
早期生长反应因子对U87MG细胞Aβ40表达的影响*
2016-08-11 04:30赵莘瑜
郑州大学学报(医学版) 2016年4期
赵莘瑜,苏 刚
郑州大学第一附属医院神经内科 郑州 450052 △女,1972年2月生,博士,主任医师,教授,研究方向:神经系统变性病和脑血管病,E-mail:cindy_zhaoxinyu@163.com
早期生长反应因子对U87MG细胞Aβ40表达的影响*
赵莘瑜△,苏刚
郑州大学第一附属医院神经内科 郑州 450052△女,1972年2月生,博士,主任医师,教授,研究方向:神经系统变性病和脑血管病,E-mail:cindy_zhaoxinyu@163.com
摘要目的:研究早期生长因子EGR1对U87MG细胞Aβ40表达的影响。方法:构建EGR1表达质粒并转染U87MG(实验组),以转染空白质粒的U87MG细胞作为对照。采用qRT-PCR法检测两组EGR1 mRNA的表达;收集细胞培养液用ELISA法检测Aβ40的水平;提取总蛋白用蛋白印迹法检测BACE1和PS1蛋白的表达。结果:质粒成功构建并表达。实验组EGR1 mRNA表达水平较对照组明显增高(P<0.001);实验组细胞培养液中Aβ40水平较对照组增高(P<0.001);实验组BACE1蛋白的表达较对照组增加(P<0.001);两组PS1蛋白的表达差异无统计学意义(P=0.367)。结论:EGR1在体外可能使Aβ40表达增加,且可能与上调BACE1有关。
早期生长反应因子(early growth response 1, EGR1)基因是即刻早期基因(immediate early gene, IEG)家族成员,是含有锌指结构的核转录因子,该类基因在细胞受外部刺激后最先表达[1-2]。其在人脑内基础表达量非常低,但在一定条件下能迅速被大量诱导[2]。最初研究[3]发现其与炎症反应、免疫调节、生长分化相关。也有研究[4]发现EGR1与认知功能存在着密切联系。文献[5-6]论述了EGR1与人长时程记忆形成密切相关,EGR1在神经细胞内的表达是长期记忆形成和突触可塑性地由短期转变为长期必不可少的条件之一。研究[7]认为EGR1可能参与阿尔茨海默病(Alzheimer′s disease,AD)的发病过程。 有报道[7]证明EGR1可导致Taus蛋白磷酸化,形成神经纤维缠结,倾向于支持Tau蛋白假说;有学者[8]认为淀粉样前体蛋白(amyloid precursor protein,APP)过量表达使EGR1的表达水平降低,相反在APP基因敲除的实验动物内EGR1的表达量增高,更倾向于支持β淀粉样蛋白瀑布假说。EGR1参与AD的发病过程似乎得到了科学家们的共识,但具体的参与机制尚存争议。研究[9]认为慢性炎症、缺氧及应激等条件下可导致Aβ蛋白沉积和神经元损伤与丢失,EGR1作为一种早期生长反应因子在慢性炎症、缺氧、应激条件下表达迅速增高[10]。以上证据提示EGR1是通过调节Aβ蛋白参与痴呆的发病过程。Aβ蛋白是β淀粉样蛋白瀑布假说的分子基础[11],是APP病理代谢的产物,而BACE1和PS1是APP病理代谢过程中的关键酶[12]。作者构建了CMV-EGR1过表达载体,通过检测恶性胶质瘤(U87MG)细胞Aβ40、BACE1及PS1的表达来初步探讨EGR1对Aβ蛋白的影响。
1材料与方法
1.1主要试剂 BACE1及PS1一抗购自Abcam公司,ELISA试剂盒(Aβ40)购自上海拜利生物公司,山羊抗兔二抗及GADPH一抗购于武汉三鹰生物公司;DMEM高糖培养基购自索莱宝公司,胎牛血清购自四季青生物公司;蛋白裂解液、RNA提取试剂盒购自康为公司;Lipofectamine 2000购自Invitrogen公司;pCMV-EGR1-FLAG、空白载体CMV-EGFP-Kanamycin、XhoⅠ/KpnⅠ内切酶及Taq polymerase 由上海吉凯基因公司提供;qRT-PCR试剂盒、逆转录试剂盒、T4连接酶购自TAKARA公司。
1.2质粒构建 以pCMV-EGR1-FLAG质粒为模板,扩增目的基因。EGR1转化子上游引物序列为5’-CACATCCGATCCACACAG-3’,下游引物序列为5’-CGTCGCCGTCCAGCTAGACCAG-3’,扩增产物大小为750 bp。反应体系:ddH2O 12.4 μL,5×Taq buffer 4 μL,dNTPs(2.5 mmol/L)1.6 μL,上游引物(10 μmol/L)0.4 μL,下游引物(10 μmol/L)0.4 μL,模板1 μL,Taq polymerase 0.2 μL。反应条件:94 ℃ 3 min,94 ℃ 30 s,60 ℃30 s ,72 ℃ 40 s 30个循环;72 ℃ 5 min,割胶纯化。载体酶切:用XhoⅠ/KpnⅠ内切酶对质粒载体进行酶切。反应体系:ddH2O 40.5 μL,10×buffer 15 μL,100×BSA 0.5 μL,纯化的DNA质粒(1 g/L)2 μL,XhoⅠ(10 kU/L)1 μL,KpnⅠ(10 kU/L)1 μL。37 ℃下酶切2 h。重组质粒构建:将添加了黏性末端的EGR1 cDNA与空载体混合,在T4连接酶的作用下进行连接反应,用感受态的大肠杆菌转化,于4 ℃过夜;37 ℃培养转化的大肠杆菌1 d,在含有卡那霉素的LB培养基上筛选克隆。结果鉴定:扩大培养后,用质粒提取试剂盒提取后对转化子行PCR鉴定。送样本至吉凯生物公司进行测序鉴定,结果与GenBank上EGR1(NM_001964)比较。
1.3细胞培养、质粒转染及总蛋白的提取 细胞培养:将U87MG细胞接种于25 mL培养瓶中,置于37 ℃、体积分数5%CO2的细胞培养箱中培养,培养液为DMEM高糖培养基。根据细胞生长情况约2~3 d换液1次。质粒转染:取生长良好的U87MG细胞按照Lipofectamine 2000说明书操作进行转染。并在转染6 h后于荧光显微镜(×200)下观察,转染成功后继续培养细胞2 d后提取总蛋白。总蛋白提取:将细胞用胰酶消化2 min,转移至2 mL离心管中1 500×g离心5 min,弃上清,加入2 μL蛋白酶抑制剂2 min后加入198 μL细胞裂解液,反复吹打均匀,于冰上裂解20 min,13 000×g4 ℃下离心10 min。BCA法测蛋白浓度,将各管蛋白浓度调整至相同水平,加入lodding煮沸5 min,置于-80 ℃冰箱备用,实验重复3次。
1.4EGR1 mRNA的qRT-PCR检测 采用Trizol法提取细胞总RNA,逆转录合成cDNA,按照qRT-PCR试剂盒说明书进行操作,以GADPH为内参,引物见1.2。反应条件为95 ℃ 30 s,95 ℃ 5 s,60 ℃ 34 s重复40个循环,采用2-ΔΔCt法进行数据的相对定量分析,实验重复3次。
1.5BACE1、PS1的蛋白印迹实验 取30 μg总蛋白样品(约25 μL)于120 V恒定电压条件下电泳90 min。于220 mA恒定电流条件下将电泳过的蛋白转移至硝酸纤维素膜上。将膜置于50 g/L的脱脂牛奶中室温封闭2 h。将封闭后的硝酸纤维素膜置于2 mL单克隆抗体稀释液中4 ℃过夜。次日用TBST液漂洗3次,每次10 min。用TBST将二抗稀释2 000倍,在摇床上孵育1 h,用TBST漂洗5次,每次10 min。于暗室中用ECL发光液曝光。实验以GAPDH作为内参对照。用Image J软件进行灰度值分析,以目的蛋白与内参灰度值的比值作为目的蛋白的相对表达量,实验重复3次。
1.6Aβ40蛋白的酶联免疫吸附实验(ELISA)检测 转染成功后继续培养48 h后更换为无血清DMEM培养基,收集无血清细胞培养液。取无血清培养液用TCA进行蛋白质沉淀,4 ℃过夜。按照Aβ40的ELISA说明书步骤操作,每样本设2个复孔。实验重复3次。
1.7统计学处理采用SPSS 21.0分析,两组EGR1 mRNA和BACE1、PS1蛋白的表达以及Aβ40水平比较采用两独立样本t检验,检验水准α=0.05。
2结果
2.1EGR1转化子鉴定结果 重组质粒转化成功(图1)。测序结果与GenBank一致。
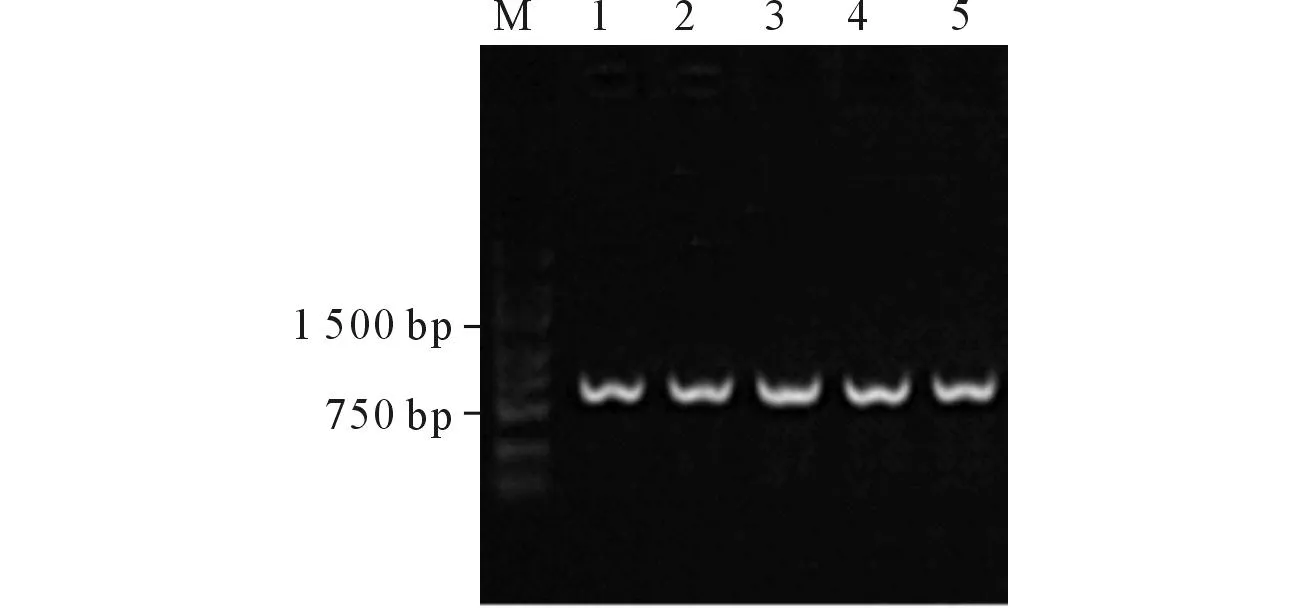
M:Marker;1~5:EGR1转化子。图1 EGR1转化子鉴定
2.2质粒转染情况荧光显微镜下可见绿色梭形细胞,表明重组质粒成功转染了U87MG细胞(图2)。

图2 CMV-EGR1-EGFP转染6 h后荧光显微镜图(×200)
2.3两组U87MG细胞EGR1 mRNA的表达和Aβ40水平情况见表1。
2.4两组U87MG细胞BACE1和PS1蛋白的表达情况见图3、4和表1。
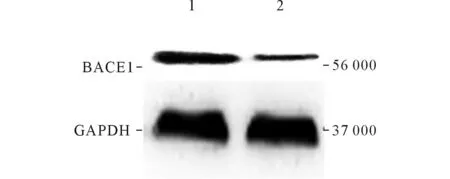
1:实验组;2:对照组。图3 各组BACE1蛋白的表达情况

1:实验组;2:对照组。图4 各组PS1蛋白的表达情况
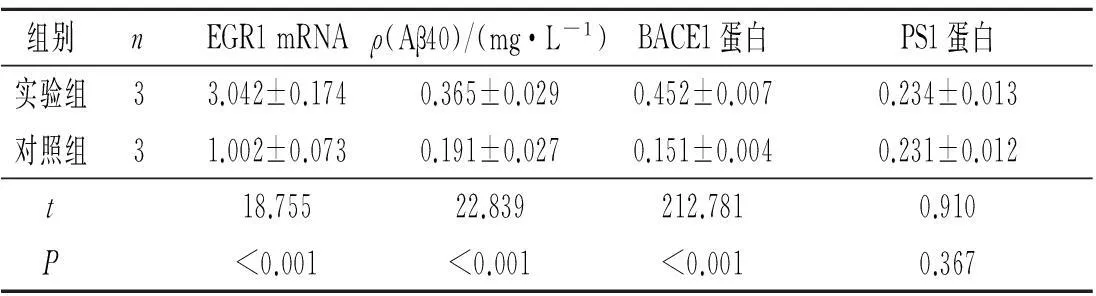
组别nEGR1mRNAρ(Aβ40)/(mg·L-1)BACE1蛋白PS1蛋白实验组33.042±0.1740.365±0.0290.452±0.0070.234±0.013对照组31.002±0.0730.191±0.0270.151±0.0040.231±0.012t18.75522.839212.7810.910P<0.001<0.001<0.0010.367
3讨论
APP的正常代谢不产生Aβ蛋白,而是经过α剪切酶的作用产生对神经细胞有保护作用的sAPPα[13];当APP进入非正常代谢途径时,BACE1作用于APP,剪切产生APP-βCTF和可溶性片段sAPPβ;APP-βCTF进一步被γ分泌酶作用,最后释放Aβ40和Aβ42[14]。γ分泌酶为四聚体结构,由PS1、APH1、PSEN2、NCT四个亚基构成,其中PS1是其催化亚基[15]。因此BACE1和PS1在APP的病理剪切过程中起着重要作用。由于Aβ42分泌量较Aβ40少很多,考虑到蛋白浓度过低时对实验结果影响较大,作者仅检测Aβ40蛋白浓度。
有研究[3]表明EGR1与AD关系密切。Hendrickx等[4]阐明了EGR1的过表达可以导致APP蛋白表达增多,在酶活性和总量不变的情况下,也可导致Aβ蛋白的增加。该研究结果显示外源性的EGR1成功转染U87MG细胞后,细胞培养液中Aβ40分泌量较对照组明显增加。研究[16]发现APP的剪切过程在很大程度上决定了Aβ蛋白的表达量,这说明Aβ蛋白的产生主要来自于APP的异常代谢。BACE1介导APP的病理剪切,并对Aβ蛋白的表达起着重要作用。PS1对Aβ蛋白的产生也起有一定的作用。因此作者推测实验组Aβ40的增多可能是由于EGR1在参与APP异常剪切的过程时,关键酶BACE1和PS1表达增加所导致的结果。
该研究结果显示实验组BACE1蛋白的表达较对照组增高。目前尚无相关研究表明EGR1与BACE1之间有明确关系。有学者[17]指出溶血卵磷脂酸能通过上调BACE1的表达影响Aβ的形成,提示EGR1使Aβ40的表达增高可能是因为BACE1的表达上调所导致。PS1是γ分泌酶四聚体的稳定表达成分,其表达的高低可以在一定程度上代表γ分泌酶的表达。虽然有研究[18]表明γ分泌酶能很大程度上影响Aβ蛋白的形成,但目前尚无明确证据显示EGR1对PS1的表达有影响。该研究结果显示两组PS1蛋白的表达差异无统计学意义,说明EGR1可能不会通过调节γ分泌酶的量而影响Aβ蛋白的表达。
综上所述,过表达EGR1可能刺激U87MG细胞内Aβ40的表达,并且可能与BACE1的上调相关。该结论尚需进一步实验验证。
参考文献
[1]KOLDAMOVA R,SCHUG J,LEFTEROVA M,et al.Genome-wide approaches reveal EGR1-controlled regulatory networks associated with neurodegeneration[J].Neurobiol Dis,2014,63:107
[2]MADDOX SA,MONSEY MS,SCHAFE GE.Early growth response gene 1 (Egr-1) is required for new and reactivated fear memories in the lateral amygdala[J].Learn Mem,2011,18(1):24
[4]HENDRICKX A,PIERROT N,TASIAUX B,et al.Epigenetic regulations of immediate early genes expression involved in memory formation by the amyloid precursor protein of Alzheimer disease[J].PLoS One,2014,9(6):e99467
[5]BOZON B,KELLY A,JOSSELYN SA,et al.MAPK, CREB and zif268 are all required for the consolidation of recognition memory[J].Philos Trans R Soc Lond B Biol Sci,2003,358(1432):805
[6]ALBERINI CM.Transcription factors in long-term memory and synaptic plasticity[J].Physiol Rev,2009,89(1):121
[7]LU Y,LI T,QURESHI HY,et al.Early growth response 1 (Egr-1) regulates phosphorylation of microtubule-associated protein tau in mammalian brain[J].J Biol Chem,2011,286(23):20569
[8]HENDRICKX A,PIERROT N,TASIAUX B,et al.Epigenetic induction of EGR-1 expression by the amyloid precursor protein during exposure to novelty[J].PLoS One,2013,8(9):e74305
[9]SY M,KITAZAWA M,MEDEIROS R,et al.Inflammation induced by infection potentiates tau pathological features in transgenic mice[J].Am J Pathol,2011,178(6):2811
[10]BHATTACHARYYA S,FANG F,TOURTELLOTTE W,et al.Egr-1: new conductor for the tissue repair orchestra directs harmony(regeneration) or cacophony(fibrosis)[J].J Pathol,2013,229(2):286
[11]BAYER TA,WIRTHS O.Focusing the amyloid cascade hypothesis on N-truncated Abeta peptides as drug targets against Alzheimer's disease[J].Acta Neuropathol,2014,127(6):787
[12]SADLEIR KR,EIMER WA,COLE SL,et al.Aβ reduction in BACE1 heterozygous null 5XFAD mice is associated with transgenic APP level[J].Mol Neurodegener,2015,10:1
[13]MORI T,KOYAMA N,SEGAWA T,et al.Methylene blue modulates β-secretase, reverses cerebral amyloidosis, and improves cognition in transgenic mice[J].J Biol Chem,2014,289(44):30303
[14]李晓秋,周中和,徐慧琴,等.淀粉养脑血管病26例尸检标本免疫组化病理研究[J].解放军医药杂志,2014,4:20
[15]ZHAO G,LIU Z,ILAGAN MX,et al.Gamma-secretase composed of PS1/Pen2/Aph1a can cleave notch and amyloid precursor protein in the absence of nicastrin[J].J Neurosci,2010,30(5):1648
[16]BRUNHOLZ S,SISODIA S,LORENZO A,et al.Axonal transport of APP and the spatial regulation of APP cleavage and function in neuronal cells[J].Exp Brain Res,2012,217(3/4):353
[17]SHI L,DONG YZ,CUI MZ,et al.Lysophosphatidic acid induces increased BACE1 expression and Aβ formation[J].Biochim Biophys Acta,2013,1832(1):29
[18]CHU J,WISNIEWSKI T,PRATICD.GATA1-mediated transcriptional regulation of the γ-secretase activating protein increases Aβ formation in down syndrome[J].Ann Neurol,2016,79(1):138
(2015-10-11收稿责任编辑李沛寰)
doi:10.13705/j.issn.1671-6825.2016.04.020
中图分类号R742
关键词阿尔茨海默病;Aβ40;EGR1;U87MG细胞
Effect of early growth response 1 on expression of Αβ40 in U87MG cells
ZHAO Xinyu,SU Gang
DepartmentofNeurology,theFirstAffiliatedHospital,ZhengzhouUniversity,Zhengzhou450052
Key wordsAlzheimer′s disease;beta amyloid protein 40;early growth response 1;U87MG cells
AbstractAim: To research the effect of early growth response 1(EGR1) on the expression of beta amyloid protein 40(Aβ40) U87MG cells. Methods: To construct overexpression plasmid of CMV-EGFP-EGR1-Kanamycin and transfect U87MG cells as the experimental group; the control group cells were transfected by the plasmids of CMV-EGFP-Kanamycin. The expression of EGR1 mRNA was detected by qRT-PCR. The culture media was collected and Aβ40 was detect by ELISA. The total proteins of U87MG cells from the 2 groups was extracted and BACE1 and PS1 was measure by Western blot.Results: The experimental group and the control group were transfected by plasmids successfully. The expression of EGR1 mRNA increased significantly in the experimental group compared with the control group(P<0.001). Average concentration of Aβ40 was significantly higher in the experimental group than that in the control group(P<0.001). There was a significant increase of BACE1 protein level in the experimental group than that in the control group(P<0.001).There was no significant difference in PS1 protein level between the two groups(P=0.367).Conclusion: EGR1 can increase the expression of Aβ40 in vitro, which may associate with up-regulating the expression of BACE1 protein.
*河南省科技厅基础与前沿项目340600531605;河南省教育厅自然科学研究计划12A320072
猜你喜欢
成都医学院学报(2022年4期)2022-08-19
湖南畜牧兽医(2021年6期)2022-01-24
中学生物学(2021年8期)2021-11-02
食品安全导刊(2021年21期)2021-08-30
江西农业学报(2021年4期)2021-04-20
三农资讯半月报(2020年11期)2020-06-21
天津医科大学学报(2019年6期)2019-08-13
中国果业信息(2019年1期)2019-01-05
中国人兽共患病学报(2018年7期)2018-07-31
中国畜牧兽医文摘(2018年6期)2018-07-28
