
玉米初级三体的鉴定及传递率研究
2019-09-21 02:45余水情赵永康杨克诚唐祈林
核农学报 2019年11期
秦 燕 余水情 赵永康 杨克诚 唐祈林,*
(1 四川农业大学玉米研究所,四川 成都 611130;2 成都市农林科学院作物研究所,四川 成都 611130;3 宜宾市屏山县人才交流开发服务中心,四川 宜宾 645350)
初级三体的超数染色体是一个正常染色体,配子的染色体数为n,其初级三体就有n种类型。初级三体材料是进行基因定位和连锁图谱构建的基础材料,利用三体分析可以将基因、分子标记和连锁群定位到特定染色体上[1]。目前已在玉米[2]、水稻[3]、番茄[4]、曼陀罗[5]、大豆[6]、谷子[7]、菜薹[8]、棉花[9]、结球甘蓝[10]、普通荞麦[11]等多种作物中选育出成套或部分三体,并利用三体遗传分析将水稻香气基因[12]、水稻光敏核不育基因[13]、大豆rps-1k基因[14]、谷子雄性核不育基因[15]、谷子胚乳糯性基因[16]、谷子矮杆基因[17]、大白菜雄性核不育基因[18]定位到了相应的染色体上。关于三体的形态学和细胞学鉴定,Blackslee等[5]首次从曼陀罗中分离出自然发生的初级三体,并根据曼陀罗的蒴果形态对三体进行了分类。Mcclintock[2]研究表明,玉米的10个初级三体植株虽然较其对应的二倍体植株弱小,但三体与二倍体间差异较小;张成合等[10]对合成的一整套结球甘蓝三体进行了形态学和细胞学鉴定,结果表明初级三体在株高、株型、叶形、花蕾大小、开花期等方面表现出一定的差异。申书兴等[19]和张成合等[20]分别对大白菜和结球甘蓝的初级三体n+1配子传递率进行研究,表明多数n+1配子可以通过雌雄配子传递,且不同植物和不同染色体的传递率不同。目前虽已有关于玉米初级三体的报道,但不同来源的初级三体形态特征及传递率的差异较大,而关于玉米初级三体细胞学、形态学鉴定及传递率的研究尚鲜见报道。本研究以引进的玉米初级三体为试验材料,对其初级三体进行形态学、细胞学鉴定及传递率研究,以期为玉米初级三体后期应用奠定一定的理论基础。
1 材料与方法
1.1 材料
供试材料为从美国引进的一套玉米初级三体Trisomic1~Trisomic10(T1~T10),三体染色体数目为2n+1=21,二倍体染色体数目为2n=20,各三体对应的二倍体简写为T1(2n)~T10(2n)。每个三体材料含有1个标记基因,具体信息详见表1。
1.2 方法
1.2.1 染色体计数方法 2016年8月秋将引进的玉米初级三体在云南元江进行自交繁种,2017年3月将繁殖的三体分批发芽,单株编号取根,采用根尖染色体计数法[21-22]鉴定染色体数目。为保证种子发芽率和染色体数目鉴定的准确性,将种子浸泡6 h,在发芽盒中铺细沙放入培养箱中27℃催芽,待根长至1~1.5 cm时,取根尖,用饱和α-溴萘溶液预处理3 h,再用新鲜固定液[甲醇∶冰乙酸=3∶1(v/v)]固定2~3 h。用2%纤维素酶与1%果胶酶混合(体积比为1∶1)于37℃酶解2.5~3 h,滴加适量改良品红染液、压片,在OLYMPUS BX41相差显微镜(OLYMPUS,日本)下鉴定染色体数目。
1.2.2 核型分析方法 核型分析采用李懋学等[23]确定的标准进行,染色体的相对长度、臂比及类型按Levan等[24]的命名系统,染色体相对长度系数采用Kuo等[25]的方法,核型分类参照Stebbins[26]的标准进行计算。用Image-ProPlus5.51(Media Cybernetics, Inc.)专业图像分析软件进行染色体长度的测量及染色体核型的配对分析。分别选取5个典型的中期细胞进行测量,取平均值制成染色体参数表,并选择具有代表性的分裂相排成核型图。
1.2.3 形态调查方法 鉴定染色体数后,对10个材料中的三体(2n+1)与二倍体(2n)分别进行形态学调查,包括株高、穗位高、雄穗特性(分枝数、穗长),雌穗特性(粒色、轴色、穗长、穗行数、行粒数、百粒重),叶片特性(叶片总数、穗三叶叶长、叶宽),花粉育性(花粉可染率),花粉育性采用碘染法[27]进行鉴定。
1.2.4 传递率及结实率统计 10个玉米初级三体染色体数目鉴定后,2017年配置初级三体自交及其与二倍体的正反交组合:T(2n+1)×T(2n+1)、T(2n+1)×T(2n)、T(2n)×T(2n+1)。2018年统计所得组合的结实数并将每个组合的籽粒逐一进行染色数目鉴定,按照公式计算结实率和额外染色体的传递率:
结实率=结实的籽粒数/(行数×行粒数)×100%
(1)
传递率=三体株数/鉴定总株数×100%
(2)。
1.3 数据统计与分析
10个玉米材料的三体与二倍体个体各调查5株,用 DPS 7.05 软件对各材料内三体及其对应的二倍体间的差异显著性进行分析。
2 结果与分析
2.1 染色体数及核型
对10个玉米材料共计1 386个单株进行染色体数目鉴定,由表2可知,10个玉米材料中均有染色体数为2n+1=21的三体出现,且不同材料出现三体频率存在差异,10个材料三体出现频率均值为15.6%,变幅为3.4%~25.0%。
由图1可知,T1~T10增加的1条染色体的形态及染色体相对长度(relative chromosome length,RL)、臂比(arm ratio,AR),着丝粒位置(position of centromere,PC)等参数均分别与其第1~第10对染色体接近,表明T1~T10是增加玉米第1~第10条染色体的玉米初级三体。由表3可知,其核型主要由中部着丝点染色体(metacentric chromosome,m)和近中部着丝点染色体(submetacentric chromosome,sm)组成,第6号染色体具有随体,无B染色,体核型类型均属于较对称的B型。

表2 玉米单株的三体频率Table 2 The frequency of trisomics in the single plant of maize
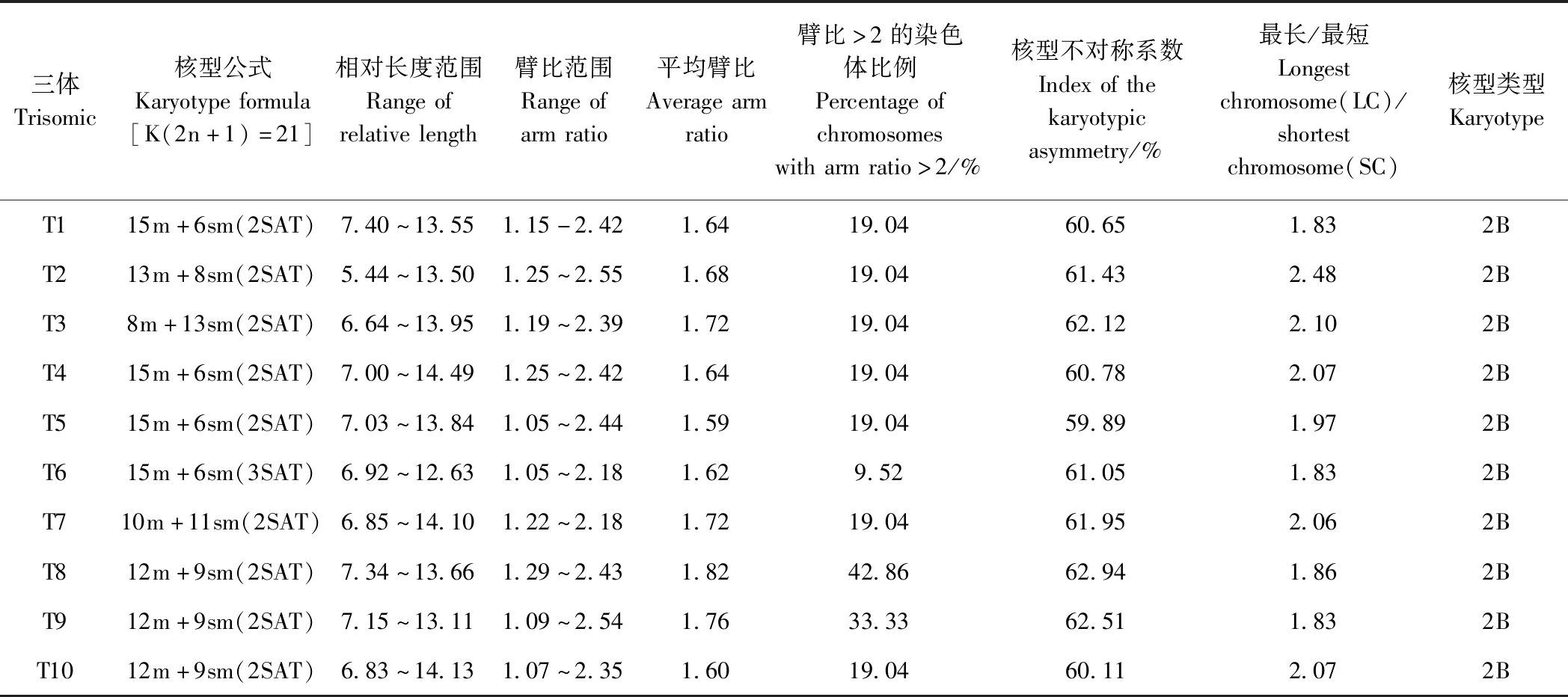
表3 玉米初级三体的核型参数Table 3 Karyotype parameters of maize primary triromics

图1 玉米三体(T1~T10)核型图与染色体形态Fig.1 Chromosome morphology and karyogram of maize trisomics (T1-T10)
2.2 形态学鉴定
由表4可知,在玉米植株形态上,除T3外,其余9个三体的株高、穗位高、叶片数、叶长、叶宽均低于对应的二倍体植株,但仅T1的叶长与T1(2n)的差异达到极显著,其余三体与其对应二倍体间差异均不显著;在雌穗性状上,10个三体的雌穗长度、穗行数、行粒数及百粒重均小于对应的二倍体,其中T6雌穗长度与T6(2n)差异达到极显著;在雄穗性状上,10个三体的雄穗长度、雄穗分枝数及花粉可染率(花粉育性)均小于对应的二倍体,其中T3、T5、T6、T7、T9的花粉育性与其对应的二倍体之间的差异均达到极显著水平。综上表明,除T3的植株较二倍体高外,其余三体的各性状均较二倍体小,即附加1条染色体导致三体的生活力及生长势大多弱于其二倍体;而各三体与二倍体间的差异也随附加的染色体不同而存在差异,附加的染色体不同,对各性状的影响也不同。t检测结果表明,T1可通过测量叶片长度对三体植株进行初步鉴定,T3、T5、T7、T9可通过花粉育性检测对三体植株进行初步鉴定,T6可通过雌穗长度、育性检测对三体植株进行初步鉴定。
对10个材料中易观察的标记基因表现型进行观察,结果表明,T1和T1(2n)中均有黄铜色籽粒出现;T4和T4(2n)均有皱缩籽粒出现;T8和T8(2n)均出现幼苗黄白,成株后缓慢变绿现象;T9和T9(2n)均有糯质玉米出现;T10和T10(2n)籽粒表面均有紫色斑块出现。综上,植株标记基因的性状表现与该植株是否为三体没有必然联系。
2.3 初级三体的传递率和结实率
由表5可知,10个自交材料的初级三体的结实率为41.3%~55.7%,平均结实率为50.3%,额外染色体传递率为3.4%~25.0%,平均传递率为15.6%,表明,初级三体自交能够结实,额外染色体能够通过自交传递。初级三体T1、T2、T4、T5、T6、T9作母本与二倍体杂交材料的结实率为52.7%~69.1%(T3、T7、T8、T10与二倍体正交组合未配置成功),平均结实率为61.3%,额外染色体传递率为1.7%~21.7%,平均传递率为9.4%,表明初级三体与二倍体正交均能结实,且结实率高于自交组配方式,但额外染色体的传递率却较自交组配方式下降了6.2个百分点;初级三体作父本与二倍体杂交材料结实率为72.9%~83.4%,平均结实率为78.4%,额外染色体传递率为0~6%,有4个三体的传递率为0,平均传递率为1.8%,表明初级三体作父本与二倍体杂交虽然结实率最高,但额外染色体传递率明显下降,通过二倍体花粉传递超数染色体的概率较小。综上,通过初级三体自交这种方式来进行三体额外染色体的传递,能够达到较高的额外染体传递率和适中的结实率。

表4 玉米三体与二倍体各性状均值及t检验结果Table 4 The mean value and t test results of morphological characters between the trisomic and diploid of maize
注*和**分别表示在0.05和0.01水平上显著。
Note:*and**indicate significant difference at 0.05 and 0.01 level, respectively.
注:()中为三体株数/鉴定总株数,“-”表示未收集到数据。
Note:()means number of 2n+1 plant/number of identified plant. ‘-’Indicates that no data had been collected。
3 讨论
初级三体的鉴定是三体利用的基础,鉴定初级三体主要通过形态学鉴定、有丝分裂染色体核型分析、粗线期核型分析、染色体分带核型分析、分子标记鉴定等方法[28]。研究表明,对初级三体染色体进行识别鉴定最直接的方法是有丝分裂染色体核型分析,但由于最初获得三体的作物是小染色体物种,利用有丝分裂较难区分,有研究者进一步应用粗线期的染色体进行核型分析[29],目前已有应用分子标记对水稻[30]、大白菜[31]等作物进行初级三体鉴定的报道,但该分析方法鉴定结果易受环境因素干扰。本研究结果表明,部分玉米初级三体与其二倍体的一些性状间差异达到显著水平,从理论上说,可以通过差异显著的性状对三体进行鉴定,但由于形态学性状受环境、气候等自然条件影响较大,鉴定结果可信度较低。因此在三体鉴定中,形态学鉴定不能作为最可靠的鉴定手段,需与细胞学鉴定或分子标记鉴定结合。
本研究中,除T3的株高、穗位高及雄穗长度略高于对应的二倍体植株外,其余9个三体各性状均小于对应的二倍体,这与Mcclintock[2]的研究结果一致,其原因为三体增加一条额外染色体,可能会导致其生活力及生长势都弱于二倍体植株,对于T3材料出现的异常现象的原因尚不明确。此外,初级三体系植株标记基因的性状表现与植株是否为三体没有必然联系,可利用分子标记等手段对初级三体标记基因的表达进一步深入研究。
额外染色体的传递率是利用三体进行遗传分析的重要参数。一般通过(2n+1)×2n、2n×(2n+1)这2种组配模式来测定额外染色体的传递率。申书兴等[19]研究发现大白菜额外染色体通过雌配子传递率为8.69%~29.42%,通过雄配子传递率为3.53%~23.53%;张成合等[20]发现结球甘蓝额外染色体通过雌配子传递率为5.36%~42.37%,通过雄配子传递率为1.54%~13.04%;罗双霞等[1]发现芥蓝额外染色体通过雌配子传递率为10.15%~27.63%,通过雄配子传递率为4.91%~11.86%,即额外染色体通过雌配子传递率一般高于雄配子的传递率。本研究增加了(2n+1)× (2n+1)三体自交模式测定传递率,该模式传递率为3.4%~25.0%,高于仅通过雌配子(1.7%~21.7%)和仅通过雄配子(0~6.0%)的传递率,表明通过玉米初级三体自交可获得更高的传递率,其原因可能是雌、雄配子同时出现n+1配子,从而增加了2n+1个体出现的几率。
4 结论
本研究结果验证了T1~T10确实是分别增加玉米第1~第10条染色体的玉米初级三体;玉米初级三体的鉴定可采用细胞学和形态学鉴定相结合方法;额外染色体通过三体自交的传递率为3.4%~25.0%,高于仅通过雌配子(1.7%~21.7%)和仅通过雄配子(0~6.0%)的传递率,通过自交对该初级三体系的额外染色体进行传递,能够达到较高的传递率和适中的结实率。对引进的初级三体进行细胞学、形态学鉴定及传递率研究为该套初级三体的进一步利用提供了应用基础,具有十分重要的意义。下一步可利用该套玉米初级三体进行相关基因定位研究、染色体或基因的重复效应研究等。
猜你喜欢
生物学通报(2021年11期)2021-09-28
上海农业学报(2019年5期)2019-11-08
江苏农业科学(2017年16期)2017-10-27
西南农业学报(2017年7期)2017-08-11
现代农业科技(2017年8期)2017-06-10
上海农业学报(2017年3期)2017-04-10
中学生物学(2017年1期)2017-03-23
中国医药导报(2015年26期)2015-02-28
中国果业信息(2015年6期)2015-01-23
草原与草业(2013年1期)2013-12-03
