
树木对干旱胁迫及复水的生理生态响应研究进展*
2023-12-11 03:00邓湘雯胡彦婷欧阳帅项文化
林业科学 2023年11期
游 韧 邓湘雯,2,3 胡彦婷,2,3 欧阳帅,2,3 陈 亮,2,3 项文化,2,3
(1.中南林业科技大学生命科学与技术学院 长沙 410004;2.湖南会同杉木林生态系统国家野外科学观测研究站 会同 438107;3.南方林业生态应用技术国家工程实验室 长沙 410004)
近年来,全球CO2浓度不断升高,导致气温的不断攀升,在此变化下水循环过程严重失衡进而引发各种极端气候(Bogardiet al., 2012; IPCC 2021)。高纬度地区发生高强度日极端降水的风险增加,而在热带地区降雨则减少(Sunet al., 2017)。气温的升高和极端气候的产生使许多地区(如干旱和半干旱温带地区)的干旱事件更加频繁、持续时间更长、强度更高(Dai,2013)。水分是对树木生长影响最大的限制性因子(Kaldenhoffet al., 2006),干旱事件给树木生存和发挥功能造成的最直接影响是水分缺失。因此,降水量及降水分配格局也就成为植被区划的主要生态因子之一。
干旱诱导树木死亡的现象在全球范围内频繁发生,在2018—2019 年连续发生的高温干旱之后,欧洲西部挪威云杉(Picea abies)和欧洲山毛榉(Fagus sylvatica)森林都发生了大面积的死亡(Obladenet al.,2021)。西伯利亚冷杉(Abies sibirica)森林由于严重干旱以及暴雨等极端气候事件,超过了西伯利亚冷杉的死亡阈值导致树木大量死亡(Kharuket al., 2019),类似的现象也发生在热带雨林(Phillipset al., 2009)、地中海区域及中国的东部(Allenet al., 2010;Zhaoet al.,2021),也因为长期干旱胁迫导致树木的大量死亡,给当地经济发展造成了严重的损伤。干旱胁迫不仅会改变森林的空间分布,还将对整个生态覆盖面积、林龄结构和系统的功能和服务(例如碳固持)具有重要影响(周贵尧等,2020)。森林生态系统是生态圈的重要组成部分,植物更是森林生态系统的主体。干旱胁迫下植物死亡的机制已经受到了国内外学者的关注(Kauweet al., 2021, Trugmanet al., 2021),水力失效最早被学者认定为植物在干旱胁迫下死亡的原因(Tyreeet al., 1983),在当时普遍认为木质部水力学是植物抗旱性的直接影响因素,但是随后有研究人员提出水力失效的原因并不准确,并提出碳饥饿假说(Trifilòet al., 2017)。他们认为,植物的死亡是由于在干旱胁迫环境下,植物为减少水分的散失,从而减小甚至关闭气孔,植物体为了维持自身的代谢从而消耗本身的碳,进而导致植物的死亡(Salmonet al., 2015)。目前,极端干旱导致植物死亡的生理机制仍然存在不确定性,干旱—复水的条件下树木生理生态机制变化的相关研究在国内屈指可数(颜佳滢等,2022)。在全球气候变化背景下,探讨极端干旱导致植物死亡的生理机制及干旱—复水过程中树木生理生态的变化,对未来树木的合理经营种植提供了重要的理论和现实意义。本文就国内外关于树木死亡的生理机制及干旱—复水整个过程树木生理生态的变化研究进展进行综述。
1 树木对干旱的生理调节
了解和预测干旱期间及之后树木死亡的驱动因素是一个长期存在的科学问题,从环境保护到缓解气候变化都有广泛的影响(吴海江等, 2021)。人为导致气候发生变化的长期研究使研究人员关注树木死亡机制的研究(闫明等, 2022)。然而,干旱胁迫下树木具有复杂的多维响应,人们对树木存活和死亡的生理机制的了解还存在局限。
1.1 干旱胁迫下树木的水力失效
水力失效(hydraulic failure)是最早树木在干旱胁迫下树木死亡的机制的假说,这个假说认为树木的死亡主要是木质部栓塞引起的水分运输功能失调造成(罗丹丹等, 2017)。空气栓塞的突然形成通常发生在干旱期间,在干旱胁迫下树木的水势降低使得木质部水分传输组织发生气穴栓塞,当木质部组织的张力增加超过一个额度时(称为“气种”阈值),空气被拉入管状水力管道(陈志成等, 2018)。由此产生的栓塞阻碍木质部的水分输送,将叶子与土壤中的水源隔离开来,导致树木木质部功能障碍,水分输导受阻。当树木长时间处于干旱胁迫环境下时,木质部多次发生空穴和栓塞达到阈值,最终导致树木的死亡(Xieet al.,2022)。树木体内水分的长距离运输被认为是干旱胁迫下树木死亡的主要原因(Chenet al., 2020)。
气种的产生解释了树木体内在进行长距离运输受到阻碍的原因(代永欣等, 2017a)。“水往低处流”是一个普遍的自然规律,植物体内的水也是由水势高的器官流向水势低的器官。树木叶片在进行蒸腾作用时,水分的散失导致叶面水势降低,从而对树木内的水分流动提供吸力,通过树木体内的导管以及其他组织器官逐渐的将拉力传导到根部。根部受到低水势提供的拉力后,从土壤中吸取水分,运输到叶片从而维持水分的平衡(代永欣等, 2017b)。在干旱环境下,树木体内有极强的压力,木质部导管组织发生气穴栓塞。在干旱胁迫过程中,土壤含水量下降,木质部中的导管处于极强的负压力下,导管发生气穴栓塞致使木质部功能失效,水分传导受到限制(万贤崇等,2007)如图1。
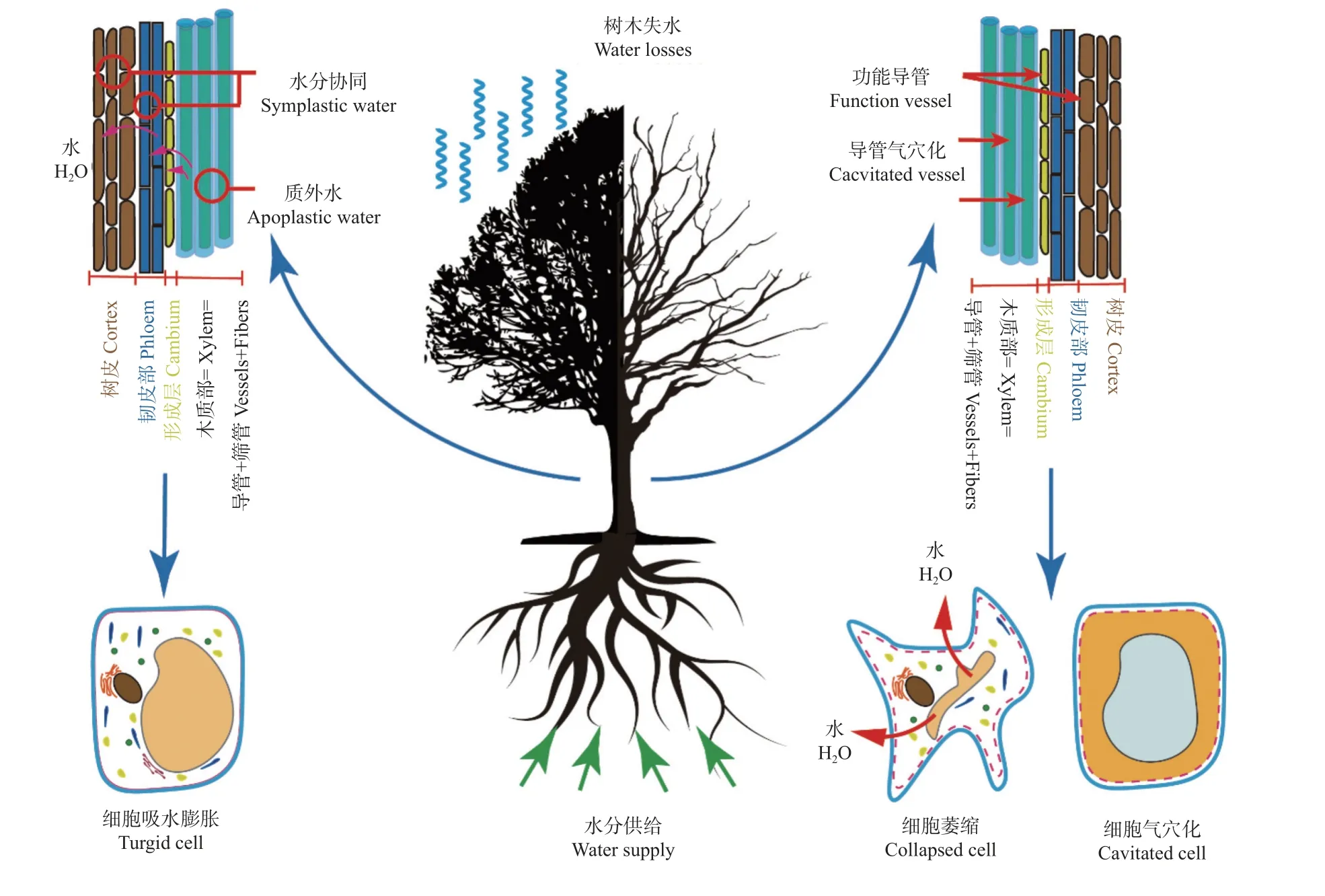
图1 水力失效假说示意图(仿Mantova et al., 2022)Fig.1 Schematic diagram of hydraulic failure hypothesis (Modified from Mantova et al., 2022)
干旱胁迫下树木的水力失效已经取得了系列进展 (刘丽燕等, 2020)。McDowell 等(2008)综述了不同类型干旱下树木的死亡机制,探讨了等水植物和异水植物的死亡率与干旱强度和持续时间的关系,认为当干旱强度增加时,干燥曲线的斜率大,在这种情况下异水植物更容易死亡;反之亦然。Choat 等(2012)将研究对象进行了划分,认定在野外自然条件下,被子植物水力学安全范围较小,更容易因水力失效死亡,被子植物短时间内的栓塞修复能力强,其具有较高的旱后恢复能力;而裸子植物正好相反,抗栓塞能力较强,栓塞修复能力相对较弱,水力失效可能不是其死亡的原因(Venturaset al., 2021)。在不同的生境中,人工生境植株比自然生境植株水力效率弱,但栓塞抗性强(韩旭丽等,2022)。在对亚马逊森林对树木的死亡原因进行探究发现,在土壤水分有效性降低10 年后,试验干旱森林中死亡造成的生物量损失大幅增加(Rowlandet al., 2015)。在中国北方的干旱环境中,研究表明水力通道的水力失效是导致树木死亡的主要原因,气孔安全边界比传统的木质部栓塞阻力和水力安全边界更能预测树木死亡(Chenet al., 2019)。
木质部水分传导组织在应对干旱胁迫下的能力因树种的差异而不同。木质部导水率丧失百分率PLC(percentage loss of conductivity)是评估水力失效最典型的方法。木质部导水性丧失50%时的水势(P50,MPa)是栓塞抵抗的常用指标,当水势低于P50之后,木质部的水分运输功能就明显受到损害,并且植物存在加速栓塞的危险。当树木长时间处于干旱胁迫环境时,木质部导管产生栓塞超过阈值,导致长时间的水分运输速率低、器官的死亡、最终整个植株的死亡(Choatet al., 2012; Xieet al., 2022)。
一些树木在死亡时体内PLC 虽然上升,但是增加的程度不相同,不同树种的安全水力范围也存在差异,所以并不能通过PLC 直接对植物死亡的原因进行判定。树木水力失效的主要原因是组织中缺乏用来渗透调节的碳储量,即渗透调节失败引发的水力失效(Sevantoet al., 2014)。当树木处于干旱胁迫环境下时,树木会调节自身的叶片气孔导度减少在蒸腾作用下的水分流失,但与此同时,气孔调节会减少二氧化碳向叶片的扩散,降低树木的光合作用,有机物合成不足,长此以往会使树木因为碳饥饿死亡(Huet al.,2022)。因此,碳饥饿也可能是树木在干旱胁迫下死亡的原因,水力失效并不能作为树木死亡的唯一理论依据(马玥等, 2021)。
1.2 干旱胁迫下树木的碳饥饿假说
碳水化合物是植物光合作用的主要产物,不但是组成细胞结构的主要成分,也是植物新陈代谢过程中重要的能源物质,对于维持树木正常的生理活动具有非常重要的作用 (Liet al., 2008)。利用树木器官中非结构性碳水化合物(non-structure carbohydrate,NSC)浓度的变化来评估碳饥饿(carbon starvation)。当植物合成的碳水化合物无法满足体内新陈代谢的需求时,非结构碳库不断被消耗,使树木长期处于碳供不应求的状态(McDowellet al., 2008)。该假说认为,当植物处于干旱胁迫环境下时,植物通过调节气孔来控制水分的蒸发散最大限度地减少了水力失效。当气孔关闭时,会影响到植物与外界环境的气体交换,从而影响植物的正常光合作用,光合作用的原料减少,碳水化合物合成受到了阻碍,但植物体为了维持正常的新陈代谢和呼吸作用依然会消耗体内的碳使非结构碳含量下降,植物在这个情况下缺乏能量,或者遭受昆虫或病原体的攻击,无论哪种情况先发生都将导致植物的死亡(韦景树等, 2018; Huet al., 2019; 胡彦婷等,2015)。相反,如果不能保持木质部水张力低于其空穴化阈值,就会导致栓塞,如果不修复,最终会导致树木水力失效、干燥和死亡(陈志成等, 2016)(图2)。
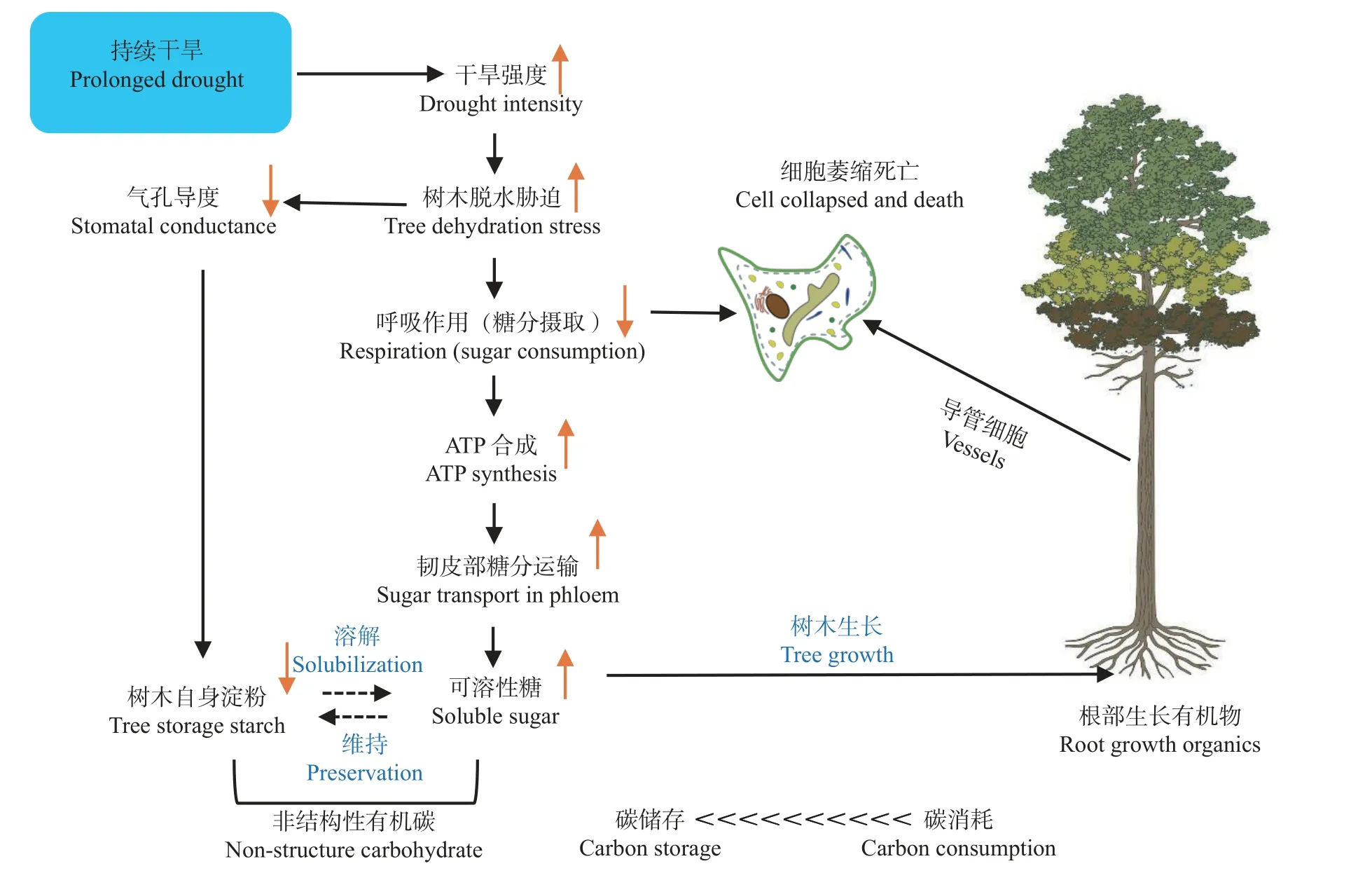
图2 碳饥饿假说示意Fig.2 Schematic diagram of the carbon starvation hypothesis
有些研究试图通过这一假说总结和解释现有关于植被死亡的现象,其中有大量的间接研究,但缺乏真实的、系统的检验(代永欣等, 2017b; 沈超等, 2019;Saviet al., 2019 )。关于利用碳水化合物的代谢限制,目前的研究还存在不确定性。Marshall 等(1985)发现遮荫的树木消耗了体内所有的淀粉,然后死亡,但树木的总糖量仍然远远高于零。侧木桉(Eucalyptus sideroxylon)幼苗在不同二氧化碳浓度和环境温度的条件下,死亡率存在差异 (Zeppelet al., 2019)。研究结果表明,气温升高加速了淀粉的消耗和随后的死亡率,这与Adams 等(2009)的最新研究结果一致。因此,并非所有碳水化合物都能被利用,特别是在干旱期间。Klein 等(2011)报道了植物在干旱状态下,与高温相联系的光抑制作用或者呼吸作用会进一步加重植物的碳饥饿。
只有当从光合作用和储存中获得的碳水化合物的可用性不等于为维持新陈代谢而消耗的碳水化合物时,组织中的碳水化合物浓度才应该下降(王凯等,2021)。因此,碳饥饿的主要驱动因素是光合作用速率降低,实际上在胁迫早期增加了碳水化合物的存储分配(段洪浪等, 2015)。类似于在细胞水平上无法利用碳水化合物,韧皮部在干旱期间的运输失败也可能会加剧碳饥饿。干旱可能通过多种机制减少韧皮部的运输,如降低碳水化合物的装载和卸载(包括降低库活性),或通过增加树液黏度降低韧皮部电导(Brodribbet al., 2016)。有研究表明,在干旱期间,光合作用下降的速度快于同化物质的运输,但其他的则相反(Zeppelet al., 2011)。然而,这些研究达不到致命的干旱胁迫,因此模式的限制可能会让结果具有不确定性。
到目前为止,已经有很多关于碳饥饿致使树木死亡的研究,其研究对象在整个生命周期NSC 的含量明显下降,从而判定树木产生了碳饥饿现象(Daiet al.,2018),但是也有相关的研究表明树木在死亡时,体内的NSC 并没有完全消耗或者消耗不明显,从而认为树木并没有发生碳饥饿(Rowlandet al., 2015)。由此可见,只用碳饥饿来解释树木的死亡也是不全面的(万贤崇等, 2007)。
1.3 树木死亡的其他机制
由于碳—水相互作用密切相关,单靠水力失效和碳饥饿假说很可能过于简单和非排他性,无法解释干旱导致的死亡率。水力失效和碳饥饿之间的耦合作用也是树木死亡的一种机制,通过对比边材面积/总叶面积的Huber 值,研究了干旱诱导幼树死亡的生理过程,发现在树木砍伐初期存在水力失效和呼吸减少,后期碳饥饿导致树木死亡(Konoet al., 2019)。其主要还是因为水力导度的下降导致木质部栓塞与光合速率的下降之间的耦合作用(Limousinet al., 2013)。树木的直径大小对水力失效跟碳饥饿响应也会存在差异,直径小的树木会更加敏感 (Pausaset al., 2016)。除了树木本身的生理因素外,生物因素也应该被考虑在内,树木在干旱胁迫的环境下,自身抵抗病虫害的能力下降,这将加剧树木的死亡进程(Garcia-Forneret al., 2015)。
2 树木不同组织的抗旱策略
2.1 树叶的抗旱性
叶片是植物与外界环境密切接触的重要器官,也是进行光合作用和蒸腾作用的主要器官,其组织结构和生理特性易受环境因子的影响,也最能反映植物对环境的适应性(万贤崇等, 2007)。在干旱胁迫环境下的叶片形态结构(叶面积、叶片厚度和气孔密度等)、解剖结构和生理功能(相对含水量、光合速率、抗氧化系统和渗透调节物质等)均会发生一系列适应性变化,从而使树木能在干旱胁迫环境下生存。在干旱胁迫环境下的树木通常叶片较厚且有毛等覆盖物,叶片面积较小以减少蒸腾作用减少水分的流失(董建芳等,2009; Huet al., 2018a)。目前,对于植物抗旱性评价的方法很多都是通过不同组织器官进行综合评定,叶片在植物抗旱性评价上应用最广。
有研究通过隶属函数法综合评价树木叶片的解剖结构特征发现具有良好抗旱性物种有较发达的角质层,主脉较发达,维管束由木质部、韧皮部和形成层构成,外部由薄壁细胞构成的维管束鞘细胞包围(李鸿雁等, 2020)。叶片的生理特性也能反映叶片的抗旱机制,通过观察叶片的相对含水量和水势得出结论,植物叶片的相对含水量和水势能直接反映植物体内水分情况,干旱胁迫会导致植物叶片的相对含水量和水势降低,一般认为植物叶片相对含水量降幅越小表明叶片保水能力越高,对干旱胁迫的适应能力也就越强(熊仕发等, 2020)。
2.2 树茎的抗旱性
树木的茎是输送水分和营养的重要器官之一,其木质化程度和输导组织等会影响树木在干旱胁迫下的生存能力,不同种类的植物,具有不同的茎干结构差异(陆世通等, 2021)。水分和无机盐从根部吸收经茎木质部中的导管运输至叶片,保证其正常的生理代谢功能;茎中韧皮部及其他组织的水分和无机盐则由木射线细胞横向运输。
树木的向上生长发育和保持长期的有效生长均需要密度较高、质地较硬的茎干提供结构支撑(杨冬梅等, 2012)。树木管道运输的结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡关系导致在整体组织水平上的效率—安全权衡。树木导管附近还分布着较多的厚壁细胞,薄壁细胞相对较为少见,这些厚壁细胞除了部分为木纤维细胞外,另一部分有可能为纤维状的木薄壁组织,称为假薄壁组织,这种结构有利于贮藏更多的水分和营养物质,同时加强了茎的机械功能以提供支撑,保障树木体内水分和营养物质运输功能的正常运行(万贤崇等,2007)。
树木木质部的厚壁细胞和薄壁细胞,其主要作用是输导水分和机械支撑,在水分胁迫情况下茎中薄壁细胞层数和皮层厚度增加,使其在干旱胁迫下仍能储存足够的能量,以提供植物正常的生长。干旱地区植物木质部及根系都较为发达,表明在植物处于胁迫环境时,其强大的输水组织具有弹性,能很好适应环境。木质部密度更高的物种通常具有更浅的根系,或者深植于更干的土壤中,这与当木质部水势下降的时候木质密度用于保护输导性的事实相一致(Westobyet al.,2006)。
2.3 树根的抗旱性
在自然环境中并非所有树木根系都能得到充足的水分,在干旱和半干旱地区大部分根系经常处于水分亏缺状态 (赵军营等, 2005)。干旱胁迫环境下植物可以通过根与地上部分之间的长距离信息传递遥控气孔的行为, 使植物在没有真正受到干旱的伤害时快速地调整植物整体的水分关系以及碳水化合物代谢,总的趋势皆为淀粉的累积下降,而积累的可溶性糖增多,并伴随有机酸、游离氨基酸和脯氨酸等的变化, 从而避免或延缓干旱对植物造成的严重影响或伤害 (陶佳等, 2015;刘静等,2008)。
树木根部的周皮既是植物的保护组织,同时也是营养贮藏组织(赵军营等, 2006)。根的解剖结构不但体现树木根部的生长发育水平,而且与水分吸收有密切联系,其木质化程度、导管直径等都会影响植物适应干旱的能力(赵祥等, 2011)。周皮木栓层细胞的细胞壁完全栓质化,不透气、不透水,能够有效减少体内水分的流失,又杜绝了外界可能产生的不确定因素,保证植物根的生理代谢活动的正常进行(赵高卷等,2016)。木栓内层是周皮组织储存功能的结构,栓内层细胞具有浓厚的细胞质,有助于提高内部的渗透压使得细胞持水能力得到提高,栓内层细胞越发达说明树木储存物质的能力越强。
韧皮部中韧皮薄壁细胞与栓内层细胞功能相似,具有贮藏营养物质以及提高根系持水能力的功能,筛管将叶器官合成的同化产物运输至根系,韧皮射线细胞则负责将产物运输至根中的木质部内(郭建荣等,2017a)。因此,根中的韧皮部是影响其自身水分供应和生理代谢活动正常进行的关键因素,反映根系运输有机产物的水平(郭建荣等, 2017b)。
能适应干旱胁迫环境的树木根结构一般都具有较大的根直径、维管柱直径、木质部与根横切面面积比值、导管直径和导管密度。木质部是根中重要的运输器官,其中导管将根系吸收的水分和无机盐向上运输至地上部分,木质部越厚,导管越丰富,导管孔径越大,运输水分的阻力越小,输水效率越高(王林等,2016)。
3 水力分割
植物水力结构是植物为满足其自身在特定环境条件下生存竞争的需要而形成的不同形态结构(潘天天等, 2020)。导水率是指水通过植物茎叶等物质时的效率。树木中的水流可以影响其他生理过程,如气体交换和生长。在土壤—植物—大气连续统中,植物体内的水分从复水潜力低的地区流向复水潜力高的地区。“水力分割”假说(HSH)意味着植物中生命周期短的“消耗”器官(如叶片)和生命周期长的“累积”器官(如茎)中的水力传导可能会解耦,叶片中的阻力成为瓶颈或“安全阀”(Pivovaroffet al.,2014)。分割原理后来又分为两种不同的机制:水力分割和脆弱性分割(Charrieret al., 2016)。
在遭受干旱胁迫时,茎干导水性较高的树木主要依靠较低的叶片导水性来限制水分流失。脆弱性划分是指远端器官比基础器官更容易发生空化。叶片相对于树茎干来说对水分的供给更加敏感,更容易受到水分变化的影响(Jianget al., 2022)。当干旱胁迫强度增加时,叶片关闭气孔以减少蒸腾耗水,树木的器官由于水势差会依次发生栓塞。Scholl 等(2021)研究发现,在树木的叶柄空化或叶空化会先于树茎干和树根发生,从而导致干旱落叶。
4 安全边界
4.1 水力安全边界
水力安全边界也是研究树木死亡时的重要水力特征,用于评估干旱条件下植物物种的水力风险和策略。植株水势最小值反映了植株在最干旱时期的水分状况。临界点(通常为P50或P88)与最小季节性木质部水势之间的差值,即植物水力安全边界(HSM)(Zhuet al., 2019)。
HSM 反映了植物在自然最低水势条件下的木质部栓塞,并评估了干旱期间水分流失的风险,与植物的水分利用策略有关 (Choatet al., 2012)。先前的研究表明,HSM 是树木死亡率的主要预测因子(Chenet al., 2020)。低HSM 表明木质部水势接近水分胁迫的阈值,反之亦然(Anderegget al., 2012)。总体而言,针叶树种的HSM 高于大多数被子植物种,但也有研究表明地中海区域的被子植物却比针叶树种高(Meinzeret al., 2013)。耐荫物种的HSM 一般比先锋物种更高,水分利用策略更保守,总的来说,不同树种之间的HSM 存在显著差异。
4.2 气孔安全边界
在水力安全边界的基础上,也有研究人员认为在自然条件下,水势的最小值通常很难确定,因为它需要在季节尺度上进行长期连续的测量,并且当遇到不同程度的干旱时,得出的结论可能会有所不同。因此,气孔调节与木质部栓塞的相互作用使树木抗旱性的评估变得难以确定。Skelton 等(2015)提出气孔安全边界(SSM),其定义为气孔关闭时的水势和栓塞阈值时的水势之间的差异。与水力安全边界中很难确定的最小水势相比,SSM 只需要测定气孔关闭时的水势。将气孔调节与木质部对栓塞的抵抗结合在一起,有助于建立一个更全面的体系来表明植物对干旱的适应。据Chen 等(2019)发现,气孔关闭时的水势范围为-0.65~-5.32 MPa。Martin-StPaul 等(2017)通过研究气孔对干旱和栓塞抗性反应相关的功能性状的影响,发现气孔关闭时的水势最小可到-5.54 MPa,并发现大多数物种气孔关闭时的水势高于P50。
5 干旱—复水对树木的响应特征
判定树木是否死亡一般通过评估该树木恢复其关键生理功能的能力,如与环境的物质和能量交换或能否在下一个生长季节产生或再生新的器官或组织(Anderegget al., 2012)。当植物遭受干旱胁迫后,体内的水分平衡被破坏,并引起气孔关闭和光合速率下降。随着干旱的加剧,植物体内会产生和积累大量的活性氧,使得叶绿体结构被破坏,光合作用的相关酶失活或变性,并最终抑制植物的生长。旱后复水(rewatering)能够使植物的生理功能得到恢复,可在一定程度上弥补干旱对植物造成的伤害,对树木的形态特征、生理特征都造成影响。
5.1 复水后对树木生长形态的影响
干旱和复水在自然界中是循序出现的 (Johnsonet al., 2018),树木在干旱胁迫下,由于叶片气孔的关闭,导致树木不能进行正常的有机物合成,只能通过消耗自身的营养来维持正常的代谢活动。干旱胁迫下树木种子发芽率降低,株高和总生物量降低,分枝密度减小,叶片容易萎蔫,叶面积减小,甚至叶片脱落,树木在持续干旱下会将更多的光和产物分配至根系,提高根系生物量(Liet al., 2019)。Zheng 等(2017)对新疆梓树(Catalpa bungei)物种的循环干旱试验,发现连续干旱和复水对植株生长、叶片和根系参数的累积功能效应是适应连续干旱和复水的有效机制(Huanget al., 2019)。
有相关学者提出,干旱—复水后的树木根系生长发育存在明显的差异(Zadwornyet al., 2011)。研究表明,干旱—复水后树木根系活力和渗透调节物质得到了提高,降低了抗氧化酶的活性,具有更强的抵御干旱胁迫的能力,复水对树木根系总长、根体积、根系平均根直径等根系指标具有促进作用。干旱—复水后树木幼苗的细根长、根直径、根干质量增加,分支强度等根系性状均有增强,从而提高了树木幼苗的存活率(Liet al., 2018)。但是在重度胁迫处理下复水,总根长、根表面积、总根体积和根冠比均有所减小,表明树木在复水后其根系的恢复生长存在阈值效应(Wanget al., 2021)。
干旱—复水处理抑制新生叶片的形成,促使叶片平均叶面积高,叶片数量减少。也说明植物苗期受到干旱胁迫会通过减少新生叶片数量,来降低叶片的蒸腾耗水,从而提高植株对外界干旱环境的耐受能力(Hájíčkováet al., 2017)。补偿效应最初是指受到害虫取食的作物所具有的一种自发性自我弥补损害的能力。后来一些学者认为取食只是影响植物生长的多种逆境因子之一,因此,补偿效应研究不应该仅限于昆虫的取食。后来研究发现植物在干旱—复水后也存在补偿效应(胡田田等,2005)。通过叶片数量变化可以看出,复水一段时间后叶片的数量开始增加,但增加幅度比较平缓,这表明植物干旱胁迫后及时复水植株能够表现出一定的补偿效应。干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(Liuet al., 2001)。并且,复水后植物的生长和生理活动是否完全恢复,恢复的速度和程度以及对干湿循环的适应能力在很大程度上取决于之前的干旱强度和持续时间、物种和遗传类型,以及干-湿循环模式有关(Marronet al., 2003)。Acevedo 等(1971)研究发现,植物经受一定强度的干旱胁迫后复水,因干旱造成的生长抑制及生物量的损失等可以弥补,复水后植物会出现短暂的加速生长。当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力。
5.2 复水后对树木生理指标的影响
树木对干旱的适应不仅表现在胁迫过程当中,在干旱胁迫后复水过程中植物的生理特征能否恢复也是对逆境适应的重要体现(Xuet al., 2010),同时是评价树木耐旱的一个重要方面。有研究表明,树木在遭受干旱胁迫后进行复水处理,生理指标基本能恢复到正常水平(Correiaet al., 2014),干旱—复水后树木生理恢复机制研究主要集中在叶片光合特性、叶绿素荧光参数等方面(Laurianoet al., 2004)。树木复水后水势增加,从而使气孔开放和生长恢复(Galleet al.,2007),树木进行光合作用、气孔导度或叶片水势的恢复时间尺度可能不同,从几天到几周不等(Zanget al.,2013)。
树木复水后光合响应参数能较快恢复至正常状态,严重缺水时光合参数虽然显著下降,但是叶绿体没有大量降解(Creechet al., 2011)。复水后树木中的PSⅡ反应中心开放比例增加,原初光能转化效率升高,光合机构逐渐恢复至正常水平。侧柏 (Platycladus orientalis)经干旱胁迫后光合速率随着胁迫程度的加剧逐渐降低,胁迫程度越小其恢复能力越强,也发现侧柏幼树光合速率、蒸腾速率在复水24 h 后均有不同程度的恢复(张玉玉等,2021a)。
树木复水后,FV/FM呈现先增加后减小的趋势,土壤干旱胁迫对树木幼苗FV/FM的影响是可逆的。复水后侧柏幼苗Yeild 值不断升高。植物非光化学淬灭(NPQ)在复水后有缓慢的下降趋势(张玉玉等,2021b)。这一结果与油菜(Brassica napus)叶绿素荧光参数的响应和枫杨(Pterocarya stenoptera)幼苗经土壤湿干交替变化叶绿素荧光的响应结果一致 (蒙祖庆等,2012;王振夏等,2013)。树木幼苗叶绿素荧光参数在复水后得到恢复,虽然干旱对光合作用组织器官造成了一定的破坏,部分树木抗干旱能力强,前期干旱胁迫对其没有造成不可逆的破坏,通过后期复水得到恢复,并出现补偿现象。
在干旱胁迫下,光合作用组织器官受到可逆损伤,但这种损伤并不影响复水后气孔的重新开放(Liet al.,2016)。此外,Brodribb (2009)提出,叶片气体交换特性在水力特性恢复之前不会恢复。与此同时,根系溶质仍有显著的积累,一种显著的机制可以解释为维持较高的根系水分传导性或渗透调节(Arango-Velezet al., 2011),即通过邻近的薄壁组织和韧皮部分泌的渗透素进入被栓塞的木质部,从而降低渗透势。然后在管道中诱导水流和正压积聚 (Saviet al., 2016; Huet al., 2018b)。光合作用恢复后主要的渗透物质也会转变为溶质。笔者的研究结果支持木质部水分传导性与叶气交换和地上水势的相互作用有助于幼苗最终从干旱中恢复。
虽然研究人员对经历干旱胁迫后复水期的树木响应机制进行深入研究,但是由于过程的复杂性,树木从干旱中恢复的能力和涉及的过程仍不清楚,研究人员以多年生作物为研究对象,针对于水分胁迫后的机制响应进行研究(Souzaet al., 2004)。相关研究表明,物种抗旱性和干旱恢复程度的差异与各种生理、形态和生化因素有关,包括根系分布、根系直径、渗透调节(OA)、糖和有机溶质积累(DaCostaet al.,2006)。Pou 等(2008)研究表明,葡萄属植物在水分亏缺期后可以提高其水分利用效率(WUE)。然而,Gomez-del-Campo 等(2007)在比较两种应对压力能力不同的葡萄品种时并没有得到同样的结果。
6 主要结论及展望
干旱导致的大面积树木死亡已经成为一个对环境和经济都具有毁灭性的生态效应。研究人员已经根据各项研究结果明确了树木生理学在了解干旱导致的死亡率方面起到了至关重要的作用。树木死亡率在生理上是由分生组织细胞活力所决定。然而,干旱对这些细胞的影响以及随后树木的恢复仍有待研究。由于最近的各种研究已经确定了水力功能损失与干旱引起的树木死亡率之间的显著相关性,目前,通过评估水力破坏对不同树木器官和组织水分含量的影响,特别是对分生组织细胞的影响,来确定这两个过程之间的联系是至关重要的。本文综述了在干旱胁迫下树木死亡原因的3 种假说,根据前人的研究分析了3 种假说的具体生理过程及缺陷,在此基础上论述了干旱—复水整个循环中树木各个组织器官对环境条件发生改变时的响应。本文也探讨从树木脱水开始,特别是当树木在树茎干达到木质部水力衰竭时,遵循分生组织细胞脱水的动态是解决与干旱致死亡相关的关键生理问题。基于以上研究,未来的研究趋势主要包括:1)进一步加深对树木在干旱胁迫下死亡机制的理解,选取全球不同区域、不同的树种进行相关研究,验证与完善水力失效,碳饥饿假说等科学假说,尤其是确定碳饥饿对树木死亡的定量化贡献;2)对树木脱水时导管栓塞及干旱复水树木生理恢复的阈值进行相关研究,构建并完善树木死亡机制模型;3)从细胞、分子等微观角度探讨树木干旱胁迫下死亡的机制;4)树木在干旱胁迫下还会受到温度、气压等其他环境因子交互的影响,这一新的科学问题也应该成为未来的研究方向之一,这将为全球树木的管理经营提供理论性的指导。
猜你喜欢
植物研究(2021年2期)2021-02-26
食品安全导刊(2020年33期)2020-12-26
山东农业科学(2017年4期)2017-05-18
水利科技与经济(2016年3期)2016-04-09
水利科技与经济(2016年4期)2016-04-09
水利科技与经济(2016年2期)2016-04-09
天津农业科学(2015年11期)2015-12-03
江苏农业科学(2015年4期)2015-06-15
江苏农业科学(2014年10期)2014-11-22
江苏农业科学(2014年9期)2014-11-15
