
大鼠δ阿片受体基因小发卡RNA 及人前脑啡肽原基因双表达慢病毒载体的构建与鉴定*
2012-12-23 05:14逄坤芳陈鹤翔卜慧莲刘希江
华中科技大学学报(医学版) 2012年3期
逄坤芳, 陈鹤翔, 杨 辉, 卜慧莲, 刘希江
华中科技大学同济医学院附属同济医院麻醉学教研室,武汉 430030
阿片类药物长期应用于慢性疼痛患者将导致药物耐受,影响治疗效果并增加副作用,其发生机制可能与μ 受体(mu opioid receptor,MOR)和δ受体(DOR)形成异源二聚体,经过细胞内吞作用,泛素化并在溶酶体中降解有关[1]。有文献证实应用小剂量的DOR 拮抗剂能抑制吗啡耐受的产生并能逆转已产生的耐受[2],表明拮抗DOR 对于抑制吗啡耐受可能有一定作用。脑啡肽作为一种内源性阿片肽,在许多动物实验中已经被证实具有一定的镇痛作用[3]。由于慢病毒载体对分裂细胞和非分裂细胞均具有较强的转染能力且转染效率高,可在细胞中稳定持久地表达[4],因此本实验拟构建具有双表达功能的pLenti-CMV-hPPE-U6-shRNA 慢病毒载体,此病毒载体含人前脑啡肽原基因(hPPE),能表达脑啡肽;同时构建的病毒中还含有特异性抑制DOR 受体表达的小发夹RNA 片段,以期将此病毒载体运用于慢性疼痛的研究,探讨其在镇痛及吗啡耐受中的作用,为找到更合理的镇痛方法提供新的思路。
1 材料与方法
1.1 主要材料
pENTR/U6vector 质粒载体、platinum pfx polymerase、pcDNA3.1(+)质粒、LR clonase Ⅱenzyme mix、pLenti7.3/V5-DEST、包装质粒、293T细胞、DMEM +10% 胎牛血清、lipofectamine 2000、Opti-MEM 培养液以及0.05% 胰蛋白酶均购自Invitrogen公司;oligo DNA 购自上海英骏生物工程技术有限公司;HindⅢ、KpnⅠ、SaⅡ、BamHⅠ以及T4DNA 连接酶均购自New England Biolabs公司;DNA 凝胶回收试剂盒和小量质粒提取试剂盒购自Axygen 公司;感受态大肠埃希菌DH5α购自天根生化公司。
1.2 DOR-shRNA干扰载体构建
1.2.1 靶向DOR 基因的shRNA 序列的合成 根据本实验室先前已筛选出的大鼠DOR 基因序列GACGAGAACTTCAAGCGCT,合成1 对shRNA寡聚单链DNA。上游:5′-CACCGACGAGAACTTCAAGCGCTCGAAAGCGCTTGAAGTTCTCGTC-3′,下游:5′-AAAAGACGAGAACTTCAAGCGCTTTCGAGCGCTTGAAGTTCTCGTC-3′。
1.2.2 DOR-shRNA 表达载体pENTR/U6vector的构建 将2对寡聚单链DNA 退火成双链,重组克隆后将退火而成的双链shRNA oligo插入shRNA 表达载体pENTR/U6vector中,构建shRNA表达质粒,并转化至感受态细胞DH5α,于37℃培养箱过夜培养。每个转化平板分别挑取3个克隆,摇菌抽提质粒后进行测序。
1.3 合成hPPE基因序列及hPPE-pcDNA3.1(+)表达载体的构建
根据hPPE基因序列(大小约为830bp),设计并合成若干条oligo进行基因合成,在序列的5′端和3′端分别加上HindⅢ和KpnⅠ酶切位点,以便亚克隆至pcDNA3.1(+)中。将合成好的oligo配成oligo mix,进行PCR 反应,电泳并回收PCR 产物。用HindⅢ和KpnⅠ对PCR 产物和pcDNA3.1(+)同时进行酶切,37℃酶切2h。电泳并回收酶切的PCR产物和载体,用T4DNA 连接酶连接回收的片段与载体,室温连接2h。取5μL连接产物转化感受态细胞DH5α。挑取多个单克隆进行测序。
1.4 pLenti-CMV-hPPE-U6-shRNA慢病毒载体构建
1.4.1 构建带hPPE 的pENTR/U6shRNA 表达载体 设计合成引物,扩增hPPE-BGHpA 片段,上游引物加Sal Ⅰ位点,下游引物加BamH Ⅰ位点。引物序列:上游:5′-ATCATCGTCGACCTTAAGCTTGCCACCATGGC-3′;下游:5′-ATCATCGGATCCCAGAAGCCATAGAGCCCACC-3′。用以上引物扩增hPPE-BGHpA(约1 000bp),回收PCR产物。用SalⅠ和BamHⅠ对hPPE-BGHpA 片段和pENTR/U6-shRNA 同时进行酶切,37℃酶切2 h。电泳并回收酶切后的hPPE-BGHpA 片段和质粒,连接回收的目的基因片段和载体,室温连接2 h,转化后测序验证。
1.4.2 慢病毒载体重组构建 将以上构建好的带hPPE的shRNA 载体与慢病毒载体pLenti7.3/V5-DEST 进行重组。25℃反应1h,加入1μL 的蛋白酶K,37℃反应10min。取转化后筛选的阳性克隆并测序验证,保留测序验证正确的LR 重组质粒。
1.5 慢病毒的包装和滴度测定
1.5.1 慢病毒包装 取处于对数生长期的293T细胞,计数后每个10cm 的培养皿接种6×106个细胞,37℃,5%CO2的培养箱中培养过夜;第2天转染前移去培养液,换5 mL Opti-MEM 培养液;制备DNA 与Lipofectamine 2000转染复合物,转染后的293T 细胞继续培养。48h 后收集细胞培养上清,3 000r/min离心10min,去除细胞和碎片,过滤;将病毒原液在50 000g下超速离心2h,去上清,重悬于800μL DMEM 培养液中,分装放置于-80℃保存备用。
1.5.2 慢病毒活性滴度测定 DMEM 培养液中加入2%胎牛血清,再添加8μg/mL 的Polybrene,混合均匀后备用。将病毒原液用慢病毒稀释用培养液10倍比稀释后,各取100μL 感染96 孔板中的293T 细胞。使100μL培养液中分别含慢病毒液2×10-3、2×10-4、2×10-5、2×10-6、2×10-7mL。吸去培养液,混匀,各取100μL加入每孔细胞中,每个稀释度设2个重复。放入37℃的细胞培养箱中培养,72h后,在荧光显微镜下观察各孔中荧光细胞数量并测定滴度。病毒滴度=各孔中表达荧光的细胞平均数/每孔中含有的慢病毒液体积。
2 结果
2.1 DOR-shRNA表达载体pENTR/U6-shRNA构建成功
测序证明重组克隆中插入片段序列与设计的寡聚单链DNA 序列一致,DOR-shRNA 表达载体pENTR/U6-shRNA 构建成功,测序结果见图1。

图1 pENTR/U6-shRNA 插入的寡聚单链DNA 序列Fig.1 Oligomeric single-stranded DNA sequence in pENTR/U6-shRNA
2.2 pcDNA3.1(+)-hPPE表达载体构建成功
电泳回收PCR 产物,结果如图2。回收PCR产物和pcDNA3.1(+)同时进行酶切,连接、转化后挑取多个单克隆行测序验证,测序结果证实hPPE序列准确插入至pcDNA3.1(+)中,结果如图3。
2.3 pLenti-CMV-hPPE-U6-shRNA慢病毒载体构建成功
设计合成引物,扩增hPPE-BGHpA PCR 结果如图4。电泳并回收酶切后的hPPE-BGHpA 片段和质粒,连接转化后挑取阳性克隆测序,测序结果显示hPPE-BGHpA 与pENTR/U6-shRNA 连接成功(图5)。

图2 hPPE基因oligo片段电泳结果Fig.2 Electrophoresis of hPPE gene oligo fragments

图3 pcDNA3.1(+)-hPPE测序结果Fig.3 Sequencing results of pcDNA3.1(+)-hPPE

图4 hPPE-BGHpA 片段电泳结果Fig.4 Electrophoresis of hPPE-BGHpA gene fragments
2.4 pLenti-CMV-hPPE-U6-shRNA慢病毒滴度测定
将成功构建的慢病毒包装所用质粒共转染293T 细胞后,在倒置显微镜下观察发现几乎所有的293T 细胞都发出强烈的绿色荧光,见图6。经计算pLenti-CMV-hPPE-U6-shRNA 病毒的滴度为1×107TU/mL。
3 讨论
经典的阿片类受体可分为3种亚型,即μ、δ以及κ型,广泛存在于脑和脊髓,可被内源性阿片肽类如脑啡肽、β内啡肽、强啡肽以及外源性阿片肽类如吗啡等激活,从而产生镇痛等作用[5]。人前脑啡肽原基因(hPPE)编码前脑啡肽,含263或267个氨基酸残基,在翻译过程经不同蛋白酶降解形成甲硫氨酸脑啡肽和亮氨酸脑啡肽[6]。脑啡肽可与μ 受体(MOR)、δ 受体(DOR)结合,参与内源性镇痛作用[7]。
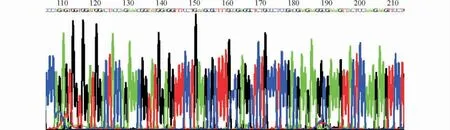
图5 pENTR/U6-shRNA-hPPE测序结果Fig.5 Sequencing results of pENTR/U6-shRNA-hPPE

图6 慢病毒感染293T 细胞的检测图(×100)Fig.6 Microscopic results of 293Tcells transduced with plasmids(×100)
目前对于急、慢性疼痛的治疗一般采用外周和中枢注入阿片肽类药物,这方法能产生明显的镇痛效果,但长期和大量用药会带来严重副作用,如药物耐受[8]。药物耐受不仅会增加药物的使用剂量,更增加了潜在的机体损伤。药物耐受的机制可能与MOR 和DOR 的相互作用有关。已经发现,MOR在细胞内吞作用后会被重新利用,DOR 却在溶酶体中降解[9],研究同时证实MOR 和DOR 可形成异源二聚体,经过细胞内吞作用,泛素化并在溶酶体中降解,而抑制DOR 引起的MOR-DOR 二聚体共降解可增强MOR 介导的镇痛作用[1]。这些均表明DOR 与MOR 的相互作用在吗啡耐受形成中起重要作用,干扰DOR 的合成可能会抑制吗啡耐受。
转基因在慢性疼痛治疗中占有重要的地位,hPPE是慢性疼痛基因治疗中一个较为理想的靶点[10]。近年的研究发现,通过病毒载体将脑啡肽原基因导入动物体内,能特异性提高蛋氨酸脑啡肽在脊髓后角神经元的表达,从而起到镇痛作用[11-12]。也有研究表明,转基因脑啡肽的表达能减轻纳洛酮诱导的吗啡戒断症状,对于治疗疼痛有良好的临床应用前景[13]。RNA 干扰技术是通过合成靶向的双链小干扰RNA(siRNA),精确降解与siRNA 序列相同的mRNA,抑制其在细胞内的翻译和表达,从而产生转录后的基因沉默,靶向特异性高。据此,我们设计了针对大鼠DOR 的干扰片段,构建DORshRNA 重组质粒,使用大鼠U6启动子指导siRNA编码序列的转录,PolⅢ识别的终止信号终止转录,符合siRNA 表达载体的设计要求。同时根据hPPE基因序列,进行基因合成,克隆至表达载体pcDNA3.1(+)中,HindⅢ和KpnⅠ酶切后进行测序。将hPPE插入已构建成功的pENTR/U6shRNA 载体中,再与慢病毒载体pLenti7.3/V5-DEST 重组,构建慢病毒载体,经酶切鉴定分析,构建的质粒重组体与预期大小一致,经测序分析无碱基突变,证实成功构建pLenti-CMV-hPPE-U6-shRNA 慢病毒表达载体,以期在激活MOR 的同时下调DOR 的表达,干扰MOR 与DOR 的相互作用,从而能持续发挥镇痛作用,降低耐受的产生。
综上所述,本实验成功构建了pLenti-CMVhPPE-U6-shRNA 慢病毒表达载体,为后续探究合理的镇痛方式奠定了基础。
[1] He S Q,Zhang Z N,Guan J S,et al.Facilitation ofμ-opioid receptor activity by preventingδ-opioid receptor-mediated codegradation[J].Neuron,2011,69(1):120-131.
[2] Abul-Husn N S,Sutak M,Milne B,et al.Augmentation of spinal morphine analgesia and inhibition of tolerance by low doses ofμ-andδ-opioid receptor antagonists[J].Br J Pharmacol,2007,151(6):877-887.
[3] Jiao L,Zhu J,Hassan M M,et al.K-rasμtation and p16and preproenkephalin promoter hypermethylation in plasma DNA of pancreatic cancer patients:in relation to cigarette smoking[J].Pancreas,2007,34(1):55-62.
[4] 卜慧莲,高峰,杨辉,等.β-arrestin 2抗基因RNA 慢病毒表达载体的构建与鉴定[J].华中科技大学学报:医学版,2011,40(1):37-40.
[5] Mansour A,Thompson R C,Akil H,et al.Δopioid receptor mRNA distribution in the brain:comparison toδreceptor binding and proenkephalin mRNA[J].J Chem Neuroanat,1993,6(6):351-362.
[6] Monstein H J,Grahn N,Ohlsson B.Proenkephalin-A mRNA is widely expressed in tissues of the human gastrointestinal tract[J].Eur Surg Res,2006,38(5):464-468.
[7] 安珂,田玉科,杨辉,等.人前脑啡肽原基因修饰的永生化大鼠星形胶质细胞株的构建[J].中国疼痛医学杂志,2005,11(3):167-170.
[8] Bailey C P,Connor M.Opioids:cellular mechanisms of tolerance and physical dependence[J].Curr Opin Pharmacol,2005,5(1):60-68.
[9] Milan-Lobo L,Whistler J.Heteromerization of theμandδ opioid receptor produces ligand-biased antagonism and alters μreceptor trafficking[J].Pharmacol Exp Ther,2011,337(3):868-875.
[10] Wilson S P,Yeomans D C,Bender M A,et al.Antihyperalgesic effects of infection with a preproenkephalin-encoding herpes virus[J].Proc Natl Acad Sci USA,1999,96(6):3211-3216.
[11] Yoshimura N,Franks M E,Sasaki K,et al.Gene therapy of bladder pain with herpes simplex virus(HSV)vectors expressing preproenkephalin(PPE)[J].Urology,2001,57(6Suppl 1):116.
[12] Yeomans D C,Jones T,Laurito C E,et al.Reversal of ongoing thermal hyperalgesia in mice by a recombinant herpesvirus that encodes human preproenkephalin[J].Mol Ther,2004,9(1):24-29.
[13] Hao S,Hu J,Fink D J.Transgene-mediated enkephalin expression attenuates signs of naloxone-precipitated morphine withdrawal in rats with neuropathic pain[J].Behav Brain Res,2009,197(1):84-89.
猜你喜欢
中学生物学(2021年8期)2021-11-02
装备制造技术(2019年12期)2019-12-25
中国果业信息(2019年1期)2019-01-05
中国畜牧兽医文摘(2018年6期)2018-07-28
食品科学(2018年10期)2018-05-23
上海建材(2017年4期)2017-04-06
现代检验医学杂志(2016年1期)2016-11-12
西南医科大学学报(2015年1期)2015-08-22
科学启蒙(2015年8期)2015-08-07
中国当代医药(2015年9期)2015-03-01
