
铵离子对多杀菌素发酵生产的影响
2021-03-24 08:12扶教龙徐敏强汪佳琪胡翠英
苏州科技大学学报(自然科学版) 2021年1期
张 松, 扶教龙, 徐敏强, 汪佳琪, 张 瀚, 胡翠英, 白 净
(苏州科技大学 化学与生命科学学院,江苏 苏州215009)
多杀菌素(spinosad)是一种由刺糖多孢菌生产的次级代谢产物,Spinosyn A(约85%)和Spinosyn D(约15%)是主要的活性成分[1-2]。在功能上,多杀菌素是一种广谱生物杀虫剂,能有效抑制膜翅目、鞘翅目、双翅目病虫害[3];在结构上,多杀菌素是由一个21-碳四环骨架与两个6-碳脱氧糖(D-福乐糖胺和L-鼠李糖)相连而成[4]。 多杀菌素与一些传统化学杀虫剂相比,具备高效性、生物安全性和环境友好性等优点[5]。 我国是一个农业大国,对杀虫剂有着极大的市场需求[6]。 目前,多杀菌素及其衍生物先后三次获得“美国总统绿色化学挑战奖”[7]。但是,我国多杀菌素研究水平较低[8]。因此,通过菌种选育和条件优化等方法来提高多杀菌素的产量具有极其重要的现实意义。
刺糖多孢菌的生长需要必要的碳氮源,对培养基的成分和浓度进行优化能有效提高多杀菌素产量[9]。赵晨等[10]通过部分因子设计、陡坡实验和中心组合设计,将多杀菌素产量由115.1 mg·L-1提至180.6 mg·L-1。张峰等[11]在Plackett-Burman、Box-Behnken 实验基础上,通过神经网络优化培养基,最终将多杀菌素产量提高了145%。铵离子(NH4+)在微生物培养中通常被作为氮素代谢原料,其在细胞内核酸、氨基酸和辅因子等含氮化合物的合成中扮演重要角色[12]。 多项研究表明,NH4+能显著影响大环内脂抗生素的生物合成。 在梅岭霉素的发酵中,发现5 mmol·L-1的硫酸铵能够有效地提升菌体量和产物合成,并且不同浓度的硫酸铵会对相关酶的活性产生影响,以此改变菌体内部代谢通量[13]。 郑梦洁在NH4+对阿维菌素生产影响的研究中发现,铵离子通过影响孢外淀粉酶等一系列酶的活性,影响终产物的合成[14]。 此外,陶文娜通过对刺糖多孢菌铵载体蛋白基因amtS 的异源表达,证明铵载体蛋白能够提高链霉菌中次生代谢产物的产量[15]。 已有文献中,多杀菌素发酵生产常用的氮源一般只有棉籽蛋白或酵母膏等单一有机氮源,而不含硫酸铵等无机氮源[16-18]。 因此,本文首次尝试在培养基中添加硫酸铵,研究铵离子对多杀菌素发酵生产的影响,以期提高多杀菌素产量。
1 材料与方法
1.1 菌株
菌种Saccharopolyspora spinosa 3-37 为本实验室由原始菌株S.spinosa ATCC49460 经抗性筛选得到。
1.2 培养基
平板活化和斜面固体培养基(g·L-1):牛肉浸膏1、酵母提取物1、葡萄糖10、酸水解酪蛋白2、琼脂20,pH 值为7.2,121 ℃灭菌20 min。
种子培养基(g·L-1):葡萄糖15、黄豆饼粉3、淀粉10、大豆蛋白胨25、棉籽蛋白3、MgSO4·7H2O 2,pH 值为7.2,121 ℃灭菌20 min。
发 酵 培 养 基(g·L-1):葡 萄 糖50、棉 籽 蛋 白20、K2HPO4·3H2O 1、NaCl 3、FeSO4·7H2O 0.05、CaCO31,pH 值为7.2,121 ℃灭菌20 min。
补料培养基(g·L-1):棉籽蛋白200、葡萄糖400,pH 值调至6.8。
1.3 培养方法
菌种活化:将甘油保存菌划线接种平板,28 ℃恒温培养7~10 d 至孢子成熟。 成熟单菌落使用接种环划线接种,28 ℃恒温培养7~10 d。
种子培养:用生理盐水冲洗斜面孢子,取1 mL 菌悬液转接种子液(50 mL),28 ℃、220 r·min-1培养3 d。
发酵培养:摇瓶发酵,在250 mL 的摇瓶中装液40 mL,10%接种量(V∶V)接种,28 ℃、220 r·min-1培养7~10 d。
1.4 实验方法
1.4.1 硫酸铵浓度对多杀菌素生产的影响
配制0、0.25、0.50、0.75、1.00、1.50 g·L-1的硫酸铵培养基,测定不同硫酸铵浓度下多杀菌素产量变化。
1.4.2 Plackett-Burman 实验设计
Plackett-Burman 实验设计是一种多因素2 水平的试验设计方法,以相对少的实验组数从中筛选重要因子[19]。 该实验选取葡萄糖、棉籽蛋白、硫酸铵、氯化钠、磷酸氢二钾、硫酸亚铁、碳酸钙作为考察因子,使用Design-Expert 软件设计7 因素2 水平的12 组组合试验,并进行回归分析,得出重要影响因子。
1.4.3 最陡爬坡实验
根据Plackett-Burman 实验得出影响程度最高的3 个因素,根据方程系数的正负大小判断爬坡的方向和步长,设计爬坡实验组。
1.4.4 响应面优化方法
根据上述陡坡实验结果,设计3 因素3 水平的响应面分析表及验证实验,得出最优的培养基浓度配比。
1.4.5 5 L 罐分批和补料分批发酵
将优化培养基配制3 L 置于5 L 发酵罐中,10%接种量(V∶V),30 ℃下进行分批和补料分批发酵。在发酵过程中,0~60 h 维持pH 值为7.2,促进菌体生长;60~180 h 维持pH 值为6.8[20]。
1.5 参数测定
1.5.1 菌体浓度和葡萄糖浓度
用移液枪吸取10 mL 发酵液至10 mL 离心管中,4 000 r·min-1离心15 min,上清液稀释一定倍数,于生物传感分析仪(SBA-40C)测定葡萄糖浓度;沉淀用蒸馏水洗涤2 次,最后将沉淀物置于60 ℃烘箱中烘干称重,计算菌体浓度(DCW)。
1.5.2 多杀菌素含量
取发酵液2 mL,加入等体积甲醇,用漩涡混合器或颠倒充分摇匀,超声破碎细胞15 min,置于暗处过夜保存,4 000 r·min-1离心15 min,上清液过滤处理后,利用高效液相色谱仪(HPLC)进行分析[21],根据峰面积和标准曲线计算多杀菌素产量。
2 结果与分析
2.1 硫酸铵浓度对多杀菌素生产的影响
前期实验发现当硫酸铵浓度大于1.5 g·L-1时,多杀菌素的生产受到抑制。 因此,适当降低硫酸铵浓度进行实验,结果如图1 所示。 由图1 可见,硫酸铵浓度在0~0.75 g·L-1范围内,多杀菌素产量会随着硫酸铵浓度的增加而增加,当硫酸铵浓度达到0.75 g·L-1时,多杀菌素产量达到最大值71.0 mg·L-1,较未添加硫酸铵培养基产量(65.0 mg·L-1)提高了9.2%。
上述结果说明低浓度(0.75 g·L-1)的硫酸铵对多杀菌素的产量提高有一定的促进作用。 在此基础上,后续实验对包含硫酸铵成分的发酵培养基继续进行优化。
2.2 Plackett-Burman 设计筛选影响多杀菌素产量的显著因素
在上述实验结果和相关文献的基础上,将培养基的7 种成分作为考察因子。 使用Plackett-Burman 实验设计,选用高(+1)和低(-1)两水平,根据实验结果筛选显著因子,见表1、表2、表3(表格中:A 为葡萄糖,B 为棉籽蛋白,C 为硫酸铵,D 为氯化钠,E 为磷酸氢二钾,F 为硫酸亚铁,G 为碳酸钙)。
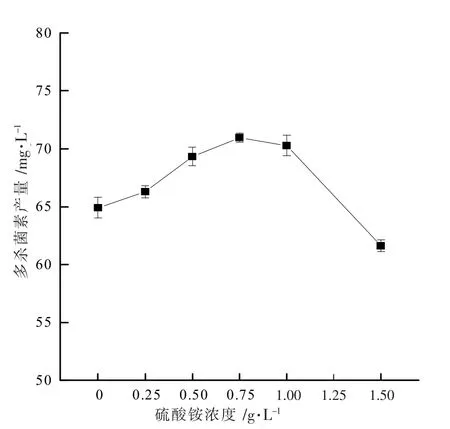
图1 硫酸铵对多杀菌素产量的影响

表1 Plackett-Burman 设计不同因子及水平 单位:g·L-1
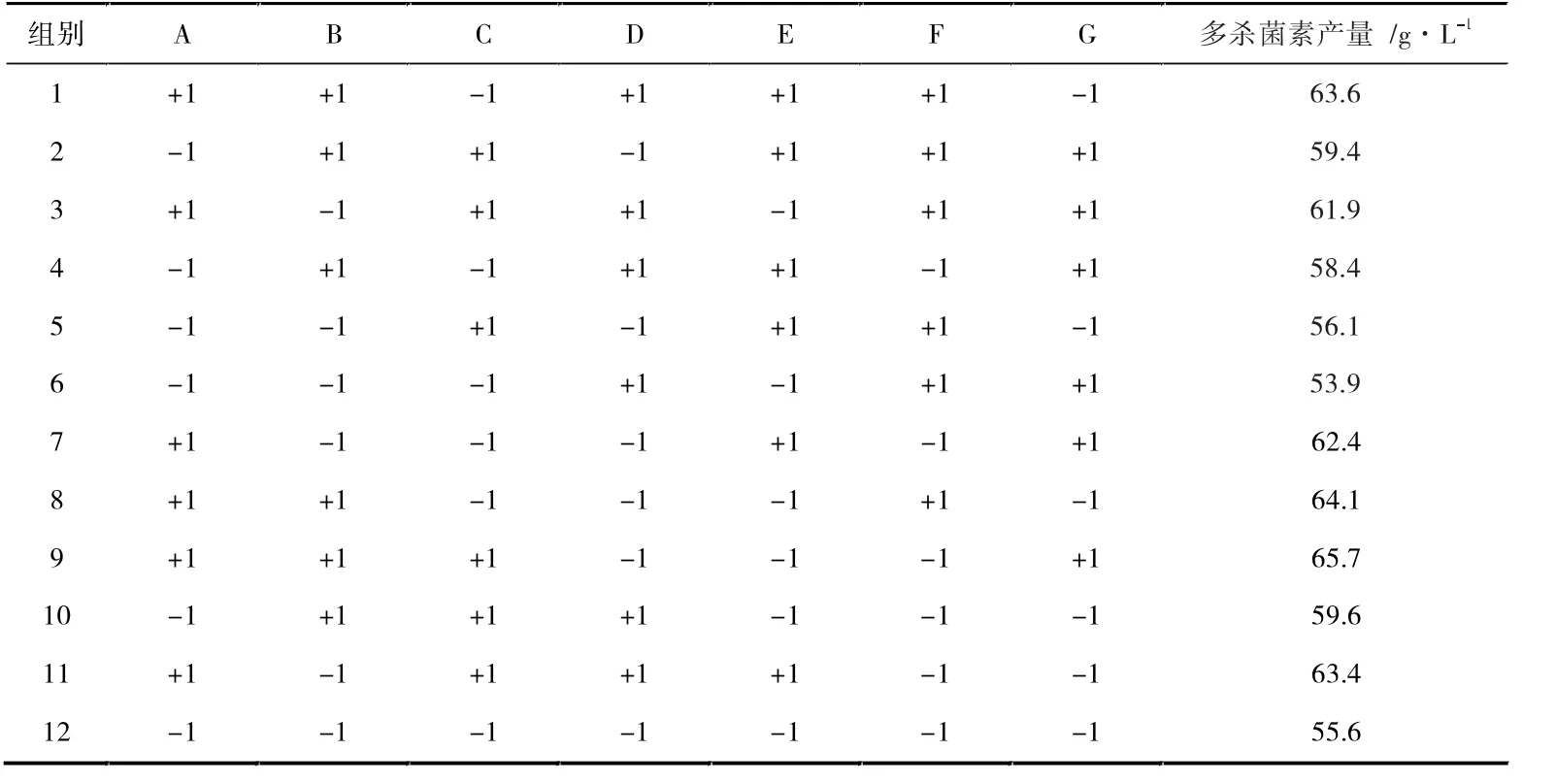
表2 Plackett-Burman 实验设计与结果
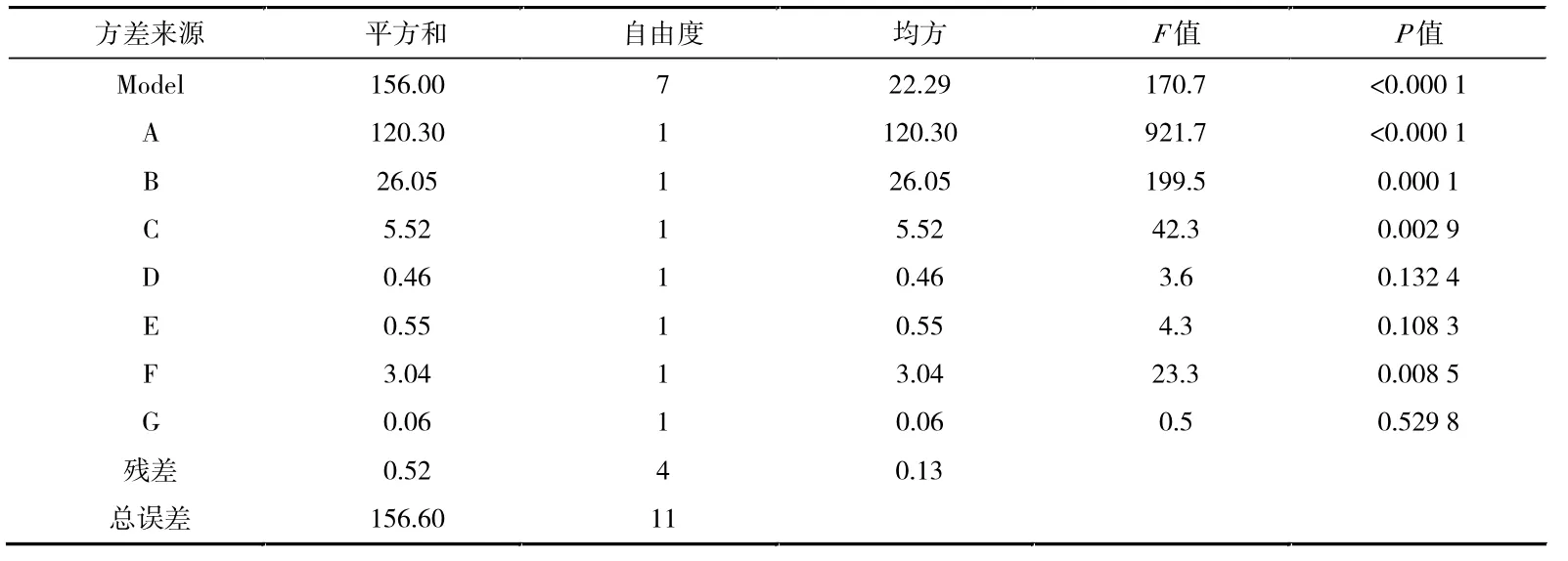
表3 Plackett-Burman 实验设计方差分析表
由表2 数据可得线性回归方程:Y=60.33+3.17A+1.47B+0.68C-0.20D+0.22E-0.50F-0.072G。 表3 数据表明,在7 个因素中,葡萄糖、棉籽蛋白和硫酸铵对多杀菌素的产量影响高于其他几种,且都为正效应,可以在后续试验中适当增加浓度,而其余因素根据系数正负,选择相应的最适水平[22]。
2.3 最陡坡实验
Plackett-Burman 设计只能判断因素重要性,但缺乏对因素最优水平的确定,而响应面只能预测相邻区域产量[23]。 因此,必须根据方程系数的正负大小设计实验组,寻求最大产量区域,具体实验设计见表4。

表 4 最陡爬坡实验设计及其结果
由于3 种因素的系数都为正值,应当逐渐增加浓度。表4 的结果说明,随着3 种因素的浓度逐渐增加时,多杀菌素的产量会呈现先增大后缩小的趋势,当葡萄糖50 g·L-1、棉籽蛋白25 g·L-1、硫酸铵1.0 g·L-1时,产量最高。 因此,将此条件作为响应面实验中心点。
2.4 Box-Behnken 响应面法分析
2.4.1 模型建立
以上述实验结果为基础,挑选了铵离子、葡萄糖和棉籽蛋白3 个因素,采用3 因素3 水平设计中心组合实验,将各因素中多杀菌素产量最高的条件标记为“0”,低于和高于此条件的数值分别标记为“-1”和“1”,见表5。
使用Design-Expert 软件将上述因素水平生成15 个组合试验[24],从而考虑不同因素的相互作用和对多杀菌素产量的影响,见表6。
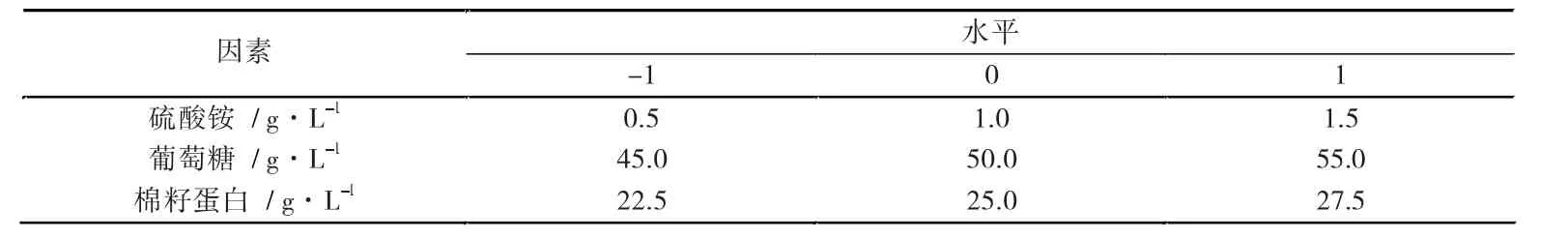
表5 多杀菌素响应面的因素水平设计
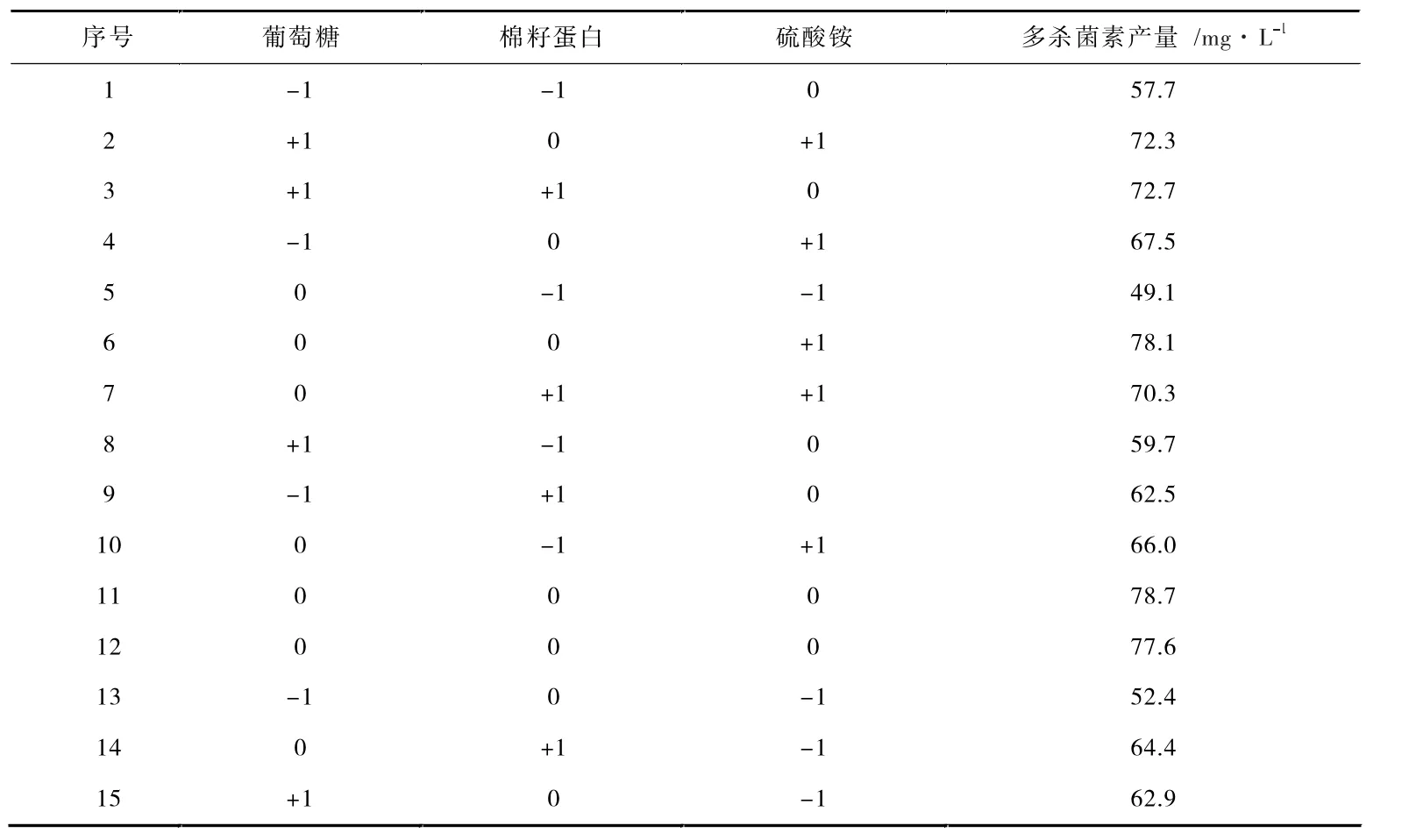
表6 Box-Behnken 实验设计及结果
2.4.2 回归模型的拟合方差分析
使用Design-Expert 软件分析实验结果,得到回归模型方差分析表(见表7)。

表7 回归模型方差分析
通过软件分析得到多杀菌素产量的回归方程为
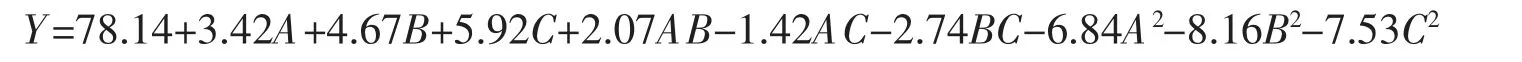
由表7 中的数据可知,此模型P<0.000 1,说明此模型达到显著水平;模型的失拟项P=0.340 1>0.05,说明了此模型的失拟项并不显著,具有应用意义。此外,模型可信度相关系数R2=0.997 9,说明模型的相关度比较好,能够很好地预测3 因素与多杀菌素产量之间的关系;校正系数R2Adj=0.994 1,表明了该模型与实验拟合度较高,可以用来做理论值推算,可信度较高。
2.4.3 响应面法优化
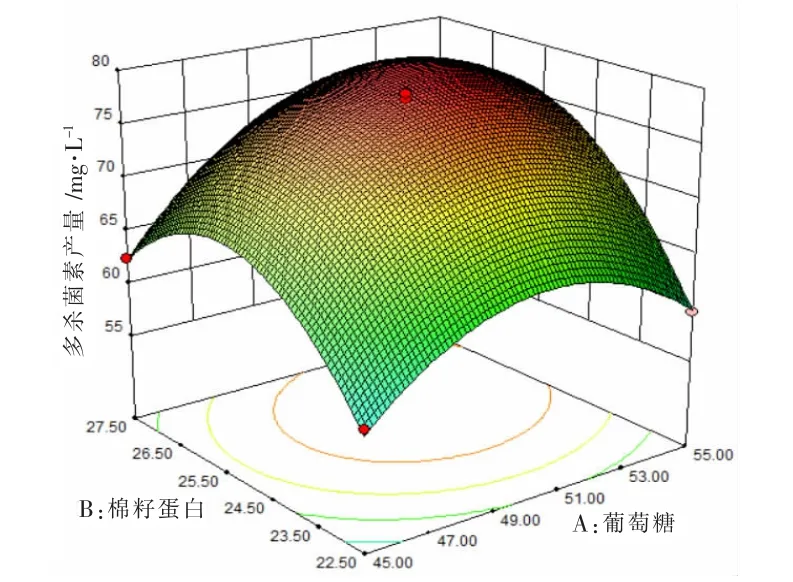
图2 葡萄糖和棉籽蛋白对多杀菌素产量影响的响应面分析图
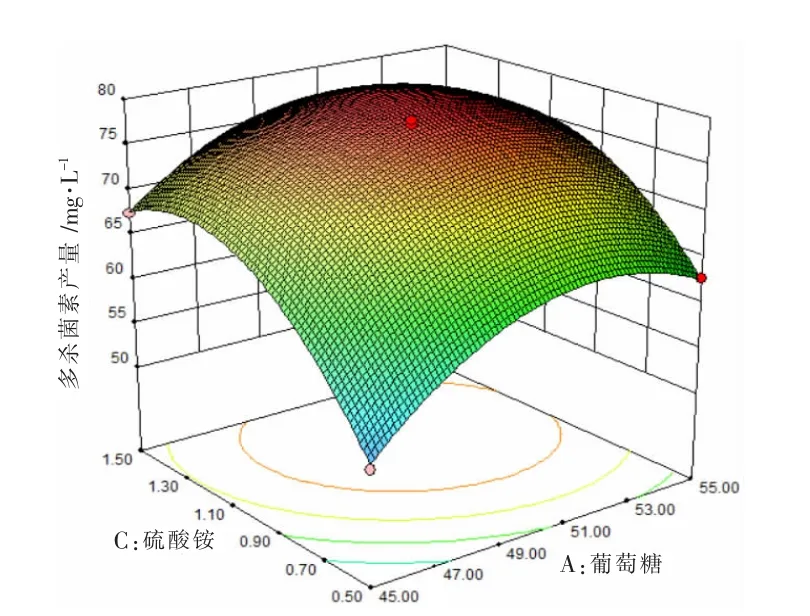
图3 葡萄糖和硫酸铵对多杀菌素产量影响的响应面分析图
根据回归方程对硫酸铵、葡萄糖、棉籽蛋白3 个因素进行响应图绘制(如图2、图3、图4 所示),每个图代表其中两个因素对多杀菌素产量影响的交互作用。 由图2、图3、图4 可知,多杀菌素的产量会受到每个因素的影响,当其中某个因素含量不变,另一成分含量增加,多杀菌素产量也会随之增加,当该成分超过某值时,多杀菌素会随着其成分的增加而降低。以图3 为例,当硫酸铵浓度一定时,多杀菌素产量会随着葡萄糖浓度增加而增加, 但当葡萄糖浓度超过51.3 g·L-1时,多杀菌素产量会呈现下降趋势。 反之,保证葡萄糖浓度一定,硫酸铵也有相同趋势。 同理,发现图2 和图4 也有相同规律。
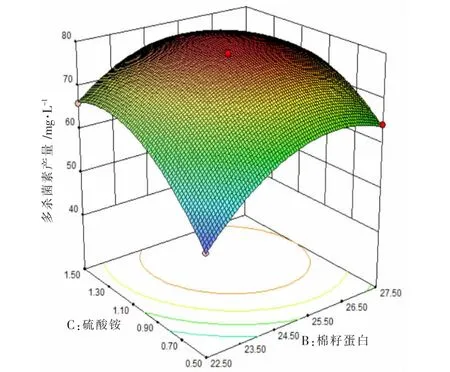
图4 棉籽蛋白和硫酸铵对多杀菌素产量影响的响应面分析图
2.4.4 优化培养基的验证
在上述实验基础上,通过软件分析,预测3 因素最佳浓度配比以及最高多杀菌素产量,结果如下:葡萄糖51.3 g·L-1,棉籽蛋白25.7 g·L-1,硫酸铵1.16 g·L-1,此条件下多杀菌素的预测产量为80.2 mg·L-1。为了验证正确性,在此条件下做3 组平行实验,平均值80.7 mg·L-1,与多杀菌素理论值接近,模型可信,具有实用价值。
2.5 5 L 罐分批和补料分批发酵
刺糖多孢菌在5 L 发酵罐中的分批发酵过程参数变化如图5 所示。 从图5 可以看出,60 h 前,葡萄糖消耗速率较快,菌体生长旺盛,而多杀菌素产量较低,碳氮源的消耗主要用于菌株生长。 80 h 后,菌体生长进入平稳期,并在108 h 时,菌体量达到峰值26.9 g·L-1,葡萄糖由于菌体生长的大量消耗,其消耗速率开始降低,浓度降至低水平。 同时,在积累充足菌体量后,多杀菌素的生产速率急剧上升,在144 h 时,产量达到最大90.5 mg·L-1,较摇瓶发酵提高了12.1%。 但是,由于前期大量碳氮源用于菌株生长,108 h 后碳氮源浓度就处于较低水平,菌体量也由于营养限制呈下降趋势,限制了产物的积累。
为了解决分批发酵中后期碳氮源不足的问题,在发酵96 h 时,采用连续补料流加葡萄糖和棉籽蛋白混合液,维持葡萄糖浓度在10 g·L-1左右,结果如图6 所示。 图6 表明,连续补料解除了营养物质匮乏的限制,在216 h 和240 h 时,菌体量和多杀菌素产量达到最高,分别为42.1 g·L-1和158.3 mg·L-1,较分批发酵提高了56%和74.8%,产量较摇瓶发酵提升了96.2%。 上述结果表明,铵离子确实可以促进多杀菌素的合成。
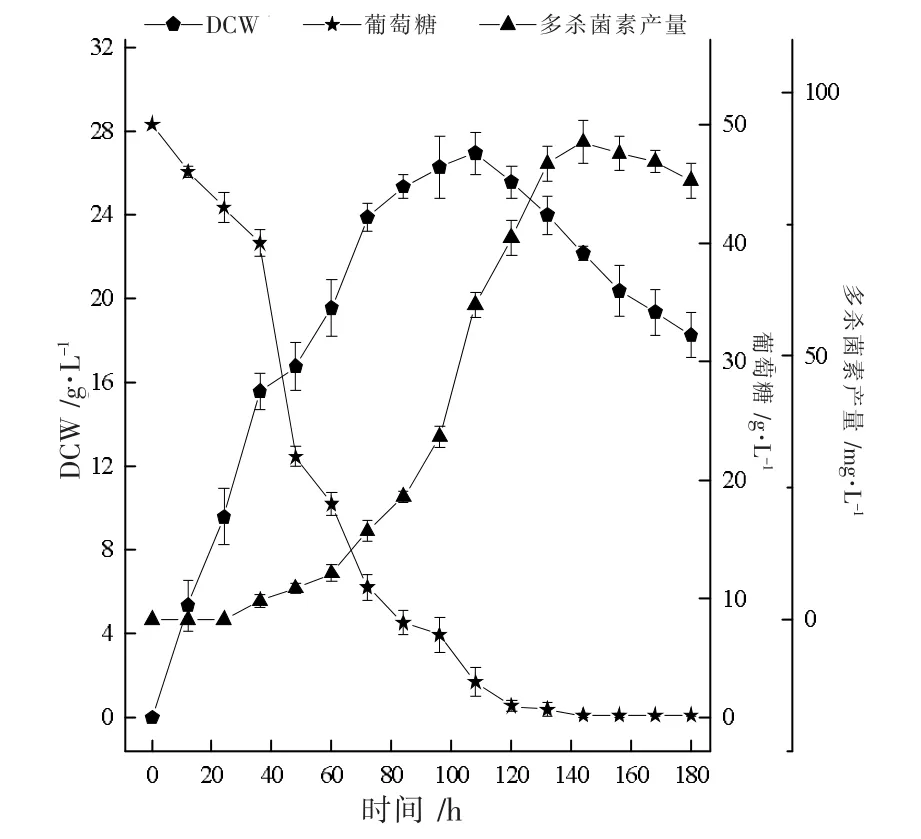
图5 5 L 罐中多杀菌素分批发酵生产动力学曲线
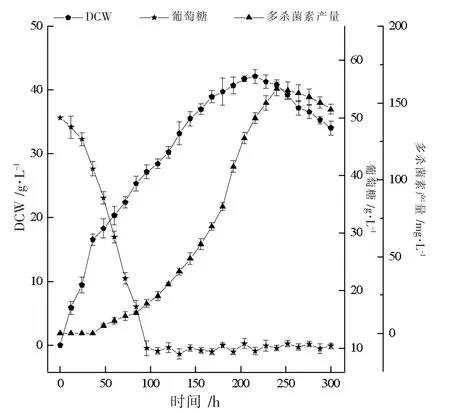
图6 5 L 罐中多杀菌素补料分批发酵生产动力学曲线
3 结语
通过在发酵培养基中加入0.75 g·L-1的硫酸铵, 多杀菌素摇瓶发酵产量提高9.2%, 后经Plackett-Burman 实验设计和响应面优化后,优化培养基配方为(g·L-1):葡萄糖51.3、棉籽蛋白25.7、硫酸铵1.16、NaCl 3、K2HPO4·3H2O 1.5、FeSO4·7H2O 0.05、CaCO31,在此优化条件下多杀菌素产量为80.7 mg·L-1,较未添加硫酸铵培养基提升13.7%。 接着,在5 L 罐中利用该优化培养基进行分批和补料分批发酵培养,最终多杀菌素产量为90.5 mg·L-1和158.3 mg·L-1,分别较摇瓶发酵产量提升了12.1%和96.2%。 相关研究果表明,铵离子能通过影响链霉菌菌株内某些氨基酸脱氢酶和酰基转移酶的活性,以此改变相关氨基酸的代谢和前体合成水平,最终影响抗生素的生物合成[25]。在本研究中,适当添加低浓度硫酸铵有效地促进了多杀菌素生产,其原因可能是低浓度的硫酸铵促进了前体乙酰CoA 和丙酰CoA 的合成。 此外,低浓度的铵离子缩短刺糖多孢菌生长周期、提高菌体生长水平的能力[26],也可能是促进多杀菌素产量增加的原因。 本文为刺糖多孢菌培养基研究提供了新思路,可为提高多杀菌素的产量提供有益的参考。
猜你喜欢
中国调味品(2022年10期)2022-10-13
中国饲料(2022年11期)2022-06-20
当代水产(2022年1期)2022-04-26
农家致富顾问·上半月(2021年6期)2021-12-26
食品界(2021年12期)2021-12-20
农民致富之友(2020年24期)2020-09-02
食品安全导刊·中旬刊(2020年6期)2020-07-28
食品界(2019年2期)2019-03-10
食品界(2019年2期)2019-03-10
江苏农业科学(2017年6期)2017-05-11
