
外源褪黑素对镉胁迫下豌豆种子萌发、幼苗抗性生理及镉含量的影响
2019-09-21 02:45王艳芳王悦华温烜琳蔡慧英郑晓蕾陈彤彤
核农学报 2019年11期
李 冬 王艳芳 王悦华 温烜琳 蔡慧英 郑晓蕾 陈彤彤 刘 领
(河南科技大学农学院,河南 洛阳 471023)
近年来随着我国工农业和城市化的快速发展,重金属造成的土壤污染和水污染已经成为全球重点关注的问题。镉(Cd)污染是最常见的重金属污染之一。Cd不是植物生长和发育必需的元素,但其在土壤中具有较强的化学活性和亲水特性,易被植物吸收积累,且存留在可食用部分,进而通过食物链富集在人体中,毒害人体器官和生殖系统及免疫系统,危害人体健康[1-2]。Cd被植物吸收后,易导致植物遗传物质损伤和变异,轻度胁迫导致植物叶片干枯萎黄,根茎缩短,侧根数量减少,降低营养元素的吸收;重度胁迫则会减少叶绿素含量,扰乱水分平衡,抑制抗氧化酶活性,引起活性氧积累,细胞膜透性降低,导致细胞损伤,进而显著抑制植物生长[3-4]。Cd对植物的伤害首先表现在对种子的萌发和幼苗生长的影响,而种子萌发时期的生长状况直接影响作物的生长和产量[5]。因此,研究作物种子萌发及幼苗生长阶段受Cd污染的影响尤为重要。

豌豆(Pisumsativum)豆科豌豆属一年生或越年生草本植物,世界第二大食用豆类,是高品质蛋白质的重要来源,具有较高的营养价值和防癌、抗癌等作用,因其具有较高的固氮能力和较强的适应性,在全国各地均有大面积栽培[15]。目前有关外源褪黑素对重金属胁迫下植物生长发育的影响已有研究,而对Cd胁迫下豌豆种子萌发及幼苗生长影响的研究鲜见报道。本研究以豌豆为材料,采用褪黑素浸种处理豌豆种子,探究外源褪黑素在缓解豌豆Cd胁迫方面的生理机制,以期为提高作物对重金属胁迫的耐受性提供科学依据。
1 材料与方法
1.1 试验材料
供试豌豆品种为中豌6号,甘肃利源农业发展有限公司。供试外源褪黑素(MT),美国SIGMA-ALDRICH公司。供试模拟镉胁迫制剂为氯化镉(CdCl2·2.5H2O,优级纯),上海国药集团化学试剂有限公司。
1.2 主要仪器与设备
GZX型光照培养箱,北京中兴伟业仪器公司;NexION300-X型电感耦合等离子体质谱仪,美国PerkinElmer公司。
1.3 试验设计
试验采用双因素随机试验设计,设置Cd胁迫和外源褪黑素处理2个因素,采用培养皿滤纸法进行豌豆种子萌发试验,Cd胁迫设3个水平,分别为:Cd0(添加去离子水,Cd2+浓度为0 μmol·L-1)、Cd10(添加氯化Cd溶液,Cd2+浓度为10 μmol·L-1)、Cd100(添加氯化镉溶液,Cd2+浓度为100 μmol·L-1)。每个镉胁迫水平下设置5个浓度的褪黑素浸种处理,分别为:MT0(去离子水浸种,不加褪黑素溶液)、MT50(50 μmol·L-1褪黑素浸种)、MT100(100 μmol·L-1褪黑素浸种)、MT200(200 μmol·L-1褪黑素浸种)和MT400(400 μmol·L-1褪黑素浸种),共组合为15个处理,每个处理重复3次,每个重复1皿。
挑选大小一致、籽粒饱满的豌豆种子,用3%次氯酸钠(NaClO)消毒10 min,然后用自来水冲洗5次,再用蒸馏水冲洗5次,吸水纸吸干种子表面水分,在室温下用不同浓度褪黑素溶液避光浸种24 h后均匀排列在垫有2层滤纸、直径9 cm的培养皿中,每皿30粒豌豆种子,最后分别向培养皿中加入预先用去离子水和水合氯化镉(CdCl2·2.5H2O)配置的不同Cd2+浓度的溶液10 mL,加盖后置于光照培养箱中培养。光照培养箱设定光照时间为12 h·d-1,昼/夜温度25℃/15℃,光照强度4 000 Lx,培养期间每天定时观察和记录种子的发芽情况,并用称重法补充蒸发的水分。
1.4 测定项目与方法
1.4.1 萌发指标测定 培养时期每天记录豌豆种子萌发的数量,以下胚轴突破种皮长度超过种子自身长度一半视为发芽标准,第3天统计种子的发芽势,第8天统计豌豆种子的发芽率;第8天从各培养皿中随机选取10株豌豆幼苗测定根长、芽长、根鲜重和芽鲜重。按照公式计算发芽势和发芽率:
发芽势=3 d内发芽的种子粒数/供试种子数×100%
(1)
发芽率=8 d内发芽的种子粒数/供试种子数×100%
(2)。
1.4.2 生理指标测定 取萌发8 d的幼芽,于-80℃超低温保存,用于生理指标的测定。MDA含量采用硫代巴比妥酸(thiobarbituric acid,TBA)溶液法测定[16];SOD、POD和CAT活性参照高俊凤[17]的方法测定。
1.4.3 镉含量的测定 随机选取豌豆幼苗,进行烘干粉碎后,加入10 mL优级纯HNO3-HClO4(体积比为 4∶1)消煮,用电感耦合等离子体质谱仪测定Cd含量。
1.5 数据处理
采用Microsoft Office Excel 2016和SPSS 22.0软件进行数据处理和分析,试验均测定3次重复,结果以平均值±标准误表示;采用Origin 9.0软件进行绘图。
2 结果与分析
2.1 镉胁迫下外源褪黑素对豌豆种子发芽势的影响
由图1可知,在同一浓度外源MT处理下,随着Cd2+浓度的增加,豌豆种子的发芽势均逐渐降低。在未添加MT处理时,10和100 μmol·L-1Cd2+浓度的发芽势分别较0 μmol·L-1Cd2+浓度处理降低了40.73%和56.36%,说明Cd胁迫严重抑制了豌豆种子的萌发。同一Cd2+浓度处理下,随着外源MT浓度的提高,豌豆种子的发芽势表现出先升高后降低的趋势。当Cd2+浓度为10 μmol·L-1时,与未添加MT处理相比,50、100、200和400 μmol·L-1MT处理的发芽势分别提高了34.98%、90.47%、49.47%和28.85%;在100 μmol·L-1Cd2+浓度下,与未添加MT处理相比,50、100、200和400 μmol·L-1MT处理的发芽势分别提高了74.99%、124.97%、99.14%和66.63%,说明在Cd胁迫下,不同浓度外源MT浸种均能显著提高豌豆种子的发芽势,且以100 μmol·L-1MT处理效果最为显著。双因素分析表明,Cd处理与MT处理对豌豆幼苗的发芽势均有极显著影响,而两者的交互作用对发芽势无显著影响。

注:*和**分别表示差异显著(P<0.05)和差异极显著(P<0.01);不同小写字母表示同一Cd水平不同褪黑素处理之间差 异显著(P< 0.05)。下同。Note: * and ** mean significant difference (P< 0.05) and extremely significant difference (P< 0.01), respectively. Different lowercase letters in the same Cd level meant significant difference among different melatonin treatments at 0.05 level. The same as following.图1 不同浓度外源MT对Cd胁迫下豌豆种子发芽势的影响Fig.1 Effects of different concentrations of exogenous melatonin on the germination potential of pea seeds under cadmium stress
2.2 镉胁迫下外源褪黑素对豌豆种子发芽率的影响
由图2可知,同一浓度外源MT处理下,随着Cd2+浓度的增加,豌豆种子的发芽率逐渐降低。在未添加MT处理时,10和100 μmol·L-1Cd2+浓度的发芽率较0 μmol·L-1Cd2+浓度处理分别降低了24.13%和32.76%,说明Cd胁迫严重降低了豌豆种子的发芽率。同一Cd2+浓度处理下,随着外源MT浓度的提高,豌豆种子的发芽率均表现出先升高后降低的趋势。当Cd2+浓度为10 μmol·L-1时,与未添加MT处理相比,50、100、200和400 μmol·L-1MT处理的发芽率分别提高了59.09%、82.94%、64.53%和49.31%;当Cd2+浓度为100 μmol·L-1时,与未添加MT处理相比,50、100、200和400 μmol·L-1MT处理的发芽率分别提高了61.55%、85.92%、63.10%和50.27%,说明Cd胁迫下,不同浓度外源MT浸种均能显著提高豌豆种子的发芽率,且以100 μmol·L-1MT处理效果最为明显。双因素分析表明,Cd处理与MT处理对豌豆幼苗的发芽率的影响均极显著,而两者的交互作用对发芽率无显著影响。

图2 不同浓度外源MT对Cd胁迫下豌豆种子发芽率的影响Fig.2 Effects of different concentrations of exogenous melatonin on the germination percentage of pea seeds under cadmium stress
2.3 镉胁迫下外源褪黑素对豌豆幼苗生长的影响
由表1可知,在同一浓度外源MT处理下,随着Cd2+浓度的增加,芽长、根长、芽鲜重和根鲜重基本表现为逐渐降低的趋势。在未添加MT处理时,与 0 μmol·L-1Cd2+浓度处理相比,10和100 μmol·L-1Cd2+浓度处理的豌豆幼苗的芽长、根长、芽鲜重和根鲜重分别降低了50.75%、65.20%、65.64%、42.93%和64.18%、75.09%、68.16%和59.69%,说明Cd胁迫严重抑制了豌豆幼苗的生长。同一Cd2+浓度处理下,随着外源MT浓度的提高,豌豆幼苗的芽长、根长、芽鲜重和根鲜重均表现出先升高后降低的趋势,且均在100 μmol·L-1MT处理下达到最大值。当Cd2+浓度为10 μmol·L-1时,100 μmol·L-1MT处理的芽长、根长、芽鲜重和根鲜重较未添加MT处理分别提高了127.27%、129.47%、131.71%和83.33%;当Cd2+浓度为100 μmol·L-1时,100 μmol·L-1MT处理的芽长、根长、芽鲜重和根鲜重较未添加MT处理分别提高了139.58%、117.65%、85.09%和92.77%,说明不同浓度外源MT浸种均能明显缓解Cd胁迫下幼苗的损伤,促进幼苗的生长发育,且以100 μmol·L-1MT处理效果最佳。双因素分析表明,Cd处理与MT处理及两者的交互作用对豌豆幼苗的芽长、根长、芽鲜重和根鲜重均有极显著影响。

表1 不同浓度外源MT对Cd胁迫下豌豆芽长、根长、芽鲜重、根鲜重的影响Table 1 Effects of different concentrations of exogenous melatonin on the shoot length, root length, shoot fresh weight and root fresh weight of pea under cadmium stress
注:同列不同小写字母表示差异显著(P<0.05)。**表示差异极显著(P<0.01)。
Note: Different lowercase letters in the same line indicate significant difference at 0.05 level. ** indicates extremely significant difference at 0.01 level.
2.4 镉胁迫下外源褪黑素对豌豆幼芽MDA含量的影响
由图3可知,在同一浓度外源MT处理下,随着Cd2+浓度的增加,豌豆幼芽的MDA含量表现出增加的趋势。在未添加MT处理时,10和100 μmol·L-1Cd2+浓度的MDA含量较0 μmol·L-1Cd2+浓度处理提高了249.62%和438.35%;同一Cd浓度处理下,随着外源MT浓度的增加,豌豆幼芽的MDA含量表现出先降低后升高的趋势。当Cd2+浓度为0 μmol·L-1时,不同褪黑素浸种处理之间无显著性差异;在10 μmol·L-1Cd2+浓度处理下,50、100、200和400 μmol·L-1MT处理的MDA含量分别较未添加MT处理下降了46.74%、57.64%、41.31%和24.03%;在100 μmol·L-1Cd2+浓度处理下,50、100、200和400 μmol·L-1MT处理的MDA含量分别较未添加MT处理下降了32.66%、46.90%、29.92%和16.76%,说明在Cd胁迫下,不同浓度外源MT浸种均能显著降低豌豆幼芽的MDA含量,抑制膜脂过氧化对细胞膜的伤害,对细胞膜的完整具有保护作用。双因素分析表明,Cd处理与MT处理及两者的交互作用对豌豆幼芽MDA含量的影响均极显著。
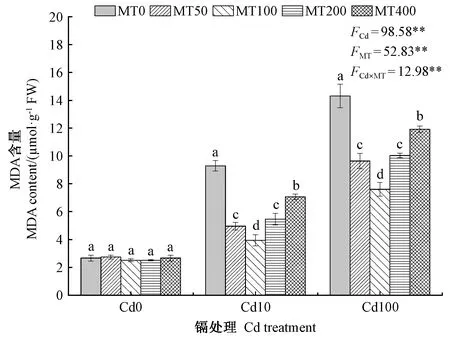
图3 不同浓度外源MT对Cd胁迫下豌豆幼芽MDA含量的影响Fig.3 Effects of different concentrations of exogenous melatonin on the MDA contents of pea plumule under cadmium stress
2.5 镉胁迫下外源褪黑素对豌豆幼芽SOD活性的影响
由图4可知,在同一浓度外源MT处理下,随着Cd2+浓度的上升,豌豆幼芽的SOD活性均逐渐下降。在未添加MT处理时,10和100 μmol·L-1Cd2+浓度的SOD活性较0 μmol·L-1Cd2+浓度处理降低了24.59%和35.08%。在同一Cd2+浓度处理下,随着外源MT浓度的增加,豌豆幼芽的SOD活性均表现出先升高后降低的趋势。当Cd2+浓度为10 μmol·L-1时,50、100、200和400 μmol·L-1MT处理的SOD活性分别较未添加MT处理提高了15.45%、24.17%、14.01%和12.35%;当Cd2+浓度为100 μmol·L-1时,50、100、200和400 μmol·L-1MT处理的SOD活性分别较未添加MT处理提高了15.10%、25.79%、17.35%和9.96%,说明在Cd胁迫下,不同浓度外源MT浸种均能显著提高豌豆幼芽的SOD活性,且以100 μmol·L-1MT处理效果最为显著。双因素分析表明,Cd处理与MT处理及两者的交互作用对豌豆幼芽的SOD活性的影响均极显著。
2.6 镉胁迫下外源褪黑素对豌豆幼芽POD活性的影响
由图5可知,在同一 浓度外源MT处理下,豌豆幼芽的POD活性随着Cd2+浓度增加逐渐降低。在未添加MT处理时,相比0 μmol·L-1Cd2+浓度处理,10和100 μmol·L-1Cd2+浓度处理的POD活性分别降低了25.44%和47.34%。同一Cd浓度处理下,随着外源MT浓度的提高,豌豆幼芽的POD活性均表现为先升高后降低的趋势。当Cd2+浓度为10 μmol·L-1时,50、100、200和400 μmol·L-1MT处理的POD活性分别较未添加MT处理提高了21.79%、40.50%、29.80%和10.91%;当Cd2+浓度为100 μmol·L-1时,50、100、200和400 μmol·L-1MT处理的POD活性分别较未添加MT处理提高了37.95%、62.05%、49.34%和20.71%,说明不同浓度外源MT浸种均能显著提高Cd胁迫下豌豆幼芽的POD活性,且以100 μmol·L-1MT处理效果最为显著。双因素分析表明,Cd处理与MT处理对豌豆幼芽的POD活性的影响均极显著,两者的交互作用对POD活性的影响显著。

图5 不同浓度外源MT对镉胁迫下豌豆幼芽POD活性的影响Fig.5 Effects of different concentrations of exogenous melatonin on the POD activity of pea plumule under cadmium stress
2.7 镉胁迫下外源褪黑素对豌豆幼芽CAT活性的影响
由图6可知,在同一浓度外源MT处理下,随着Cd2+浓度的增加,豌豆幼芽的CAT活性逐渐降低。在未添加MT处理时,10和100 μmol·L-1Cd2+浓度处理的CAT活性较0 μmol·L-1Cd2+处理降低了44.28%和62.66%。同一Cd浓度处理下,随着外源MT浓度的提高,豌豆幼芽的CAT活性均表现为先升高后降低。当Cd2+浓度为10 μmol·L-1时,50、100、200和400 μmol·L-1MT处理的CAT活性分别较未添加MT处理提高了54.37%、76.91%、62.03%和35.48%;当Cd2+浓度为100 μmol·L-1时,50、100、200和400 μmol·L-1MT处理的CAT活性分别较未添加MT处理提高了32.91%、86.40%、48.50%和28.92%,说明不同浓度外源MT浸种均能显著提高Cd胁迫下豌豆幼芽的CAT活性,且以100 μmol·L-1MT处理效果最为显著。双因素分析表明,Cd处理与MT处理及两者的交互作用对豌豆幼芽的CAT活性的影响均极显著。

图6 不同浓度外源MT对镉胁迫下豌豆幼芽CAT活性的影响Fig.6 Effects of different concentrations of exogenous melatonin on the CAT activity of pea plumule under cadmium stress
2.8 镉胁迫下外源褪黑素对豌豆幼苗镉含量的影响
由图7可知,在同一浓度外源MT处理下,随着Cd2+浓度的增加,豌豆幼苗内的Cd含量逐渐增加,当Cd2+浓度为0 μmol·L-1时,豌豆幼苗内Cd含量为0。同一Cd浓度处理下,随着外源MT浓度的增加,豌豆幼苗Cd含量表现出先降低后升高的趋势,但始终低于未添加MT处理。当Cd2+浓度为10 μmol·L-1时,50、100、200和400 μmol·L-1MT处理豌豆幼苗Cd含量分别较未添加MT处理降低了46.72%、57.62%、41.28%和24.00%;当Cd2+浓度为100 μmol·L-1时,50、100、200和400 μmol·L-1MT处理豌豆幼苗Cd含量分别较未添加MT处理降低了34.54%、48.60%、31.66%和12.41%,表明外源MT能减少Cd在豌豆幼苗体内的积累,降低豌豆幼苗Cd含量,且对低浓度Cd2+处理效果更为明显。双因素分析表明,Cd处理与MT处理及两者的交互作用对豌豆幼苗Cd含量的影响均极显著。

图7 不同浓度外源MT对Cd胁迫下豌豆幼苗镉含量的影响Fig.7 Effects of different concentrations of exogenous melatonin on the Cd contents of pea seedlings under cadmium stress
3 讨论
Cd是危害作物生长发育的重金属元素,经植物吸收进入细胞后能够结合酶活性中心或蛋白巯基,取代蛋白反应中心的Ca2+、Mg2+、Fe2+等必需金属离子,引发膜脂过氧化,导致细胞膜遭受损伤,进而抑制植物种子萌发及植株幼苗生长[18-20]。种子萌发与幼苗生长是植物生命的开始,也是植物生命周期中最早接触外界环境、对外界做出反应的重要阶段,其中发芽势和发芽率是衡量作物种子发芽能力的重要指标[21-22]。当植物体内累积了一定量的Cd后,会产生毒害作用,严重阻碍植株对水分、营养元素的吸收转运,导致植株生长缓慢乃至死亡[23]。尚宏芹等[24]研究发现,低浓度Cd2+对桔梗种子萌发作用无显著影响,但会抑制桔梗幼苗生长;高浓度Cd2+会显著抑制种子萌发和幼苗生长,导致植株体内Cd含量明显增加。欧丽等[5]研究发现,低浓度Cd2+胁迫能够促进野茼蒿种子萌发,高浓度Cd2+胁迫则显著抑制其种子萌发。刘自力等[25]研究发现,外源褪黑素可能通过影响IRT1(iron-regulated transporter 1)、IRT2(iron-regulated transporter 2)的转录来影响小白菜对Cd的吸收,降低植株幼苗Cd含量。本研究结果表明,Cd胁迫下豌豆种子的萌发受到抑制,其发芽势和发芽率均明显降低,豌豆幼苗的根长、芽长、根鲜重和芽鲜重均明显降低,豌豆幼苗的Cd含量明显增加;而添加外源褪黑素能显著促进豌豆种子的萌发以及幼苗的生长,降低豌豆幼苗的Cd含量,缓解Cd胁迫对豌豆种子的毒害作用,这与蒋航等[14]研究褪黑素对重金属砷胁迫下水稻种子萌发的结果相符,说明外源褪黑素能增强胚细胞合成代谢,提高种子耐重金属胁迫的萌发能力,从而促进种子萌发、幼苗生长及根系的生理代谢。
在逆境胁迫下,植物细胞会产生大量的自由基,自由基通过启动膜脂过氧化作用使细胞膜受到损伤,导致活性氧过量积累[26-27]。活性氧过度积累容易引发细胞膜中不饱和脂肪酸发生膜脂过氧化作用,产生MDA等产物,MDA与蛋白质进行反应使其内部发生交联,从而使蛋白质变性失活,因此MDA常用来反映膜损伤程度和植物抗逆性强弱[28-29]。研究表明,植物在Cd胁迫下会导致抗氧化系统受损,植物体内活性氧清除酶(SOD、CAT和POD等)活性降低,使植物体内的活性氧、羟基自由基、单线态氧和过氧化氢等含量明显增加[30]。高浓度的活性氧会进一步破坏细胞膜的流动性和通透性,损伤脂质、核酸和蛋白质等主要的生物大分子,细胞逐渐程序性死亡,最终导致植物生长发育受抑制[31-34]。本试验结果表明,随着Cd2+处理浓度的增加,豌豆幼芽的抗氧化酶活性均逐渐降低,膜脂过氧化产物MDA含量逐渐升高,说明较高浓度的Cd2+进入植物体内,对植物产生的伤害远远超出了植物自我修复的能力范围,导致细胞受到严重的损伤。这与田丹等[35]研究发现随着Cd胁迫浓度的升高,生菜幼苗根芽中的MDA含量和POD活性逐渐提高的结果不一致,可能与Cd胁迫浓度以及不同植物材料对Cd胁迫的耐受性不同有关。本研究通过褪黑素浸种处理发现,随着MT浓度增加,豌豆幼芽的SOD、CAT和POD活性均表现出先升高后降低的趋势,MDA含量先降低后升高,说明MT作为一种抗氧化剂,直接参与了清除活性氧自由基的反应,显著提高了豌豆幼芽的SOD、CAT和POD活性,而对逆境胁迫下豌豆幼芽体内的活性氧清除能力及细胞膜保护效应会随MT的浓度升高有所减弱。这与Posmyk等[36]在研究不同浓度褪黑素前处理对紫甘蓝在铜(Cu)胁迫下种子萌发的影响时所得结论较为一致,说明褪黑素具有明显的剂量效应,对逆境胁迫下植物体的代谢影响是一个复杂的过程,与很多生理生化过程和信号传递都有关,也可能与不同植物体内褪黑素的含量有关。
4 结论
本研究结果表明,Cd胁迫严重抑制了豌豆种子的萌发以及幼苗的生长,施用外源褪黑素(MT)能有效缓解Cd胁迫对豌豆种子及幼苗的损害,且以100 μmol·L-1MT处理效果最佳。外源MT能够通过提高抗氧化酶活性,增强活性氧及自由基的清除能力,降低脂质过氧化的终产物MDA含量,减少植物Cd的积累,从而增强植株的抗逆性,促进豌豆种子萌发及幼苗的生长,减少重金属胁迫伤害。本研究结果为受重金属污染的农田作物的抗逆栽培提供了理论依据和技术支撑,但有关MT对镉胁迫下植株体内活性氧代谢的直接关系与调节机理仍有待进一步深入研究。
猜你喜欢
现代青年·精英版(2022年4期)2022-05-31
北方人(B版)(2021年11期)2021-12-28
少男少女·小作家(2021年3期)2021-08-06
现代园艺(2020年19期)2020-10-02
种子(2019年6期)2019-07-13
作文周刊·小学一年级版(2019年20期)2019-06-27
种子(2019年4期)2019-05-28
现代农村科技(2019年4期)2019-01-06
课外生活(小学1-3年级)(2017年5期)2017-06-10
故事作文·高年级(2016年9期)2016-10-25
