
硫化氢参与植物抗逆境胁迫研究进展及其在果蔬保鲜中的应用
2019-09-21 02:45:48姜爱丽刘程惠管磬馨孙小渊胡文忠
核农学报 2019年11期
陈 晨 姜爱丽 刘程惠 管磬馨 孙小渊 胡文忠
(大连民族大学生命科学学院/生物技术与资源利用教育部重点实验室,辽宁 大连 116600)
硫化氢(H2S)是继一氧化氮(NO)和一氧化碳(CO)之后在动物体内发现的第三种气体信号分子[1]。哺乳动物体内能合成H2S,其在人体内大部分组织和血清中的平均浓度为5.2×10-6mol·L-1,主要参与调节血管舒张、神经传导等多种生命活动进程[2]。植物体内也存在内源H2S。研究发现低浓度的H2S参与调控植物气孔关闭、种子萌发、根系发育等多种生理过程以及缓解干旱、重金属等逆境胁迫带来的氧化损伤[3-5]。作为小分子气体信号分子,H2S可以自由穿过细胞膜进入细胞内部,快速到达靶细胞,直接与相应的靶细胞或靶分子结合,具有痕量即可快速发挥高效生理调节功能、安全且残留量低等优点[6]。近年来,大量研究表明,外源低浓度的H2S供体NaHS(最高浓度为 1.0 mmol·L-1,释放的H2S气体浓度约为1.5×10-10mol·L-1)能够激活采后果实抗氧化系统,调节活性氧(reactive oxygen species,ROS)代谢平衡,延缓果实衰老[7-10]。H2S处理也能通过调控ROS代谢抑制鲜切果蔬褐变的发生,从而提高其保鲜效果[11-12]。本文通过对H2S参与植物抗逆境胁迫响应的研究进展,以及H2S在果蔬采后保鲜中的应用现状进行综述,旨在为H2S的深入研究提供理论依据。
1 植物体内H2S的生物合成途径及调控
H2S在植物体内合成途径如图1所示。催化H2S合成的生物酶可以分为两类:一类是半胱氨酸脱硫基酶(Cys desulfhydrases,CDes),包括L-半胱氨酸脱硫酶(L-Cys desulfhydrase,L-CD)和D-半胱氨酸脱硫酶(D-Cys desulfhydrase,D-CD)[13]。其中,L-CD以L-半胱氨酸(L-cysteine,L-Cys)为底物,催化生成H2S、氨和丙酮酸,当植物受到病原菌侵染时,L-CD活性和表达量上升,L-CD是植物抗病反应中生成H2S的主要酶[14];D-CD主要调控D-半胱氨酸(D-cysteine,D-Cys)合成H2S,活性通常较L-CD低。另一类是O-乙酰基-L-丝氨酸(硫醇)裂解酶[o-acetyl-L-serine (thiol) lyase,OAS-TL],催化无机硫生成半胱氨酸,同时也会产生少量的H2S[15]。在拟南芥中已鉴定出9个OAS-TL基因,分别位于细胞质、线粒体或质体中[16]。最近研究发现的一种新型L-半胱氨酸脱硫基酶(desulfhydrase 1,DES1),通过序列特征比对,属于OAS-TL家族[17]。此外,拟南芥中还含有固氮酶铁硫簇(nitrogenase Fe-S cluster,Nifs/NFS),具有与L-CD相似的活性,推断Nifs/NFS也可能与拟南芥合成H2S相关[18]。
2 H2S在植物抗逆境胁迫中的作用
植物的逆境胁迫包括生物胁迫和非生物胁迫,其中生物胁迫主要包括病虫害等作用,非生物胁迫包括重金属、高盐、干旱、高温等作用。研究表明,外源H2S能够增强植物对逆境胁迫的抗性。一方面H2S通过增强植物细胞的抗氧化系统或直接清除ROS,缓解或修复逆境胁迫对植物的氧化损伤;另一方面,H2S作为信号分子,与植物体内其他信号分子相互作用,共同参与抗逆信号转导和诱导抗逆基因表达[4]。
2.1 抵御重金属胁迫
重金属对植物的毒性作用表现在其可以诱导产生过量的ROS,从而引起氧化损伤,包括脂质过氧化、蛋白质氧化、酶活性丧失、DNA损伤等[19]。Chen等[20]研究表明,H2S可以直接清除过量的ROS或者通过提高花椰菜种子内部非蛋白硫醇和总谷胱甘肽含量螯合金属铅,从而减轻氧化损伤,促进铅胁迫下花椰菜种子的萌发及生长。H2S还通过激活抗氧化防御系统,提高植物抗氧化能力,进而增加其对重金属的抵抗作用。Qian等[21]和Montesinos-Pereira等[22]研究表明,H2S处理能够显著提高油菜和甘蓝幼苗的酶与非酶抗氧化能力,从而减轻氧化损伤,提高2种蔬菜在铝胁迫下的生长能力。Zhang等[23]H2S能通过调节抗氧化物酶活性,缓解Cu2+对小麦种子萌发的抑制作用。此外,H2S还能够与脯氨酸联合缓解镉(Cd)对谷子幼苗的损伤[24]。
2.2 耐盐性
H2S处理可以缓解NaCl对小麦种子萌发和幼苗生长的抑制作用[25]。研究认为H2S处理通过提高幼苗的抗氧化能力,减轻盐胁迫对其造成的氧化损伤,从而在一定程度上提高了幼苗对盐胁迫的耐受能力[26]。Shan等[27]也发现H2S通过上调玉米抗坏血酸和谷胱甘肽代谢维持抗坏血酸和谷胱甘肽的氧化还原状态,提高其抗氧化能力,进而增强了玉米耐盐性。此外,H2S还可以通过其他途径提高植物的耐盐性。如通过调节非选择性阳离子通道(nonselective cation channels,NSCCs)和盐过敏感调控(salt overly sensitive,SOS1)途径来缓解盐胁迫对小麦幼苗生长的抑制作用[28]。H2S还可以提高植物幼苗的光合效率,增强植株的抗盐性[29]。Wang等[14]采用NaHS和NO供体(硝普钠)预处理紫花苜蓿(Medicagosativa),发现其能够促进100 mmol·L-1NaCl胁迫下种子的发芽及后期幼苗的生长,提高根部的K+/Na+,进一步研究发现添加特定NO清除剂(carboxy-PTIO potassium salt,cPTIO),能够逆转上述保护作用,表明H2S的保护作用可能与诱导产生内源性NO有关。
2.3 抗旱能力
Jin等[30]通过研究拟南芥抗旱相关基因的表达调控及其与抗旱相关性,发现干旱相关基因表达模式与L-CD和D-CD基因表达模式一致,干旱胁迫能够诱导植物产生H2S。当采用NaHS处理后,幼苗的成活率显著提高,进一步分析发现,H2S能够减小植物的气孔孔径,从而减少蒸腾作用,缓解干旱胁迫。H2S还可以通过调节离子流引发气孔关闭,进而提高拟南芥抗干旱胁迫能力[31]。此外,H2S能够显著提高植物的抗氧化能力,有效缓解ROS引发的氧化损伤,从而促进干旱胁迫下水稻种子的萌发以及玉米幼苗的生长[32-33]。Li等[34]采用转录组学技术分析发现,H2S对小麦干旱损伤的缓解作用与运输系统、植物激素信号、蛋白质加工途径以及脂肪酸、氨基酸代谢有关。由此可见,H2S调控植物耐干旱胁迫是一个复杂的生理过程,受多种信号分子调节并涉及众多的代谢途径。
2.4 抵御高温胁迫
研究发现H2S处理能够提高植物的耐热性。Christou等[35]采用10 mmol·L-1NaHS水溶液预处理草莓根部48 h,显著提高了热激胁迫下草莓叶片的叶绿素荧光,气孔导度(stomatal conductance,Gs)和含水量,同时降低了细胞离子渗透率和脂质过氧化水平,从而增强了其耐热性。研究发现NaHS处理能以剂量依赖的方式提高小麦幼苗的耐热性,它是通过调节抗氧化防御系统缓解高温胁迫下小麦幼苗的抗氧化损伤,从而提高小麦幼苗对高温胁迫的耐受性[36]。Li等[37]研究认为,外源H2S一方面可以提高烟草悬浮细胞抗氧化酶活性,缓解氧化伤害;另一方面可以诱导胞外Ca2+通过Ca2+载体CaM进入细胞,传导热激信号,调控植物代谢变化,抵抗高温胁迫。研究发现NaHS预处理可以提高Δ1吡咯啉-5-羧酸合成酶活性(Δ1-pyrroline-5-carboxylate synthetase,P5CS)和降低脯氨酸脱氢酶(proline dehydrogenase,ProDH)活性,使玉米幼苗内源脯氨酸大量积累,从而显著提高热胁迫下玉米种子发芽率和幼苗存活率[38]。上述研究表明,H2S可以提高植物耐热性和获得耐热性,这可能与抗氧化系统的激活以及脯氨酸的积累有关。
2.5 增强抗病性
研究表明,高浓度H2S(1.5 mmol·L-1NaHS 浸泡处理4 h)会引起植物叶片病变,落叶,甚至引起敏感植株的死亡,而低浓度H2S(0.5 mmol·L-1NaHS)能提高植物的抗病性,并促进植物生长[39]。在受病原菌感染的植物中H2S合成关键酶LCD的活性会增加,同时释放一定量的内源性H2S[40]。上述研究表明,H2S信号可能参与植物的抗病反应。同时,H2S还具有一定的杀菌能力。Fu等[41]和Hu等[11]研究发现,外源性H2S可能通过引发真菌ROS爆发直接抑制采后水果病原菌,如黑曲霉、意大利青霉菌和扩展青霉菌的生长。
3 H2S与其他信号分子相互作用
3.1 H2S与Ca2+的相互作用
研究表明,H2S信号与钙(Ca)信号通路相互作用,调节植物的生命活动[4]。Ca2+赋予细胞壁刚性结构,并通过Ca调素调节植物生长进程。外源H2S诱导胞外Ca2+通过Ca2+载体CaM进入植物细胞内,从传导热激信号,调控植物代谢变化,抵抗高温胁迫[36]。Li等[37]研究发现NaHS预处理能提高细胞外Ca2+进入玉米悬浮培养细胞的能力,通过细胞内Ca调素介导来提高其耐热性。
3.2 H2S与H2O2的相互作用
非生物胁迫能够诱导植物合成H2S和H2O2,但目前对二者在应答逆境胁迫过程中如何相互作用仍存在争议。王文杰等[42]研究发现受霜霉病菌侵染后,葡萄叶片H2O2含量猝发早于H2S,且H2O2清除剂抗坏血酸可显著抑制霜霉病菌诱导的H2S含量上升,而H2S清除剂次牛磺酸(hypotaurine,HT)对霜霉病菌诱导的H2O2含量变化影响不显著,推测H2S作用于H2O2的下游参与葡萄抵御霜霉病过程。Hou等[43]也认为在茉莉酸(jasmonic acid,JA)诱导的蚕豆气孔运动中H2S存于H2O2信号分子的下游。Liu等[44]对拟南芥的研究也发现了类似的结果。然而,也有一些学者持相反观点,Wang等[45]研究发现H2S可通过胞外ATP(eATP)调控H2O2,进而调节拟南芥气孔保卫细胞钾离子通道诱导气孔关闭。H2O2能够激活质膜钙通道,H2S通过提高胡杨细胞抗氧化水平降低H2O2含量,进而调节质膜钙通道抑制镉进入细胞内,缓解镉胁迫损伤[46]。上述结果表明,H2S作用于H2O2的上游参与植物抗逆境胁迫。
3.3 H2S与NO的相互作用
NO是重要的气体信号分子之一,在植物生长和发育中发挥着重要作用,同时还可作为植物生长调节物质调控植物的成熟和衰老[47]。研究发现镉胁迫能够诱导狗牙根产生内源性NO和H2S,NO和H2S处理通过提高狗牙根抗氧化能力缓解镉胁迫对植物生长的抑制以及对细胞的损伤,且二者均能够提高内源性NO和H2S的含量。进一步研究发现NO和H2S清除剂均能够抑制镉胁迫下NO信号的产生,但NO清除剂并不能阻断H2S信号,只有H2S清除剂能够阻断H2S信号的产生,由此推测H2S信号调节了NO介导的抗镉胁迫反应[48]。研究表明,NO通过诱导内源性H2S的积累提高植物的抗逆境胁迫[49-50],H2S可能作为NO下游的信号分子参与植物的抗逆境胁迫反应[26,51]。同时,也有研究认为NO作用于H2S下游参与植物的逆境胁迫[52-53]。Amooaghaie等[54]研究发现H2S和NO均能够提高芝麻幼根对铅的耐受能力,NaHS处理能够提高内源性NO的含量,NO清除剂处理能够阻断H2S介导的芝麻幼根对铅胁迫的耐受响应,而采用H2S清除剂处理并未影响NO介导的抗逆境反应。上述研究表明,H2S与NO存在着复杂的相互作用,推测H2S处理提高了植物体内NO含量,NO又反馈调节内源性H2S的合成,二者联合共同激活植物防御系统,增强植物的抗逆境胁迫。
3.4 H2S与植物激素的相互作用
H2S和植物激素在植物抗逆境胁迫反应中有着复杂的相互作用。干旱胁迫下,植物体内CDes基因表达上调,同时H2S含量上升[30]。进一步研究H2S和脱落酸(abscisic acid,ABA)的相互作用,发现在H2S缺陷的拟南芥lcd突变株中,ABA诱导气孔关闭的作用会减弱,表明H2S在ABA诱导植物气孔关闭中发挥重要作用。H2S参与离子通道基因的表达调控,也影响ABA受体的表达,因此,H2S可能在ABA信号通路上游起调控作用[55]。水杨酸(salicylic acid,SA)预处理可以提高玉米幼苗的耐热性,SA诱导的耐热性通过添加NaHS而增强,当添加特异性的H2S生物合成抑制剂DL-炔丙基甘氨酸和H2S清除剂后减弱,当采用SA生物合成抑制剂多效唑和氨基茚磷酸预处理玉米幼苗后,并没有显著影响NaHS诱导的耐热性,且NaHS处理后在玉米幼苗中未检测到内源SA,因此,推测H2S可能在SA信号通路下游参与玉米幼苗的耐热反应[56]。此外,Shan等[57]研究发现H2S也参与了茉莉酸(JA)对拟南芥抗坏血酸-谷胱甘肽代谢的调节作用。外源H2S处理的番薯、甘薯、大豆根均会诱导吲哚乙酸(indole acetic acid,IAA)产生[58]。
4 H2S对采后果蔬的保鲜效果
4.1 采后果蔬
采后果蔬在贮藏过程中衰老进程加快,易遭受病原微生物的侵染,导致其腐烂变质,保鲜期缩短,造成较大的经济损失。近年来,许多学者研究发现H2S处理可以有效减缓贮藏期间果蔬衰老及腐烂等问题,H2S在采后果蔬的应用情况如表1所示。H2S熏蒸处理能够显著抑制采后草莓的腐烂,并提高其硬度,降低呼吸强度,进一步分析发现H2S处理显著提高了草莓抗氧化酶[过氧化氢酶(catalase,CAT)、过氧化物酶(peroxidase,POD)、抗坏血酸过氧化物酶(ascorbate peroxidase,APX)和谷胱甘肽还原酶(glutathione reductase,GR)]活性,抑制脂氧合酶(lipoxidase,LOX)活性,同时减少了H2O2和MDA含量的积累。由此可见,H2S通过提高采后草莓的抗氧化能力,减轻氧化损伤,从而延缓其衰老,延长了贮藏期[7]。Shen等[10]和Hu等[59]研究也表明,H2S可以通过激活低温贮藏期间采后猕猴桃和桑葚的抗氧化系统,从而保持其贮藏期间的品质,延缓其成熟和衰老。H2S还能够通过抑制乙烯的释放,降低绿叶蔬菜在贮藏期间叶绿素的降解,从而延缓其衰老进程[60-61]。此外,H2S通过增加脯氨酸含量和提高抗氧化酶活性,参与能量代谢以维持能量电荷来降低采后香蕉果实冷害[62-63]。但H2S对采后果蔬衰老的调控具有双重生物效应,低浓度的H2S能较好保持采后果蔬贮藏期间的品质,而高浓度H2S会加速其成熟和衰老,不利于采后果蔬的保鲜。汪伟等[9]研究发现低浓度H2S(15 μmol·L-1)通过提高采后桃果实活性氧代谢来延缓果实衰老,但高浓度H2S(30、45 μmol ·L-1)会加速采后桃果实的衰老。
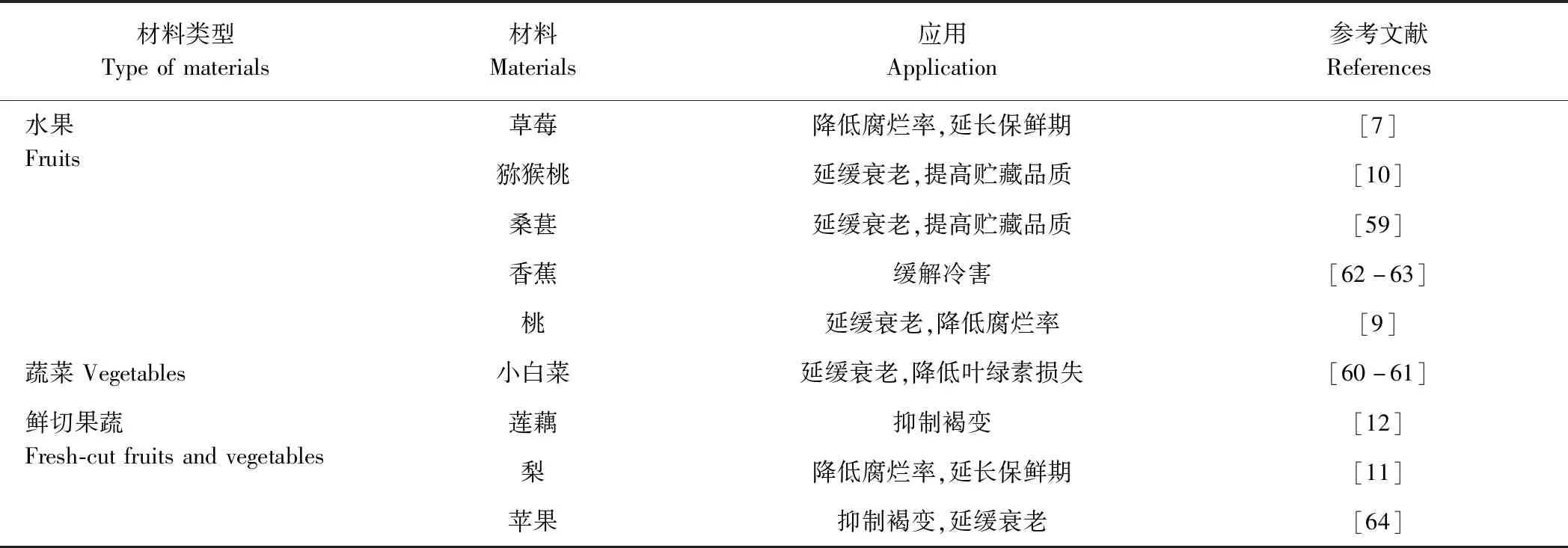
表1 硫化氢在采后果蔬中的应用Table 1 Application of hydrogen sulfide in the preservation of postharvest fruits and vegetables
4.2 鲜切果蔬
新鲜果蔬在去皮、切分等加工过程中,细胞组织结构遭到伤害,呼吸速率升高,营养物质流失,极易受到微生物侵染以及产生褐变等,从而加速果蔬的衰老和腐败。H2S作为一种抗氧化剂可以直接清除切割伤害引发的ROS,同时H2S还可以激活果蔬自身的抗氧化防御系统,及时清除ROS,减缓氧化损伤。此外,H2S可以抑制鲜切莲藕的多酚氧化酶(polyphenol oxidase,PPO)和POD活性,从而有效抑制其在贮藏过程中的褐变反应,获得较好的保鲜效果[12]。NaHS熏蒸处理也能够上调鲜切梨APX、CAT、POD等抗氧化酶活性,下调LOX、苯丙氨酸解氨酶(phenylalnine ammonialyase,PAL)和PPO活性,降低ROS及MDA含量的积累,同时,NaHS熏蒸还可以有效抑制鲜切梨的病原真菌(黑曲霉和扩展青霉菌)的生长,表明H2S可作为抗氧化剂和杀菌剂来延长鲜切梨的贮藏保鲜[11]。Zheng等[64]研究发现H2S通过下调衰老相关基因(MdLOX2、MdPG1、MdPPO、MdACO1、MdERS1和MdETR1)的表达同时提高其抗氧化能力,进而延缓了鲜切苹果的衰老。
5 展望
低浓度的H2S在植物生长发育以及抗逆境胁迫响应中发挥着重要的调节作用,但H2S在植物整个生命周期的关键作用及其抗逆境胁迫反应机制尚不清楚。植物细胞如何感知H2S信号,以及在H2S参与植物抗逆境胁迫反应过程中,中间产物及其他物质如何影响H2S信号转导、基因表达等方面都有待进一步研究。在植物抗逆境胁迫响应中,H2S与其他信号分子(如H2O2、NO、植物激素等)有着密切的联系,但这些信号分子之间的相互作用机制,以及它们各自在发挥作用时是否受相互之间的调控(或引导或抑制),也将是未来的研究重点。近年来,H2S在采后果蔬保鲜中的应用越来越多,但H2S延缓果蔬产品成熟衰老和抗病的作用尚处于生理阶段的研究,还需进一步深入明确H2S如何调控果蔬采后代谢、延缓组织成熟和衰老的分子机理,为探索H2S在果蔬采后贮藏保鲜的广泛应用提供理论依据。
猜你喜欢
做人与处世(2022年6期)2022-05-26 10:26:35
顺德职业技术学院学报(2022年2期)2022-05-24 03:32:58
疯狂英语·新阅版(2022年5期)2022-04-29 05:07:07
杭州化工(2020年2期)2020-01-16 15:26:40
童话世界(2019年26期)2019-09-24 10:57:56
启蒙(3-7岁)(2018年8期)2018-08-13 09:31:14
基层中医药(2018年2期)2018-05-31 08:45:16
草业科学(2016年6期)2016-07-16 01:29:16
童话世界(2016年8期)2016-06-02 09:21:05
安徽建筑(2014年6期)2014-08-15 00:50:38
