
羽毛球专项运动对踝关节肌肉共收缩活动的影响研究
2017-08-01 00:19王乐军陈景源马爱迪游松辉牛文鑫龚铭新
中国体育科技 2017年4期
王乐军,陈景源,马爱迪,游松辉,李 岩,牛文鑫,龚铭新
WANG Le-jun1,CHEN Jing-yuan1,MA Ai-di1,YOU Song-hui1,LI Yan2,NIU Wen-xin3,GONG Ming-xin1
羽毛球专项运动对踝关节肌肉共收缩活动的影响研究
王乐军1,陈景源1,马爱迪1,游松辉1,李 岩2,牛文鑫3,龚铭新1
WANG Le-jun1,CHEN Jing-yuan1,MA Ai-di1,YOU Song-hui1,LI Yan2,NIU Wen-xin3,GONG Ming-xin1
目的:对比高水平羽毛球专项运动员与普通大学生在等速屈伸踝关节运动中的拮抗肌共收缩水平,探讨羽毛球专项运动对屈伸踝关节运动中拮抗肌共收缩水平的影响作用,从神经生理学角度揭示羽毛球专项运动与踝关节神经肌肉控制的内在联系。方法:受试者以8名高校高水平羽毛球运动员作为实验组,10名普通大学生为对照组。分别进行屈伸踝关节静态最大自主收缩力测试,并以30°/s、60°/s、120°/s、180°/s的角速度进行踝关节屈伸运动。记录实验过程中受试者屈伸踝关节峰值力矩和受试者胫骨前肌、外侧腓肠肌表面肌电信号(sEMG)。基于sEMG计算踝关节背屈和跖屈运动中的拮抗肌共收缩水平。采用重复测量方差分析对实验组与对照组测试参数的差异性进行检验(主体内因素:关节收缩角速度;主体间效应:实验组-对照组)。结果:实验组在4种角速度下的屈、伸踝关节峰值力矩都显著高于对照组,其中背屈峰值力矩的差别尤为明显。实验组受试者踝关节背屈时的拮抗肌共收缩水平要显著低于对照组,而跖屈时的拮抗肌共收缩水平则无显著性差异。结论:长期的羽毛球专项运动引起踝关节背屈运动的拮抗肌共收缩水平降低,关节净力矩及肌肉收缩效率提高。这些羽毛球运动引起的拮抗肌共收缩活动改变,是在专项运动对人体运动能力的特定需要下,运动皮层及脊髓等中枢神经系统功能活动产生适应性改变,并在此基础上形成拮抗肌活动控制的特定固化模式而产生的。
羽毛球运动;踝关节;拮抗肌共收缩;表面肌电信号
1 前言
在人体运动中,中枢神经系统对主动肌与拮抗肌的协同控制是实现肢体协调运动的基本条件[3,24]。一方面,中枢神经系统通过“交互抑制”弱化拮抗肌的活动,以保证关节收缩效率和产生适宜的关节净力矩,进而完成特定的运动任务[14,18,20]。另一方面,在主动肌收缩的同时,拮抗肌在高位中枢的调控下维持一定的收缩活动,以保障运动中关节的稳定性和合理分配关节周围组织载荷等[19,26,35]。交互抑制与共激活共同作用于拮抗肌的活动控制过程,并构成关节收缩效率与关节稳定性相互联系和相互制约的对立统一关系。
运动训练可以显著提高肌肉收缩力量和关节净力矩水平。前期研究表明,除了肌肉本身围度增加的因素,运动训练所引起的神经肌肉控制机制适应性变化亦在肌肉力量增长中发挥重要的作用[2]。在诸多引起肌肉力量增加的神经控制机制中,拮抗肌共收缩活动水平的降低被认为是引起关节净力矩增加的重要潜在原因。例如,Bazzucchi等[10]的研究发现,高水平网球运动员在等速屈肘运动中的拮抗肌(肱三头肌)共收缩水平要显著低于普通受试者;对膝关节伸肌的等动收缩测试结果显示,优秀运动员的半腱肌拮抗力明显低于经常静坐的受试者[2,8]。但也有研究发现,在运动训练引起肌肉力量增加的同时,拮抗肌的拮抗作用在不同年龄和性别组内并未表现出一致性的减小[2,25]。因此,对拮抗肌共收缩活动在肌肉力量训练中所起作用的认识,仍需进一步的研究与探讨。
羽毛球是一项剧烈的运动,在羽毛球运动过程中,运动员需要完成众多快速启动、疾跑急停和快速跳跃等动作,对运动员下肢各关节的运动能力都有非常高的要求。此外,长期的羽毛球专项运动也在促使下肢神经肌肉活动得以训练和改进。前期研究发现,羽毛球运动员下肢运动能力表现出较强的专项特征,并在下肢运动能力、下肢肌肉收缩效率等方面显著优于普通受试者[9,37]。但是,对于这些改变背后的潜在机制却鲜有研究报道。因此,本研究通过对比分析高水平羽毛球专项运动员与普通大学生等速屈伸踝关节运动中的拮抗肌共收缩水平,探讨羽毛球专项运动对屈伸踝关节运动中拮抗肌共收缩水平及神经肌肉控制机制的影响作用。
2 研究对象、方法与步骤
2.1 研究对象
本实验受试者皆为男性,随机选取8名高校高水平羽毛球运动员作为实验组,上海市高校普通在读本科生和研究生10名作为对照组(未进行过任何系统的专业运动训练)。实验组都接受过5年以上的羽毛球专项运动,且皆为国家一级以上运动员,平时每周至少进行5次、每次2 h的羽毛球专项运动。所有受试者在实验前需接受问卷调查,确保受试者无踝关节神经肌肉系统疾病和重大疾病史,身体健康状况良好。在实验前告知受试者相关的测试方法、程序及注意事项,并签订研究内容知情同意书。受试者基本情况如表1所示。
2.2 实验程序
受试者在测试前进行充分的准备活动后,仰卧在Contrex肌力测试系统(瑞士CONTREX公司,型号PM1/ MK2a,b)的测试床上,采用绑带将上身与测试床缚在一起以限制躯干的运动。两手交叉置于胸前。右足放置于踏板上并固定。调节杠杆臂的位置,使受试者髋关节固定于145°,右膝固定于120°,踝关节处于中立位[29]。受试者保持此姿势,对受试者进行3次踝关节屈、伸肌静态自主最大收缩力(MVC)测试,每次测试时间20 s,各次测试的时间间隔为5 min,取最大值作为该受试者最大屈、伸踝关节的MVC。
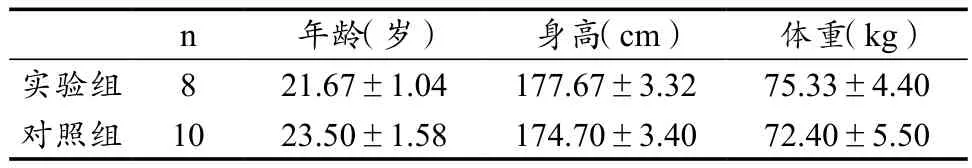
表1 受试者基本情况Table 1 Basic Information of Subjects
受试者进行充分的休息后,继续保持上述MVC测试姿势,分别按以下顺序进行4组恒定角速度的测试:第1组角速度:30°/s,向心性踝关节屈伸运动×3次;第2组角速度:60°/s,向心性踝关节屈伸运动×3次;第3组角速度:120°/s,向心性踝关节屈伸运动×3次;第4组角速度:180°/s,向心性踝关节屈伸运动×3次。前后两组测试之间休息3min。采用伸肌向心/屈肌向心测试模式。测试时踝关节角度的运动范围为-10°~ 30°[5]。记录实验过程中受试者屈伸踝关节峰值力矩等参数。
采用Delsys无线EMG采集系统(DS-B04,Bagnoli™-16 Desktop EMG system,DelsysInc.,Boston,MA)记录实验过程中受试者右侧胫骨前肌和外侧腓肠肌表面肌电信号(sEMG)。放置电极前首先进行去毛处理,并用磨砂膏打磨皮肤表面以去除角质和表面污垢,之后用75%酒精棉球清理皮肤表面,以减小阻抗。采用双面胶将测试电极固定于测试肌肉皮肤表面。电极位置参照Seniam(http:// seniam.org)所建议的测试标准进行贴放,信号采样频率为2 000 Hz。
2.3 数据的处理与分析
1.Tongji University,Shanghai 200092,China;2. Shanghai University of Sport,Shanghai 200438,China;3.Tongji University,Shanghai 200092,China.
实验测试完成后,将Contrex测试数据导出,获取屈、伸踝关节峰值力矩参数。sEMG数据从Delsys EMG works Analysis 4.2.0.0软件系统导出后,通过MATLAB 2009 Ra软件编程实现sEMG数据的处理与分析工作,其主要分为以下步骤:
1.sEMG滤波
采用4阶Butterworth滤波器对采集的sEMG进行带通滤波处理,滤波频率5-500Hz,滤波模式为零相位偏移滤波,滤波强度24 dB/oct。
2.sEMG全波整流和标准化处理
对MVC测试和4种关节角速度等速收缩过程中记录的sEMG信号分别进行全波整流和平均化处理,平均化处理的时间宽度设置为0.01 s。
3.sEMG分段
对于某受试者以特定关节角速度进行的3次踝关节等速收缩,根据Contrex测试数据,选择最大峰值力矩所对应的sEMG数据为分析数据源。对该次收缩数据,以主动肌sEMG为参照,确定主动肌激活时间阶段。肌肉激活时间标准为:sEMG的幅值高于肌肉安静时肌电水平标准差的3倍且持续50 ms时间以上[4,12]。以该时间阶段内的主动肌与拮抗肌sEMG为后期分析数据源。对于MVC测试数据,选择峰值力矩出现前后的1.024 s sEMG数据作为计算EMGMVC指标的数据源。
4.AEMG计算
设EMGt为所测定的某肌肉在时间t时刻肌电信号值(已进行全波整流和平均化处理),则该肌肉在运动时间[t1,t2]阶段内的平均肌电振幅AEMG为:
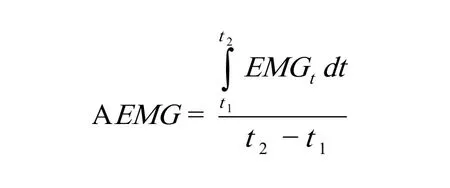
5.AEMG指标的标准化处理
对于在角速度J(J=30º/s、60º/s、120º/s、180º/s)屈伸踝关节运动过程中记录sEMG的AEMGJ指标,其标准化公式如下:
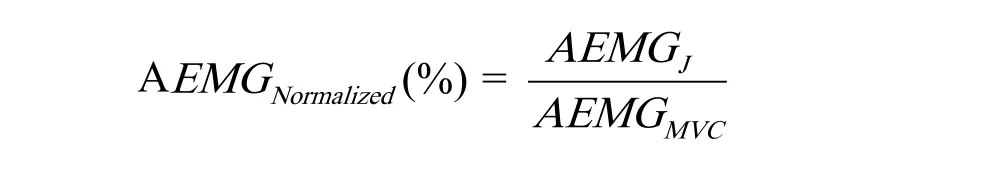
其中,AEMGJ为某肌肉sEMG的AEMG指标在标准化之前的值,为标准化后的值,AEMGMVC为该肌肉在进行MVC测试时记录的sEMG平均肌电幅值。
2.4 数据的统计分析
选用KS检验(Kolmogorov–Smirnov test)对各分析数据进行正态性检验。实验组与对照组静态屈、伸踝关节自主最大收缩力的差异性采用独立样本t检验进行检验。实验组与对照组不同角速度等速屈、伸踝关节运动中关节最大输出力矩和主动肌、拮抗肌收缩水平的差异性采用重复测量的方差分析进行检验(主体内因素:关节收缩角速度;主体间效应:实验组-对照组)。差异的显著性水平为P<0.05。数据的统计分析工作在IBM SPSS Statistics 13.0软件环境下完成。
3 研究结果
3.1 踝关节静态屈、伸自主最大收缩力
静态屈、伸踝关节自主最大收缩力矩如图1所示。通过图1可以看出,实验组踝关节背屈和跖屈静态自主最大收缩力矩分别为57.80±9.00 N·m和60.37±12.21 N·m,对照组分别为44.85±6.88 N·m和50.43±15.41 N·m。实验组踝关节背屈和跖屈静态自主最大收缩力显著高于对照组(背屈肌:P=0.005;跖屈肌:P=0.045)。
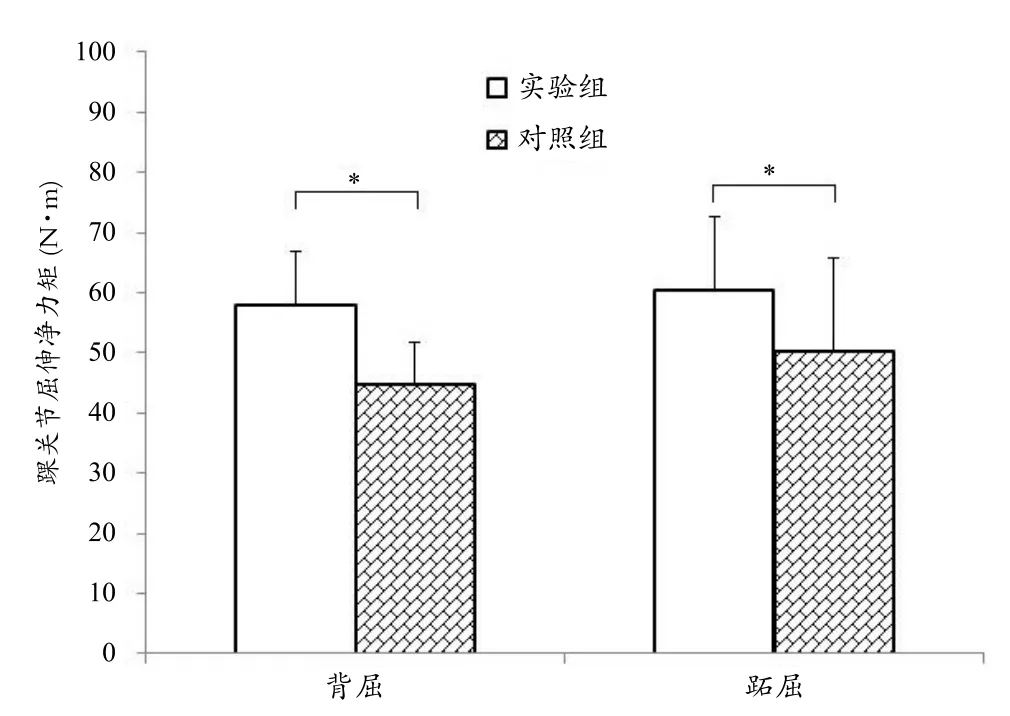
图1 实验组和对照组踝关节静态屈、伸自主最大收缩力矩比较图Figure 1 Comparison of Ankle Flexor and Extensor MVC between Non-players and Badminton Players
3.2 踝关节峰值力矩-关节角速度关系
实验组与对照组受试者在4种角速度等速屈、伸踝关节运动过程中的关节最大输出力矩如图2所示。在踝关节进行背屈运动时,随着关节运动角速度的增加,实验组踝关节最大背屈力矩由30º/s时的44.33±5.82 N·m逐渐降低至180º/s时的23.20±2.98 N·m,对照组则由34.51±8.59 N·m逐渐降低至15.69±5.43 N·m。在踝关节进行跖屈运动时,随着关节运动角速度的增加,实验组踝关节最大背屈力矩由30º/s时的46.48±9.41 N·m逐渐降低至180º/s时的24.15 ±4.89 N·m,对照组则由41.42±7.33 N·m逐渐降低至18.83±3.33 N·m。从均值看,实验组在各关节角速度的最大屈、伸踝关节力矩都要显著高于对照组,其中背屈峰值力矩的差别尤为明显。重复测量方差分析结果显示,主体内效应(关节角速度)对踝关节最大屈、伸力矩具有显著性的影响(背屈:F=16.241,P=0.000;跖屈:F=613.829,P=0.000),主体间效应(实验组-对照组)对实验结果亦有显著性的影响(背屈:F=8.599,P=0.010;跖屈:F=4.743,P=0.045)。
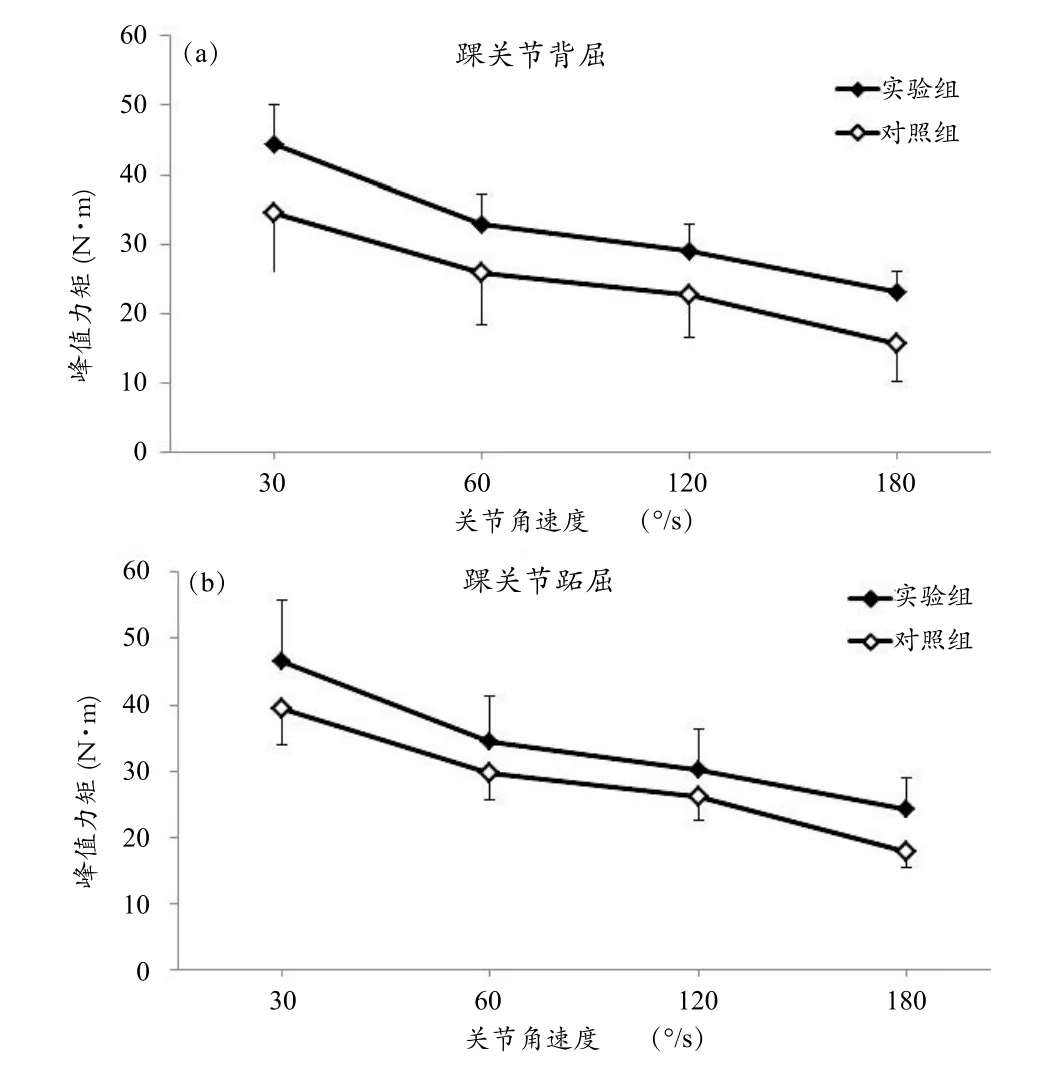
图2 不同角速度等速屈、伸踝关节运动中关节最大输出力矩-关节角速度关系曲线Figure 2 Torque-angular Velocity Relationships of Ankle Flexors and Ankle Extensors for Non-players and Badminton Players
3.3 屈、伸踝关节运动中主动肌的放电幅值
实验组与对照组在不同角速度等速屈、伸踝关节运动中的主动肌放电幅值比较如图3所示。重复测量方差分析结果显示,主体内效应1(关节收缩速度)和主体内效应2(肌肉)对踝关节等速跖屈中的外侧腓肠肌和背屈中的胫骨前肌sEMG的AEMG指标无显著性的影响(关节角速度:F=1.343,P=0.300;肌肉:F=1.048,P=0.321),主体间效应(实验组-对照组)对实验结果亦无显著性的影响(F=0.377,P=0.548)。此外,主体内因素和主体间因素之间无显著性的交互效应(收缩速度-分组:F=0.426,P=0.735;肌肉-分组:F=0.097,P=0.759)。
3.4 屈、伸踝关节运动中拮抗肌的共收缩水平
实验组与对照组在不同关节角速度等速屈、伸踝关节运动中的拮抗肌共收缩水平比较如图4所示。重复测量方差分析结果显示,主体内效应1(关节收缩速度)对踝关节等速跖屈中的胫骨前肌和背屈中的外侧腓肠肌sEMG的AEMG指标无显著性的影响(F=1.286,P=0.318),主体内效应2(肌肉)对测试指标具有显著性的影响(F=43.859,P=0.000),主体间效应(实验组-对照组)对实验结果亦有显著性的影响(F=13.322,P=0.002)。收缩速度-分组之间无显著性交互作用(F=1.340,P=0.272),而肌肉-分组之间具有显著性的交互效应(F=0.878,P=0.006)。多重比较分析结果显示,在4种等速收缩速度下,实验组与对照组腓肠肌共收缩水平具有显著性的差异,实验组受试者外侧腓肠肌共收缩水平都要显著低于对照组,而实验组与对照组的胫骨前肌共收缩水平无显著性差异。此外,实验组不同肌肉共收缩水平亦无显著性差异,但对照组在各关节角速度下的胫骨前肌和腓肠肌外侧共收缩水平皆有显著性的差别。
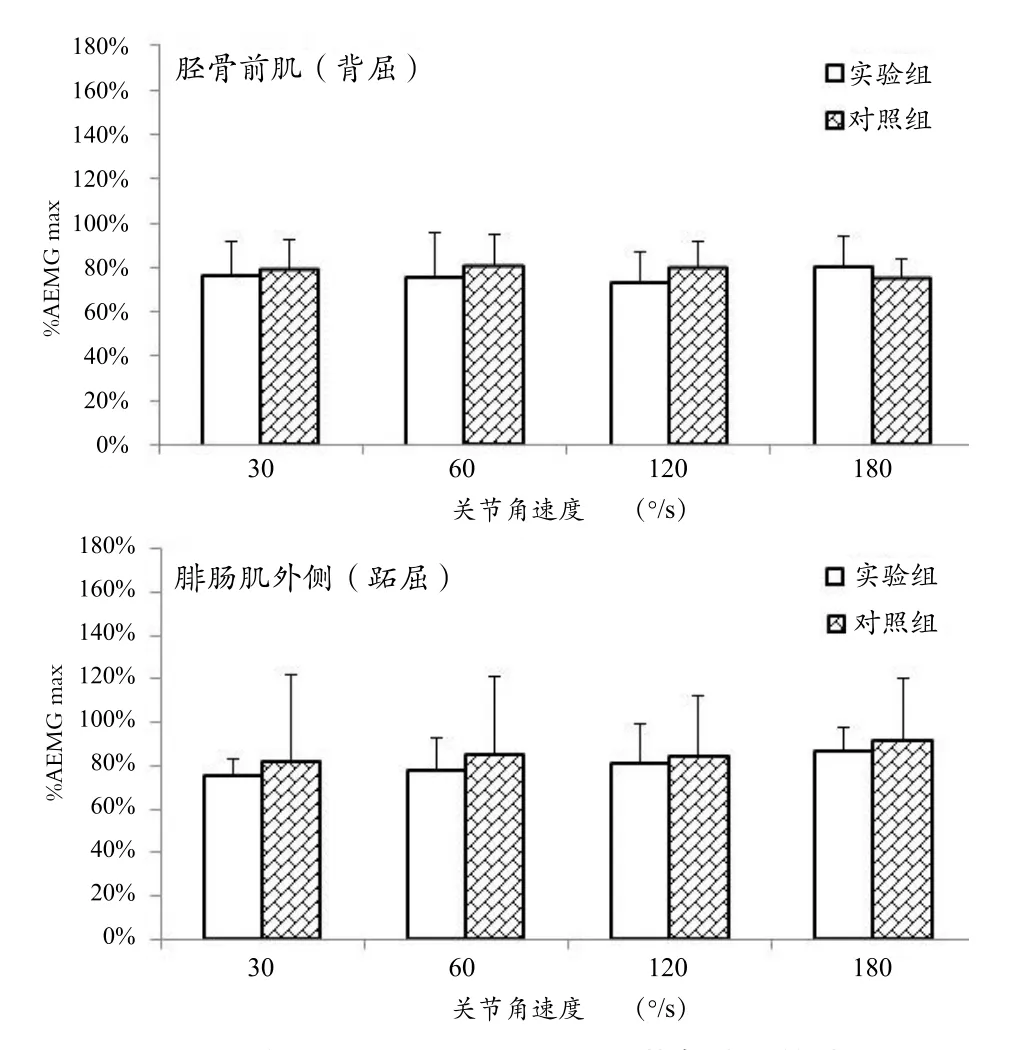
图3 实验组与对照组在不同关节角速度等速屈、伸踝关节运动中的主动肌放电幅值比较Figure 3 Agonist EMG Activities for Tibialis Anterior and Lateral Grastrocnemius in Non-players and Badminton Players
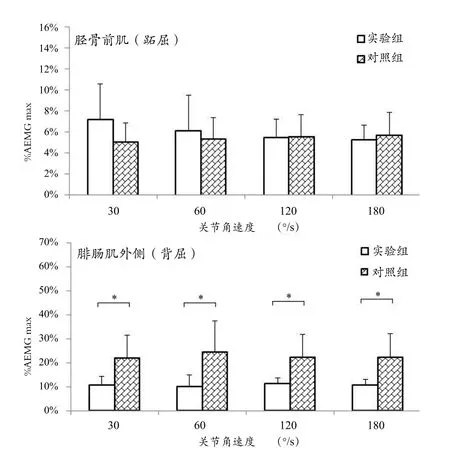
图4 实验组与对照组在不同关节角速度等速屈、伸踝关节运动中的拮抗肌共收缩水平比较Figure 4 Antagonist EMG Activities for Tibialis Anterior and Lateral Grastrocnemius in Non-players and Badminton Players
4 讨论
在拮抗肌共收缩活动的sEMG信号记录中,主动肌sEMG信号所造成的串扰作用会在一定程度上降低信号的可信度。因此,如何减小信号串扰作用对相关测试结果的影响已引起前期研究的广泛关注[21,22,30]。在本研究中,首先,Delsys表面肌电测试电极采用较小的接触面积和电极间距,可以有效降低串扰的影响作用[17,27,35];其次,Delsys测试电极选用双电极差分技术,具有良好的降低串扰的效果[17,21,23]。此外,在拮抗肌共收缩水平的指标计算上,使用基于最大自主收缩sEMG指标标准化的方法,可以有效降低受试者、肌肉、电极位置等因素的差异性对测试结果的影响作用。上述研究方法保证了本研究测试方案的科学性和实验结果的可信度。
实验组踝关节屈、伸静态自主最大收缩力和在4种角速度下的屈、伸踝关节峰值力矩都显著高于对照组,其中实验组与对照组最大背屈力矩的差值比跖屈要更加明显。这些研究结果提示,羽毛球专项运动对静态自主最大收缩和4种角速度等速收缩中的踝关节最大背屈和跖屈关节力矩都有显著性的提高作用,其中对踝关节背屈力矩的提高作用较跖屈力矩要更加明显。与之相对应,实验组受试者踝关节背屈时的拮抗肌共收缩水平要显著低于对照组,而跖屈时的拮抗肌共收缩水平则无显著性的差异。此外,4种角速度情况下,实验组与对照组屈、伸踝关节肌肉放电幅值亦无显著性差别。这些研究结果从一定程度上暗示,运动训练引起拮抗肌共收缩水平的变化与关节输出力矩之间存在着一定的联系。
在完成特定运动任务的过程中,未经过特定训练的受试者,其中枢神经系统并不能支配肌肉以最高效的方式参与收缩活动[13]。运动技能学习过程中,对拮抗肌共收缩活动的调控是中枢神经系统协调运动的重要方式,并在此基础上形成运动技能控制的固化模式[24,34]。运动训练是运动技能学习的重要方式,长期的运动训练可以促使中枢神经系统对肌肉活动的控制方式发生改变,并伴随拮抗肌共收缩活动及关节运动效率的变化。Bazzucchi等[10]的研究显示,高水平网球运动员在不同速度等速屈肘运动中的拮抗肌(肱三头肌)共收缩水平较普通受试者要低10%~20%RMS max(RMS max为肱三头肌以15º/s的角速度最大离心收缩计算的肌电均方根振幅);对不同速度的向心和离心等动伸膝运动测试结果显示,优秀跳高运动员的拮抗肌半腱肌肌电RMS较经常静坐的受试者要低24%左右[2,8]。Dal Maso等[15]的研究发现,经常参加力量锻炼的受试者,其在不同力量静态屈膝过程中的拮抗肌(股四头肌)共收缩水平较耐力性活动组受试者要低1%~2%MVC线性包络值。在本研究中,实验组以不同角速度进行踝关节背屈时的拮抗肌共收缩水平较对照组要低10%~15%AEMG max。在这些研究中,拮抗肌共收缩水平的下降,无疑会减小拮抗力矩进而增加关节净力矩和关节收缩效率。但由于目前对关节周围肌肉,特别是深层肌肉对关节力矩贡献率仍认识不足。此外,反映拮抗肌共收缩水平的肌电指标与肌肉收缩力之间也存在着非线性的复杂关系,因此目前仍然很难量化因拮抗肌共收缩水平下降引起肌力增加的贡献率。
然而,前期也有研究未发现专项运动水平与拮抗肌共收缩活动之间存在相关性。例如,Zehr等[38]的研究发现,高水平空手道运动员与普通受试者在快速伸肘运动中的拮抗肌共收缩水平并没有显著性的差别。Häkkinen等[25]的研究也发现,经过6个月的力量训练后,在膝关节伸肌显著增加的同时,拮抗肌股二头肌的共收缩水平在男、女老年组都出现显著性的下降,而在男、女中年组则无显著性的改变。因此,运动训练对拮抗肌共收缩水平的影响作用,与受试者的年龄、专项运动的方式、运动训练的时间、受试者自身的运动水平、测试的关节肌肉等都有一定的联系。
羽毛球是一项技能主导类隔网对抗性项目,对运动员的技术、速度、耐力等能力都有很高的要求。前期研究发现,在羽毛球比赛中最常用的蹬跨步、垫步等步法中,胫骨前肌不仅是踝关节中最先被募集的肌肉,而且在整个步法运动过程中是激活时间最长、放电幅值最高的肌肉[1],而在跑步运动中踝关节背屈肌参与了70%的时间周期过程[7]。因此,在羽毛球运动中,踝关节背屈肌是参与程度很高并在专项运动中不断得到训练的肌肉。在本研究中,实验组踝关节背屈运动中的拮抗肌共收缩水平明显低于对照组,表明长期的羽毛球专项运动,可以诱发中枢神经系统对踝关节背屈运动中的神经肌肉控制方式发生改变,在保证关节稳定性的同时降低拮抗肌共收缩水平和提高关节运动效率。
从跖屈踝关节运动中的拮抗肌共收缩水平看,实验组与对照组踝关节跖屈运动中的拮抗肌共收缩水平均值都在8%以下,要低于背屈踝关节运动中的拮抗肌共收缩水平。前期研究也发现,等速伸膝运动中拮抗肌(腘绳肌)的共收缩比率要高于屈膝时的拮抗肌(股四头肌)共收缩比率[6]。对等速屈伸肘关节运动中拮抗肌共收缩比率的比较研究结果发现,等速屈肘时的拮抗肌共收缩水平要高于伸肘[11]。这种同一关节不同维度运动中拮抗肌共收缩水平的差异性,可能与肌群力量配布、日常生活中的体育锻炼等因素有关[11]。步行是日常生活中最为常见的运动方式之一。在步行运动中,小腿肌在支撑期时相主动向心收缩,是步行中人体向前推进的主要动力来源之一,而胫骨前肌则主要在支撑前期中的触地缓冲和摆动时相中的足部姿势控制等方面发挥作用,其用力大小和激活时间都要小于步态中踝关节跖屈肌的活动[33]。因此,长期的步行运动对踝关节跖屈肌的锻炼效果是非常显著的。在此基础上,中枢神经系统已形成对踝关节跖屈运动控制的固化模式,这或许是造成普通人群踝关节跖屈时拮抗肌共收缩水平低于背屈时拮抗肌共收缩水平的原因。
在拮抗肌共收缩活动的调控机制上,De Luca C J与Erim Z[16]提出,中枢神经系统对主动肌与拮抗肌采取“共驱动”的控制策略。之前大量研究也认为,疲劳后主动肌与拮抗肌肌电活动的同步增加,正是中枢神经系统对主动肌与拮抗肌“共驱动”指令增加造成的[19,31]。此外,Levenez M等[28,29]基于电刺激神经肌肉通路实验的研究表明,共驱动并不是拮抗肌共收缩活动的唯一调控方式,其中,伦肖细胞的活动可以改变Ia型中间抑制性神经元和来自高尔基腱器官的Ib型传入神经元的兴奋性,被认为与拮抗肌共收缩活动的调节有一定的关系[10,36]。前期研究发现,运动训练可以改变脊髓反射的兴奋性,并改变肌张力反射性调节过程[8,32]。Dal Maso等的研究发现,力量训练引起拮抗肌共收缩水平的降低,伴随特定区域运动皮层激活水平的改变,提示力量训练引起的拮抗肌共收缩水平变化与训练引起运动皮层活动的适应性改变有关[15]。这些研究为认识拮抗肌共收缩活动的中枢调控机制提供了新的思路与视角,并为运动训练引起拮抗肌共收缩活动的改变提供了潜在的生理学解释。
因此,长期的羽毛球专项运动引起踝关节背屈运动的拮抗肌共收缩水平降低和关节净力矩及肌肉收缩效率提高。这些羽毛球运动引起的拮抗肌共收缩活动改变,是在专项运动对人体运动能力的特定需要下,运动皮层及脊髓等中枢神经系统功能活动产生的适应性改变,并在此基础上形成拮抗肌活动控制的特定模式而产生的。实验组与对照组踝关节背屈运动中的拮抗肌共收缩水平无显著性差异,可能与日常步行等活动已促使中枢神经系统形成对踝关节跖屈运动控制的固化模式有关。
5 结论
长期的羽毛球专项运动引起踝关节背屈运动的拮抗肌共收缩水平降低和关节净力矩及肌肉收缩效率提高。这些羽毛球运动引起的拮抗肌共收缩活动改变,是在专项运动对人体运动能力的特定需要下,运动皮层及脊髓等中枢神经系统功能活动产生的适应性改变,并在此基础上形成拮抗肌活动控制的特定固化模式而产生的。
[1] 刘宝林. 羽毛球运动员前场上网步法和专项力量训练动作肌肉用力特征的肌电分析[D]. 西安:西安体育学院,2012.
[2] 美扎齐奥尔斯基. 运动生物力学:运动成绩的提高与运动损伤的预防[M].北京: 人民体育出版社,2004.
[3] 王健,杨红春,刘加海. 疲劳相关表面肌电信号特征的非疲劳特异性研究[J]. 航天医学与医学工程. 2004,(1):39-43.
[4] 王乐军,黄勇,龚铭新,等. 10 s全力蹬踏自行车运动过程中股直肌sEMG活动特征研究[J]. 体育科学,2010,30(11):50-55.
[5] 张秋霞,张林,王国祥. 局部肌肉疲劳对踝关节本体感觉的影响[J]. 体育科学,2011,(3):68-73.
[6] AAGAARD P,SIMONSEN E B,ANDERSEN J L,et al. Antagonist muscle coactivation during isokinetic knee extension[J]. Scand J Med Sci Sports,2000,10(2):58-67.
[7] ADELAAR R S. The practical biomechanics of running[J]. Am J Sports Med,1986,14(6):497-500.
[8] AMIRIDIS I G,MARTIN A,MORLON B,et al. Coactivation and tension-regulating phenomena during isokinetic knee extension in sedentary and highly skilled humans[J]. Eur J Appl Physiol Occup Physiol,1996,73(1-2):149-156.
[9] ANDERSEN L L,LARSSON B,OVERGAARD H,et al. Torque-velocity characteristics and contractile rate of force development in elite badminton players[J]. Eur J Sport Sci,2007,7(3):127-134.
[10] BAZZUCCHI I,RICCIO M E,FELICI F. Tennis players show a lower coactivation of the elbow antagonist muscles during isokinetic exercises[J]. J Electromyogr Kinesiol,2008,18(5):752-759.
[11] BAZZUCCHI I,SBRICCOLI P,MARZATTINOCCI G,et al. Coactivation of the elbow antagonist muscles is not affected by the speed of movement in isokinetic exercise[J]. Muscle Nerve,2006,33(2):191-199.
[12] BERCIER S,HALIN R,RAVIER P,et al. The vastus lateralis neuromuscular activity during all-out cycling exercise[J]. J Electromyogr Kinesiol,2009,19(5):922-930.
[13] CARSON R G,RIEK S. Changes in muscle recruitment patterns during skill acquisition[J]. Exp Brain Res,2001,138(1):71-87.
[14] CRONE C,NIELSEN J. Central control of disynaptic reciprocal inhibition in humans[J]. Acta Physiol,1994,152(4):351-363.
[15] DAL MASO F,LONGCAMP M,AMARANTINI D. Training-related decrease in antagonist muscles activation is associated with increased motor cortex activation:evidence of central mechanisms for control of antagonist muscles[J]. Exp Brain Res,2012,220(3-4):287-295.
[16] DE LUCA C J,ERIM Z. Common drive in motor units of a synergistic muscle pair[J]. J Neurophysiol,2002,87(4):2200-2204.
[17] DE LUCA C J,KUZNETSOV M,GILMORE L D,et al. Inter-electrode spacing of surface EMG sensors:Reduction of crosstalk contamination during voluntary contractions[J]. J Biomech,2012,45(3):555-561.
[18] DE LUCA C J,MAMBRITO B. Voluntary control of motor units in human antagonist muscles:coactivation and reciprocal activation.[J]. J Neurophysiol,1987,58(3):211-216.
[19] DUCHATEAU J,BAUDRY S. The neural control of coactivation during fatiguing contractions revisited[J]. J Electromyogr Kinesiol,2014,24(6):780-788.
[20] ENOKA R M,BAUDRY S,RUDROFF T,et al. Unraveling the neurophysiology of muscle fatigue[J]. J Electromyogr Kinesiol,2011,21(2):208-219.
[21] FARINA D,MERLETTI R,INDINO B,et al. Surface EMG crosstalk between knee extensor muscles:Experimental and model results[J]. Muscle Nerve,2002,26(5):681-695.
[22] FARINA D,MERLETTI R,INDINO B,et al. Surface EMG crosstalk evaluated from experimental recordings and simulated signals - Reflections on crosstalk interpretation,quantification and reduction[J]. Methods Inf Med,2004,43(1):30-35.
[23] FARMER S F,GIBBS J,HALLIDAY D M,et al. Changes in EMG coherence between long and short thumb abductor muscles during human development[J]. J Physiol-London,2007,579(2):389-402.
[24] GRIBBLE P L,MULLIN L I,COTHROS N,et al. Role of cocontraction in arm movement accuracy[J]. J Neurophysiol,2003,89(5):2396-2405.
[25] HÄKKINEN K,KALLINEN M,IZQUIERDO M,et al. Changes in agonist-antagonist EMG,muscle CSA,and force during strength training in middle-aged and older people.[J]. J Appl Physiol,1998,84(4):1341-1349.
[26] HANSEN S,HANSEN N L,CHRISTENSEN L O,et al. Coupling of antagonistic ankle muscles during co-contraction in humans[J]. Exp Brain Res,2002,146:282-292.
[27] JASKOLSKA A,KISIEL-SAJEWICZ K,BRZENCZEK-OWCZARZAK W,et al. EMG and MMG of agonist and antagonist muscles as a function of age and joint angle[J]. J Electromyogr Kinesiol,2006,16(1):89-102.
[28] LEVENEZ M,GARLAND S J,KLASS M,et al. Cortical and spinal modulation of antagonist coactivation during a submaximal fatiguing contraction in humans[J]. J Neurophysiol,2008,99(2):554-563.
[29] LEVENEZ M,KOTZAMANIDIS C,CARPENTIER A,et al. Spinal reflexes and coactivation of ankle muscles during a submaximal fatiguing contraction[J]. J Appl Physiol,2005,99(3):1182-1188.
[30] LOWERY M M,STOYKOV N S,KUIKEN T A. A simulation study to examine the use of cross-correlation as an estimate of surface EMG cross talk[J]. J Appl Physiol,2003,94(4):1324-1334.
[31] MULLANY H,O’MALLEY M,GIBSON A S,et al. Agonist-antagonist common drive during fatiguing knee extension efforts using surface electromyography[J]. J Electromyogr Kinesiol,2002,12:375-384.
[32] NIELSEN J A C C. H-reflexes are smaller in dancers from The Royal Danish Ballet than in well-trained athletes[J]. Eur J Appl Physiol Occup Physiol,1993,66(2):116-121.
[33] NORDIN M,FRANKEL V H. 肌肉骨骼系统基础生物力学[M].北京:人民卫生出版社,2008:162-166.
[34] OSU R,FRANKLIN D W,KATO H,et al. Short- and long-term changes in joint co-contraction associated with motor learning as revealed from surface EMG[J]. J Neurophysiol,2002,88(2):991-1004.
[35] WANG L,LU A,ZHANG S,et al. Fatigue-related electromyographic coherence and phase synchronization analysis between antagonistic elbow muscles[J]. Exp Brain Res,2015,233(3):971-982.
[36] WARGON I,LAMY J C,BARET M,et al. The disynaptic group I inhibition between wrist flexor and extensor muscles revisited in humans[J]. Exp Brain Res,2006,168(1-2):203-217.
[37] YUKSEL M F,CENGIZ A,ZORBA E,et al. Effects of Badminton Training on Physical Parameters of Players[J]. Anthropol,2015,21(3):542-547.
[38] ZEHR E P,SALE D G,DOWLING J J. Ballistic movement performance in karate athletes[J]. Med Sci Sports Exe,1997,29(10):1366-1373.
A Comparative Study on the Coactivation of Ankle Antagonist Muscles in Badminton
Objective:The aim of this study was to examine the difference of ankle antagonist muscle coactivation between badminton players and college students and thus to explore the effect of badminton movement on the neuromuscular control and antagonist muscle coactivation of ankle joint. Methods:10 college students are in control group,8 college badminton players are in experiment group. Subjects were instructed to perform three maximal voluntary isometric contractions (MVC)and three maximal ankle dorsiflexion and plantar flexion at angular velocity of 30º,60º,120º,180º/ s. Surface electromyogram (sEMG) of tibialis anterior (TA) and lateral grastrocnemius (LG) were recorded and average EMG amplitude (AEMG) of sEMG was calculated. Results:Maximal torques of badminton players group were all higher than non-players at four angular velocities,especially on ankle dorsiflexion torque. Antagonist coactivation (AEMG) of LG was significantly lower in badminton players than non-players group at all angular velocities. On the contrary,there was no significant difference of TA antagonist coactivation between badminton players and non-players group. Conclusions:Badminton movement induces the central nervous system to modify the pattern of muscle activities to make optimal use of the neuromuscular system and decrease the antagonist coactivation of ankle plantar flexors.
badminton movement;ankle joint;antagonist muscle cocontraction;surface electromyogram
1002-9826(2017)04-0058-07
10. 16470/j. csst. 201704008
G804.6
A
2016-11-01;
2017-05-27
上海市体育局科技综合计划项目(16Z013),同济大学教学改革研究与建设项目,同济大学第十二期实验教学改革项目。
王乐军,男,实验师,博士,主要研究方向为运动生物力学,Tel:(021)65981711,E-mail:wlj0523@163. com。
1.同济大学 体育教学部运动与健康研究中心,上海200092;2.上海体育学院 运动科学学院,上海 200438;3.同济大学 医学院,上海 200092
猜你喜欢
体育科技文献通报(2022年4期)2022-10-21
汽车与驾驶维修(维修版)(2021年10期)2021-11-05
汽车与驾驶维修(维修版)(2021年10期)2021-11-05
新视线·建筑与电力(2021年2期)2021-09-10
发明与创新·大科技(2021年3期)2021-04-30
直升机技术(2020年2期)2020-06-16
中学课程辅导·教师通讯(2018年10期)2018-09-04
家庭医学(2017年8期)2017-09-06
中学生数理化·高一版(2017年3期)2017-07-08
