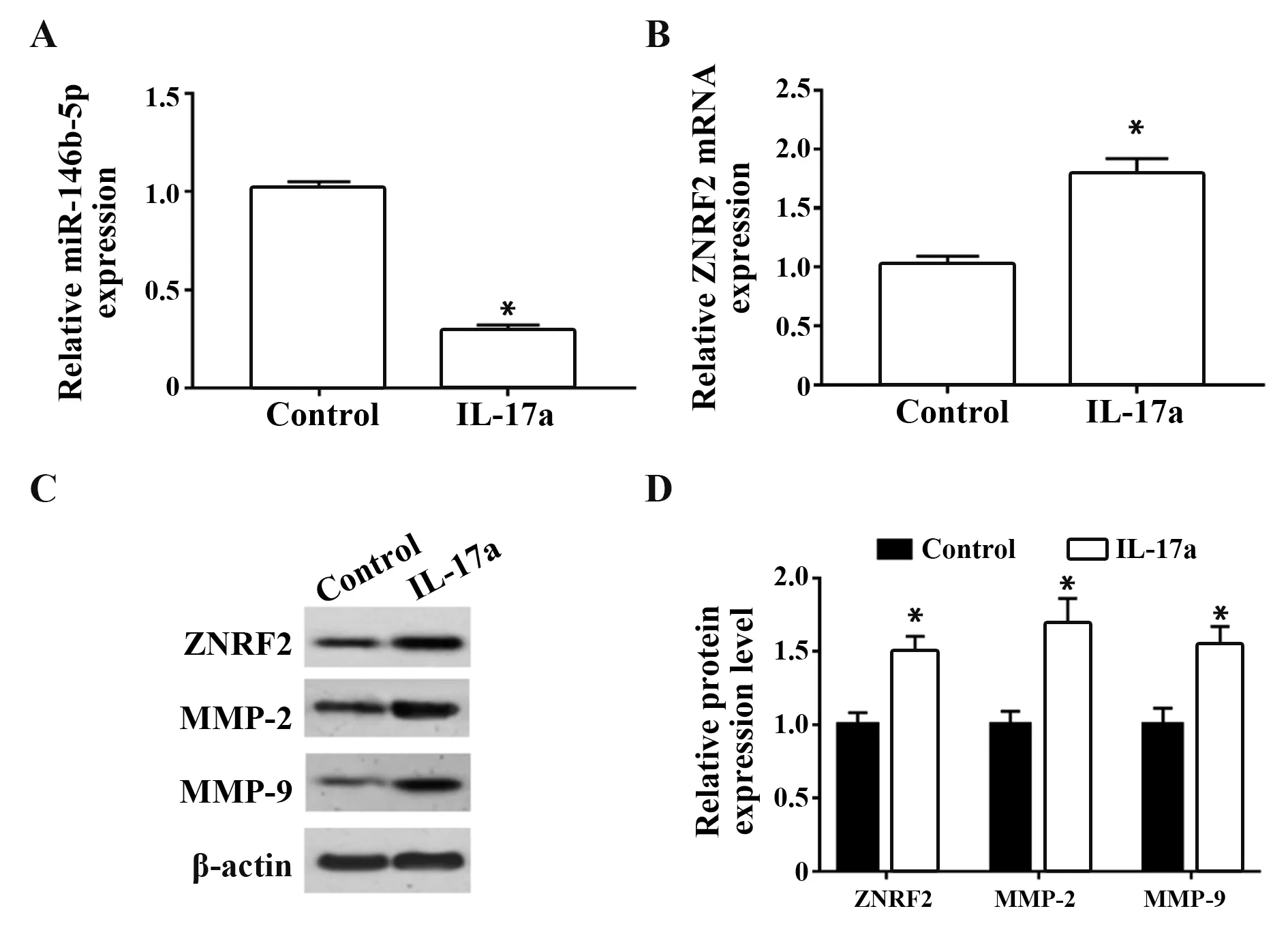
图3 IL-17a处理MDA-MB-231细胞对miR-146b-5p、ZNRF2、MMP-2和MMP-9含量的影响Fig.3 Effects of IL-17a treatment on levels of miR-146b-5p,ZNRF2,MMP-2 and MMP-9 in MDA-MB-231 cells
2.4 miR-146b-5p靶向调控ZNRF2 ZNRF2的3'-UTR序列中含有与miR-146b-5p互补的核苷酸序列,见图4A。双荧光素酶报告系统结果如图4B所示,过表达miR-146b-5p时,WT型ZNRF2萤火虫荧光素酶相对活性明显降低(P<0.05);而MUT型ZN-RF2的萤火虫荧光素酶相对活性无明显变化。Western blot结果表明,过表达miR-146b-5p时ZNRF2蛋白表达量明显降低(P<0.05);抑制miR-146b-5p表达时ZNRF2蛋白表达量明显升高(P<0.05),见图4C。
经检测,与癌旁组织相比,在20例乳腺癌组织中,miR-146b-5p含量明显降低(P<0.05),ZNRF2蛋白含量明显升高(P<0.05),IL-17a表达量与miR-146b-5p呈负相关,miR-146b-5p与ZNRF2含量呈负相关,见图4D~G。
2.5 IL-17a可逆转过表达miR-146b-5p对MDAMB-231细胞增殖、迁移和侵袭及STAT3信号通路的影响 结果显示,与Control组相比,IL-17a组MDAMB-231细胞增殖活性、迁移和侵袭数及细胞中ZNRF2、MMP-2、MMP-9蛋白表达升高,磷酸化STAT3(p-STAT3)蛋白表达升高;与Control组或miR-con组相比,miR-146b-5p组MDA-MB-231细胞增殖活性、迁移和侵袭细胞数及细胞中ZNRF2、MMP-2、MMP-9蛋白表达降低(P<0.05),p-STAT3蛋白表达降低(P<0.05);与miR-146b-5p组 相 比,IL-17a+miR-146b-5p组MDA-MB-231细胞增殖活性、迁移和侵袭细胞数及细胞中ZNRF2、MMP-2、MMP-9蛋白表达升高(P<0.05),p-STAT3蛋白表达升高(P<0.05),见图5。说明IL-17a可逆转过表达miR-146b-5p对MDA-MB-231细胞增殖、迁移、侵袭及STAT3信号通路的抑制作用。
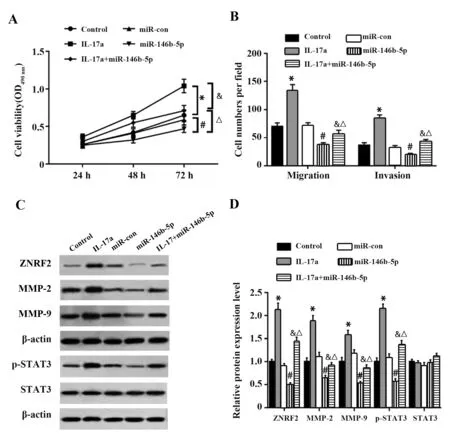
图5 IL-17a可逆转过表达miR-146b-5p对MDA-MB-231细胞增殖、迁移和侵袭的影响Fig.5 IL-17a reversed effects of miR-146b-5p overexpression on proliferation,migration and invasion of MDA-MB-231 cells
2.6 IL-17a可逆转沉默ZNRF2对MDA-MB-231细胞增殖、迁移和侵袭及STAT3信号通路的影响 结果显示,与si-con组相比,si-ZNRF2组MDA-MB-231细胞增殖活性、迁移和侵袭细胞及细胞中NRF2、MMP-2、MMP-9蛋白表达降低(P<0.05),p-STAT3蛋白表达量降低(P<0.05);与si-ZNRF2组相比,si-ZNRF2+IL-17a组MDA-MB-231细胞增殖活性、迁移和侵袭细胞及细胞中NRF2、MMP-2、MMP-9蛋白表达升高(P<0.05),p-STAT3蛋白表达增加(P<0.05),见图6。说明IL-17a可逆转沉默ZNRF2对MDAMB-231细胞增殖、迁移、侵袭及STAT3信号通路的抑制作用。

图6 IL-17a可逆转沉默ZNRF2对MDA-MB-231细胞增殖、迁移和侵袭及STAT3信号通路的影响Fig.6 IL-17a reversed effects of miR-146b-5p overexpression on STAT3 signaling pathway of MDA-MB-231 cells
3 讨论
IL-17由Th17细胞产生,可诱导细胞产生炎症效应物和抗微生物蛋白等因子,参与免疫性疾病和炎症的发生。近年研究表明,IL-17也在肿瘤中发挥促癌或抗癌效应[9-10]。IL-17a是IL-17家族成员之一,其单核苷酸多态性(SNPs)与中国女性患乳腺癌风险有关[11],而IL-17与乳腺癌的侵袭和转移有关[12-13]。本研究结果发现,IL-17a在乳腺癌组织中呈阳性表达,IL-17a可在体外抑制乳腺癌细胞MDAMB-231的增殖、迁移和侵袭,与相关报道结果一致[14],表明IL-17a可促进乳腺癌的发生发展。
miRNA是一类小分子非编码RNA,在真核生物体内广泛存在,参与调控细胞的增殖、凋亡和迁移等生物学过程,在癌症的发生发展中发挥重要作用[15]。研究显示,上调miR-146b-5p通过靶向抑制MMP-16表达降低肾癌细胞的增殖和侵袭能力[16]。姜黄素可通过上调miR-146b-5p表达抑制乳腺癌细胞转移,诱导细胞凋亡[17]。本研究结果表明,在乳腺癌组织中miR-146b-5p表达水平下降,过表达miR-146b-5p可抑制MDA-MB-231细胞增殖、迁移、侵袭,与相关报道结果一致,表明miR-146b-5p作为抑癌基因参与乳腺癌的发生发展过程,是乳腺癌治疗的潜在分子靶点[18]。本研究还显示,IL-17a可抑制MDA-MB-231细胞中miR-146b-5p表达,过表达miR-146b-5p可降低IL-17a对MDA-MB-231细胞增殖、迁移和侵袭的促进作用,与肖庆等[19]报道的IL-17a通过抑制miR-195-5p表达促进子宫内膜癌细胞增殖、迁移和侵袭的结果一致。
生物信息学预测结果发现,ZNRF2的3'-UTR序列中含有与miR-146b-5p互补的核苷酸序列,miR-146b-5p可能靶向调控ZNRF2表达。本研究通过双荧光素酶报告基因实验和Western blot证实了miR-146b-5p在乳腺癌MDA-MB-231细胞中靶向负调控ZNRF2表达,这也与IL-17a抑制MDA-MB-231细胞中miR-146b-5p表达,促进ZNRF2表达的结果一致。ZNRF2是环超家族E3泛素连接酶,可与细胞膜上的哺乳动物雷帕霉素靶点(mTOR)协同作用感知氨基酸供应和调节细胞生长,在多种癌症中发挥作用[20]。但目前,ZNRF2在乳腺癌中的表达和作用尚不明确。本研究发现,ZNRF2在乳腺癌组织中表达上调,IL-17a可促进MDA-MB-231细胞中ZNRF2的表达,沉默ZNRF2可抑制MDA-MB-231细胞增殖、迁移、侵袭,表明ZNRF2作为促癌基因参与乳腺癌的发生发展进程,靶向抑制其表达可延缓乳腺癌发展进程。
STAT3信号通路在多种恶性肿瘤中处于高度激活状态,参与肿瘤细胞的增殖和侵袭过程,如膀胱癌、肺癌[21-22]。本研究显示,IL-17a可激活乳腺癌细胞中STAT3信号通路,与相关报道结果一致[23]。本研究还显示,过表达miR-146b-5p或沉默ZNRF2可抑制IL-17a对乳腺癌细胞中STAT3信号通路的激活作用,进一步表明过表达miR-146b-5p可能通过负调控ZNRF2抑制STAT3信号通路的激活从而降低IL-17a对乳腺癌细胞恶性生物学行为的影响。
综上所述,IL-17a可促进乳腺癌细胞增殖、迁移和侵袭,过表达miR-146b-5p可通过靶向抑制ZNRF2表达降低IL-17a对乳腺癌细胞增殖、迁移和侵袭的促进作用。
猜你喜欢
保健医苑(2022年5期)2022-06-10 07:46:38
中学生物学(2021年8期)2021-11-02 04:53:14
天津医科大学学报(2021年4期)2021-08-21 02:14:50
中国临床医学影像杂志(2021年6期)2021-08-14 02:21:56
中日友好医院学报(2021年1期)2021-04-14 01:58:32
肝博士(2020年5期)2021-01-18 02:50:18
山东医药(2020年9期)2020-05-20 01:12:16
中国果业信息(2019年1期)2019-01-05 17:41:42
中国畜牧兽医文摘(2018年6期)2018-07-28 02:30:16
科学启蒙(2015年8期)2015-08-07 03:54:46