
生物炭和有机肥对毛白杨人工林地木质分解及土壤养分循环相关酶活性的影响*
2023-12-11 03:00赵蕊蕊
林业科学 2023年11期
赵蕊蕊 刘 勇 王 凯
(北京林业大学林学院 北京 100083)
毛白杨(Populus tomentosa)分布地域广泛,是我国北方速生丰产林建设的重要树种。据第九次全国森林资源清查结果,我国杨树人工林面积为757 万hm2,蓄积为5.46 亿m3,占人工乔木林面积和蓄积的13%和16%,是人工栽培数量最多的阔叶树种(国家林业和草原局,2019)。然而,长期单作、连作和短轮伐等经营方式导致人工林出现了不同程度的地力衰退(王文波等,2016;杨承栋,2022),严重影响了林木生长和森林立地可持续经营。土壤有机质具有降低土壤压实和侵蚀、增强土壤持水能力和维持立地生产力等作用,是森林土壤养分循环和根系呼吸等土壤过程的重要枢纽,也是衡量土壤质量的重要指标 (Jaroszewiczet al., 2021; Bhattacharyyaet al., 2022)。提高土壤有机质是提高森林土壤健康和立地生产力的关键措施(Page-Dumroeseet al., 2021a;杨承栋,2016)。
生物炭是有机物质在限氧、热解条件下生成的一种难降解碳形式(Liet al., 2022),其在提高农林业土壤有机质含量、增加土壤碳固存以及提高作物产量等方面发挥巨大作用(Palviainenet al., 2020; Criscuoliet al., 2021; Yuet al., 2021)。研究表明,生物炭可以通过改善林地土壤理化(密度、水分特性、pH、有机质含量和土壤肥力等)和生物学特性(微生物群落组成和结构等)来减缓人工林地力衰退和维持人工林长期生产力(殷丹阳等,2016)。有机肥(厩肥、园林废弃物等)因含有植物生长需要的大量元素、密度小等特点常用于改善林地土壤结构(胡文杰等,2021)。生物炭和有机肥在提高土壤有机质含量(Liet al., 2018)、促进团聚体稳定性(Aiet al., 2018)、增加养分保持率(Seyedsadret al., 2022)、调 节 土 壤pH(Wanget al.,2021)、增加土壤阳离子交换能力和酶活性(Renet al.,2021a)以及通过改变微生物群落结构来改善土壤功能(Lehmannet al., 2011)等方面的作用已经得到充分研究。例如,生物炭和有机肥搭配施用能显著提高毛白杨幼树生长季土壤氮矿化速率(王凯等,2022)、改变土壤养分(氮、磷和钾)含量和酶活性(Zhaoet al.,2022)。然而,土壤理化性质的改变是动态过程,定期取样监测土壤性质的研究往往受限于测定指标与取样时期,不足以反映土壤质量的长期综合变化。另外,通过土壤改良剂或其他森林经营管理措施提高土壤质量对制定合理的森林立地可持续经营政策至关重要,但“土壤质量”仍是个模糊概念,无法得到直接和充分监测(Janzenet al., 2021),需要更综合性的指标来评估森林经营管理效果。
林地地表和地下的木质分解是陆地森林生态系统中的一个关键土壤过程,分解过程中释放的碳、氮、磷和钾等元素是土壤微生物和植物的主要营养来源,对提高森林生产力至关重要 (Berget al., 2003; Martinset al., 2021; Gottschallet al., 2022)。此外,木质分解速率是土壤非生物(温度、湿度、团聚体和养分有效性等)和生物(微生物生物量、多样性和组成等)因素共同作用的结果(Middletonet al., 2021; Konget al., 2022),土壤性质的改变能直接或间接引起微生物功能及分解速率的改变。因此,林地内木质分解速率可综合反映土壤改良剂或其他森林管理措施对土壤质量的影响 (Finéret al., 2016; Page-Dumroeseet al., 2021b; Wanget al., 2020)。
除土壤条件外,基质质量(碳、氮、木质素含量、碳氮比和木质素∶氮等)是影响特定气候条件下有机物质分解速率的另一重要因素(Viottiet al., 2021;Rischet al., 2022)。研究表明,碳氮比大于27~30∶1的基质会抑制异养生物的定殖(Zimmermanet al.,1995; Kayeet al., 1997);木质素∶氮含量比值高的针叶树种往往较落叶树种分解缓慢 (Wanget al., 2020)。然而,土壤改良剂或其他森林管理措施也可以通过改变凋落物(叶、根系等)性质间接影响其分解速率(Renet al., 2021b),这样就使得很难区分有机物质分解差异多大程度上源于土壤改良或其他森林管理措施引起的土壤质量的改变。利用 “标准规格”的不同质量(碳、氮和木质素含量等)木质作为林地内长期分解材料可以保持基质质量恒定,探讨森林管理实践引起的土壤非生物和生物变化。例如,利用统一规格美洲山杨(Populus tremuloides)和火炬松(Pinus taeda)木条在林地表面和0~20 cm 土壤内的分解,学者评价了皆伐(Finéret al., 2016)、不同恢复疏伐(Wanget al., 2019)及其他土地管理措施(Page-Dumroeseet al., 2021b)的效果。
目前,关于生物炭和有机肥如何影响人工林地木质分解的研究较少。Qu 等(2022)利用挪威云杉(Picea abies)木块在芬兰南部欧洲赤松(Pinus sylvestris)林地表面进行分解发现,生物炭改变了分解过程中细菌群落结构而对木质分解进程无影响。土壤温湿度是影响微生物定殖的重要因素,Qu 等(2022)的研究忽视了地下部分木质分解进程,以及单一树种的木质分解难以排除木栖真菌等微生物的偏好对分解的影响。利用标准规格的毛白杨、美洲山杨和火炬松木条在毛白杨人工林内进行了短期(6 个月)分解,Zhao 等(2022)发现,生物炭和有机肥促进了林地表面和土壤内木条的分解并改变了分解过程中的养分(氮、磷和钾)循环。但是,生物炭和有机肥是否会长期影响木质分解及其如何影响土壤养分循环相关酶活性尚未可知。考虑到生物炭和有机肥在长期碳固存、改善土壤健康和应对气候变化等方面的潜在价值(Yanget al., 2021),这些研究对制定短轮伐期经营人工林土地管理政策至关重要。
本研究利用3 个不同树种(毛白杨、美洲山杨和火炬松)标准规格木条在毛白杨人工林地内进行了近3 年的分解研究,并结合土壤养分循环相关酶活性(酸性磷酸酶、纤维素酶、α-葡萄糖苷酶、β-葡糖苷酶、N-乙酰-β-D-葡萄糖苷酶和芳基硫酸酯酶)变化,以期阐明生物炭和有机肥对森林土壤质量提升的作用。该研究将为杨树人工林林地可持续经营提供科学数据支撑和理论依据。基于前人研究,提出2 个假设:生物炭和有机肥能促进林地表面和土壤内木质分解;能提高土壤酶活性。
1 材料与方法
1.1 研究地概况
研究区位于山东省冠县国有毛白杨林场(36°31′21′ N, 115°21′37′′ E),海拔46 m,属于半干旱大陆性气候,研究期间月平均温度和降水量见(Zhaoet al., 2023)。该研究区域0~20 cm 土壤为碱性(pH8.2)砂壤土(砂土∶粉土∶粘土比例为62.3%∶35.0%∶2.7%),全碳、氮、磷和钾含量分别为11.00、2.23、 1.40 和8.34 mg·g-1(Zhaoet al., 2022)。
1.2 试验设计
本研究采用裂区试验设计,主区为有机肥,设3个水平 (M:0、3、9 t·hm-2)分别用M0、M3、M9 表示;副区为生物炭,设3 个水平(B:0、2.5、10 t·hm-2)分别用B0、B2.5、B10 表示;共计9 个土 壤处理(M0B0、M0B2.5、 M0B10、 M3B0、 M3B2.5、 M3B10、 M9B0、M9B2.5、M9B10),重复3 次。每个土壤处理为24 m ×24 m 正方形,相邻土壤处理之间设立8 m 宽缓冲带。在林地内将3 个树种(毛白杨、美洲山杨、火炬松)标准规格木条按照2 种方式(水平放置于林地地表,垂直插入0~20 cm 土壤内)放置在每个土壤处理中心,每个放置方式每树种木条放置5 根。
1.3 试验材料
本研究使用的猪粪有机肥(河北润东肥业有限公司),含水量为24%、pH 为7.9、全氮、磷、钾和有机质含量分别为14.64、19.52、26.84 和450.00 mg·g-1;玉米秸秆生物炭是在限氧、450~500 °C 条件下缓慢热解2 h 制得(勤丰众成生物质新材料(南京)有限公司),其pH 为8.8,碳和氢含量分别为47.56%和2.80%,全氮、磷和钾含量分别为10.70、1.60 和2.60 mg·g-1,碳氮比为44,比表面积为5.0 m2·g-1,空隙容量为6.9×10-3cm3·g-1,孔径为5.60 nm(Zhaoet al., 2022)。
毛白杨木条制作过程参考刘勇等(2015)。首先,在当地选取胸径约为40 cm,树高约30 m ,树干直挺,树结少的毛白杨去皮,利用其边材按照 2.5 cm × 2.5 cm ×40 cm (I 类)和2.5 cm × 2.5 cm × 50 cm (Ⅱ类)制作成木条。然后,将I 类木条裁成A 段2.5 cm × 2.5 cm × 15 cm、B 段 2.5 cm × 2.5 cm × 10 cm 和C 段2.5 cm × 2.5 cm ×15 cm 3 部分, 分别钉标签做标记。其中,B 段作为对照进行初始性质的测定,将所有A 段和C 段木条进行称重,作为表层分解材料。类似地,将 Ⅱ 类木条裁成D 段2.5 cm × 2.5 cm × 20 cm、 E 段2.5 cm × 2.5 cm ×10 cm 和F 段 2.5 cm × 2.5 cm × 20 cm 3 部分, 分别钉标签做标记。其中,E 段作为对照部分进行初始性质的测定,将所有D 段和F 段木条一端涂抹木材填孔剂来减少埋放后水分的流失,然后称重,作为土壤内分解材料。
美洲山杨和火炬松标准木条由美国农业部林务局落基山研究所提供,其制作方式、规格与毛白杨标准木条相同,详见Jurgensen 等(2006)。毛白杨、美洲山杨和火炬松木条的初始碳含量分别为46.75%、46.40%和48.62%; 全 氮 含 量 分 别 为1.40、 1.10 和0.80 mg·g-1;碳氮比分别为354.64 、 421.81 和 631.65;美洲山杨和火炬松木条的木质素含量分别为20.64%和31.09%,木质素∶氮分别为189.50 和403.72 (Wanget al., 2018; Zhaoet al., 2022)。
1.4 试验布设
2018 年4 月,造林地整地后,将有机肥和生物炭按照试验设计均匀撒施在林地表面,并利用旋耕机(1GQN- 200, 潍坊圣旋机械有限公司)翻入20 cm 土壤。随后,将1 年生毛白杨(三倍体 ‘北林雄株1 号’[(P.alba×P.glandulosa) × (P.tomentosa×P.bolleana)])苗木按照3 m × 4 m 株行距栽植。2019 年1 月,在M0B0、M0B10 和M9B10 处理10 cm 深处安装Onset Hobo 温度数据采集器和湿度传感器(onset computer corporation,Bourne,马萨诸塞州,美国)以2 h 的间隔监测土壤温湿度。
2018 年7 月,将每个树种木条按照30 cm 间隔放置在林地表面,每个树种5 根,并用不锈钢景观钉固定。在林地土壤内,为防止土壤压实,先用孔径为2.5 cm×2.5 cm 取芯工具以30 cm 间隔钻2.5 cm×2.5 cm×20 cm 孔,并将木条含填孔剂的一端朝上放入,与地面齐平 (刘勇等,2015)。本研究总计放置标准木条810根(3 个有机肥施用量 × 3 生物炭施用量 × 2 种木条放置方式 × 3 个树种 × 3 次重复 × 5 根/放置方式/树种)。
1.5 土壤和木条样品采集
2020 年7 月,使用直径为35 mm 的土钻在每个土壤处理随机选取6 个点收集0~20 cm 土壤,过2 mm筛去除根系等杂物后放置冰上运送至实验室进行细胞外酶分析;2020 年12 月(木条分解2.5 年后),将所有木条取出,移除附着物,记录编号,立即称取鲜质量并运送至实验室。
1.6 土壤酶活性、木条密度损失和含水量测定
土壤酸性磷酸酶、纤维素酶、α-葡萄糖苷酶、β-葡萄糖苷酶、N-乙酰基-β-D-葡萄糖苷酶和芳基硫酸酯酶活性采用微量法(关松荫, 1986)利用微量法试剂盒测定(北京索莱宝科技有限公司)。
在实验室内,将所有木条放置在恒温恒湿箱(20 ℃,65%相对湿度)中10 天后取出,量取其长、宽和高计算体积,然后在105 °C 条件下干燥至恒质量并称重。木条的密度损失通过初始木条密度(g·cm-3)减去分解2.5 年后木条密度来计算,以百分比表示(Shorohovaet al., 2008;Clineet al., 2017),木条含水量(%)为水分含量占木条干质量的百分比。
1.7 数据分析
使用R 4.1.1(Team 2021) 完成数据分析。采用单因素方差分析(ANOVA)比较了不同土壤处理(M0B0、M0B10 和M9B10)土壤温度和水分差异;以有机肥和生物炭为自变量,单个土壤酶活性作为因变量建立2因素线性混合效应模型(LME)研究生物炭和有机肥对土壤酶活性的作用。
对于木条,首先以有机肥、生物炭、木条放置方式和树种为自变量,木条密度损失和含水量分别为因变量建立4 因素LME 模型。因木条放置方式、树种及其二者交互作用为木条密度损失和含水量的主要方差来源(表1),故先采用Tukey 法对每个放置方式下不同树种木条间的密度损失和含水量进行事后检验。再将数据按照放置方式和树种分开,以有机肥、生物炭为自变量建立2 因素LME 模型研究生物炭和有机肥对不同放置方式下各树种木条分解和含水量的影响(表2)。
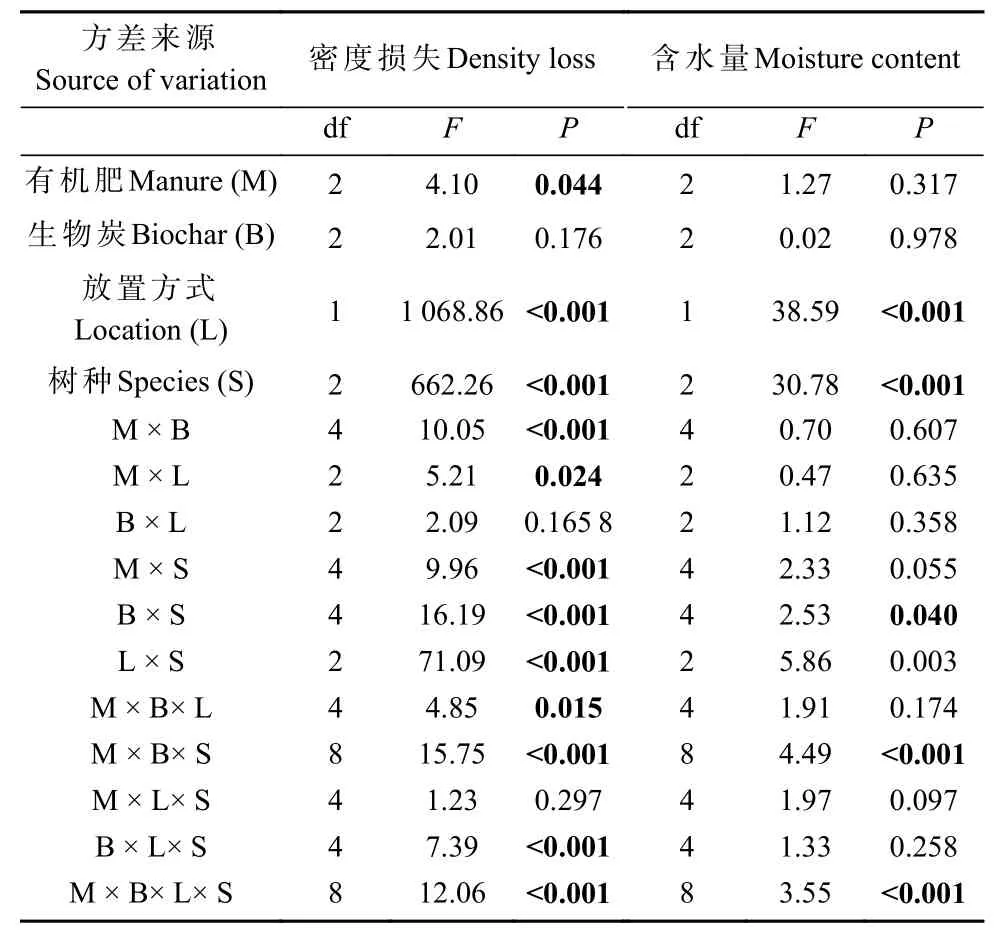
表1 木条分解2.5 年后密度损失和含水量变化4 因素方差分析①Tab.1 The four-factor ANOVA of wood stake density loss and moisture content after 2.5 years of decomposition
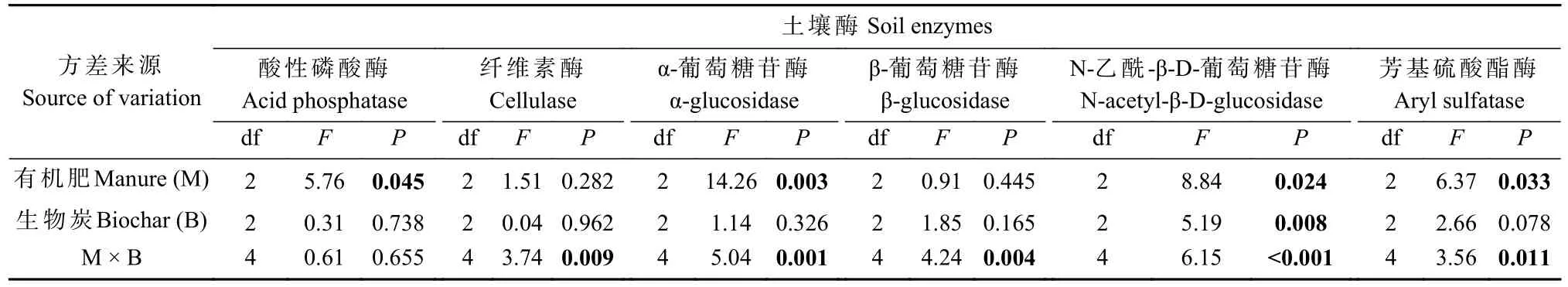
表2 有机肥、生物炭对土壤酶活性的主效应和交互作用①Tab.2 The main effects and interactions of manure, and biochar on soil enzyme activities
所有LME 模型均使用lmerTest 包(Kuznetsovaet al., 2017)完成,当给定因变量的F检验在P≤ 0.05水平下显著时,使用Emmeans 包(Lenthet al., 2022) 和Tukey-Kramerfa 法进行事后检验。所有2 因素LME模型中,当生物炭和有机肥存在交互作用时只对不同处理组合间进行比较,当仅受生物炭或有机肥主效应显著影响时,只进行主效应分析。另外,利用皮尔逊相关分析研究了不同放置方式下各树种木条密度损失与含水量之间的相关关系。采用Origin Pro 2022(OriginLab,马萨诸塞州,美国)进行绘图。
2 结果与分析
2.1 有机肥和生物炭对土壤条件的影响
2.1.1 土壤温湿度 因试验条件限制,本研究只对单一重复的未经处理土壤(M0B0)、高生物炭(M0B10)和高有机肥和生物炭(M9B10)处理10 cm 深处土壤温湿度进行了监测(图1)。研究期间,3 个土壤处理间土壤温度无显著差异(图1A)。然而,相较于M0B0,M0B10 和M9B10 处理有提高土壤含水量的趋势,该现象在雨季(6—8 月)尤为明显(图1B)。
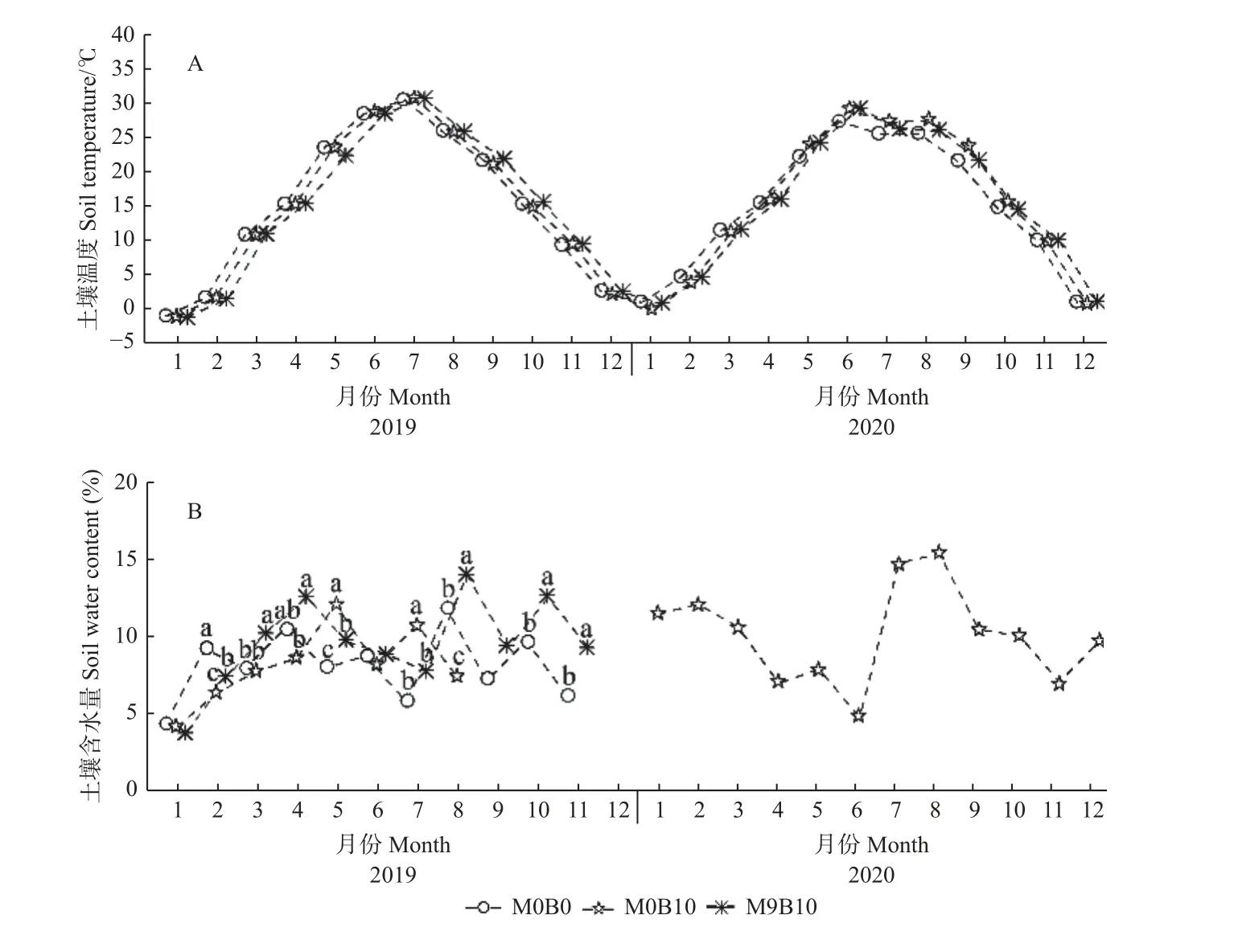
图1 不同有机肥和生物碳处理条件下土壤的月平均土壤温度(A)和含水量(B)Fig.1 Monthly average soil temperature (A) and water content (B) under different organic fertilizer and biochar treatments
2.1.2 土壤碳和氮循环相关酶活性 生物炭和有机肥处理2 年后,土壤酸性磷酸酶只受有机肥主效应的显著影响(表2)。其中,9 t·hm-2有机肥条件下土壤酸性磷酸酶活性较不施用有机肥提高了12.01%(图2A)。生物炭和有机肥对土壤纤维素酶、α-葡萄糖苷酶、β-葡萄糖苷酶、N-乙酰-β-D-葡萄糖苷酶和芳基硫酸酯酶活性存在显著交互作用(表2)。其中,与M0B0 相比,单独施用生物炭(M0B2.5 和M0B10)显著降低了土壤纤维素酶活性(图2B);然而,M3B10、M9B0 和M9B2.5 处理条件下土壤α-葡萄糖苷酶分别较M0B0处理提高了29.21%、68.54%和40.45%(图2C);单独施用生物炭或与有机肥搭配施用(M0B10、M3B2.5 和M9B2.5)提高了土壤β -葡萄糖苷酶活性,提高幅度分别为58.38%、76.76%和68.09%(图2D);对于N-乙酰-β-D-葡萄糖苷酶,低施用量的有机肥和生物炭(M3B2.5)及9 t·hm-2有 机 肥 处 理(M9B0、M9B2.5 和M9B10)条件下其活性较M0B0 处理提高了177.89%、198.03%、111.94%和92.79%(图2E);另外,所有生物炭和有机肥处理都提高了土壤芳基硫酸酯酶活性,提高幅度为62.84%~248.14%(图2F)。
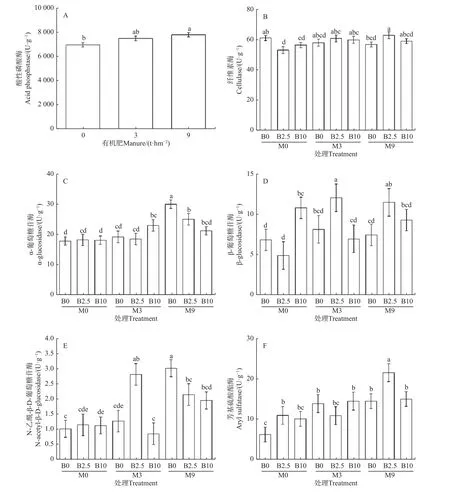
图2 生物炭和有机肥处理2 年后不同处理条件下的土壤酶活性Fig.2 Soil enzyme activities in different treatments after two years of soil biochar and manure amendments
2.2 有机肥和生物炭对木条分解(密度损失)和含水量的影响
2.2.1 木条密度损失 在林地表面分解2.5 年后,2 种杨树类木条分解较火炬松快,但毛白杨和美洲山杨间无显著差异(图3A)。方差分析结果表明,林地表面3种木条的密度损失均受有机肥和生物炭交互作用的影响(表3)。其中,与未经处理的M0B0 相比,单独施用 生 物 炭(M0B10)、施 用3 t·hm-2有 机 肥(M3B0、M3B2.5、M3B10)以及M9B2.5 处理显著促进了毛白杨木条的分解;另外,除M9B0 和M9B10 外,所有土壤处理显著提高了美洲山杨木条的分解,而M3B0 和M9B2.5 处理条件下火炬松木条的密度损失显著高于M0B0(图3C)。
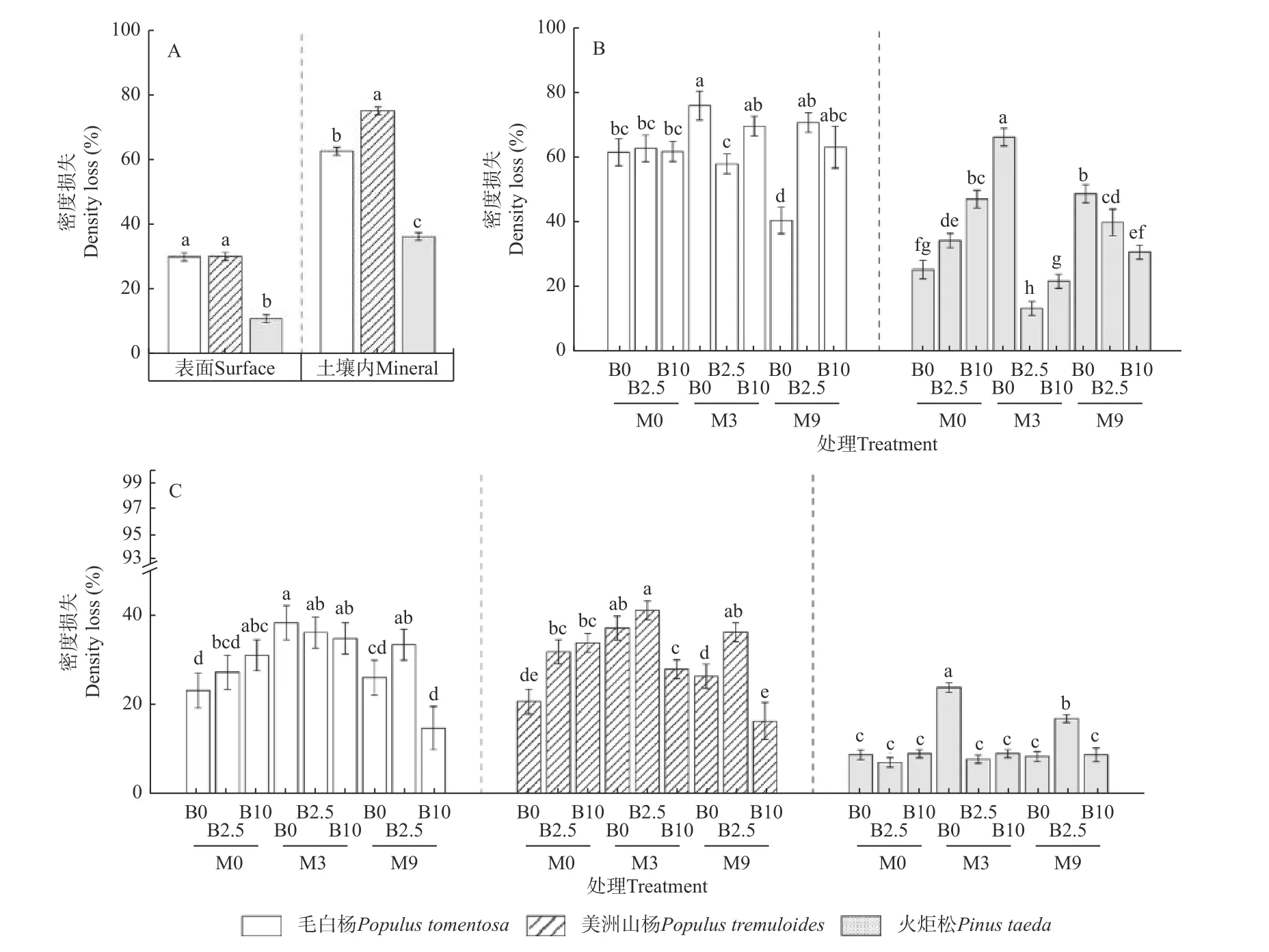
图3 表面和土壤内木条密度损失(A);不同土壤处理条件下土壤内(B)和表面(C)各树种木条密度损失Fig.3 The density loss of surface and mineral stakes with all soil treatments combined (A); mineral (B), and surface (C) stake density loss of each species under different soil treatments
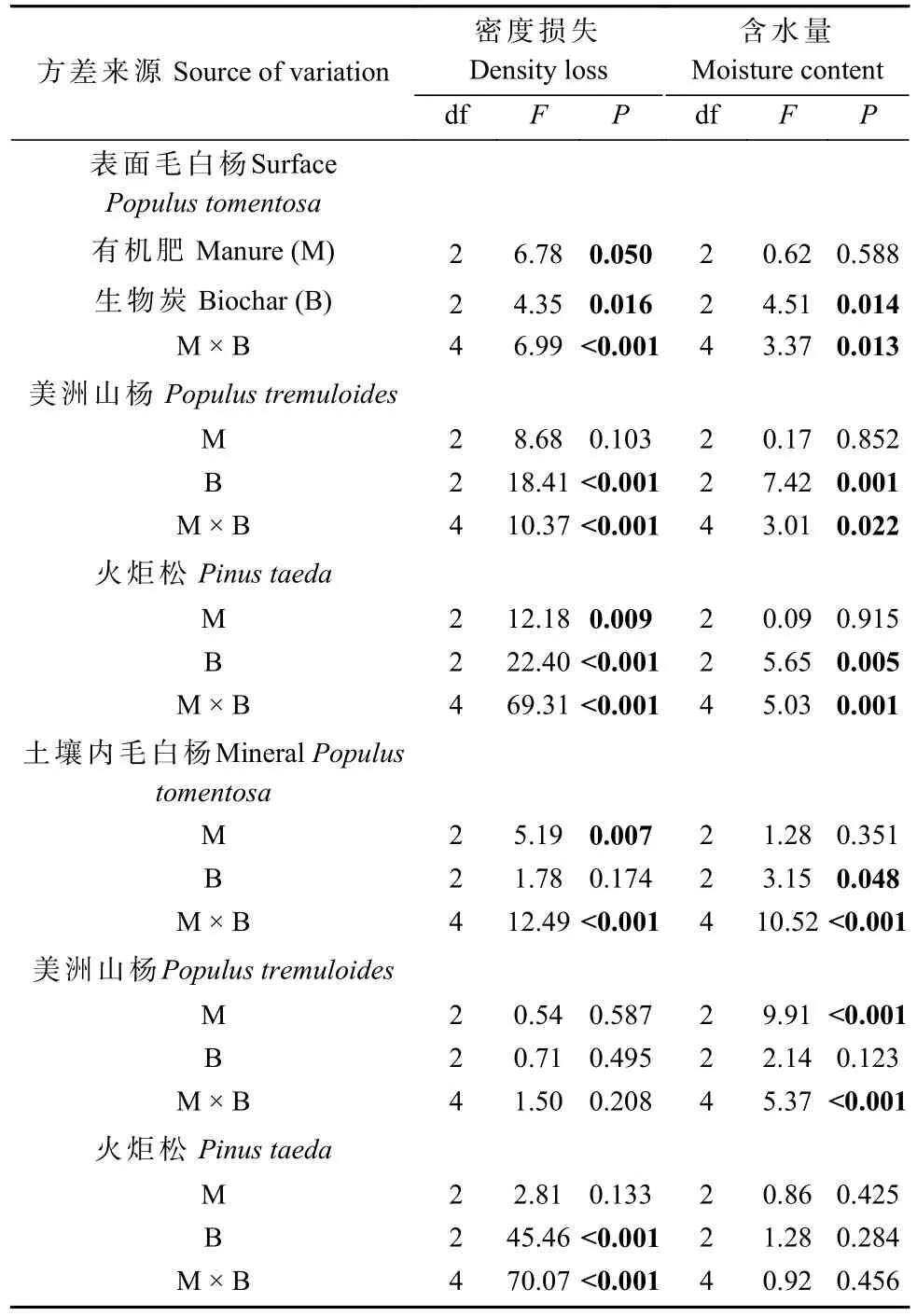
表3 有机肥、生物炭对不同放置方式、不同树种木条密度损失和含水量的主效应和交互作用①Tab.3 The main effects and interactions of manure and biochar on the density loss and moisture content of wood stakes of different locations and wood species
在林地土壤内,木条分解呈现美洲山杨>毛白杨>火炬松的顺序(图3A)。其中,有机肥和生物炭对毛白杨和火炬松木条密度损失存在显著交互作用,而美洲山杨木条的分解不受生物炭和有机肥处理影响(表3)。与M0B0 相比,M3B0 处理显著促进了毛白杨木条的分解,而M9B0 反之;另外,除M3B2.5、M3B10和 M9B10 外,所有土壤处理显著提高了火炬松木条的分解(图3B)。
2.2.2 木条含水量 林地表面木条含水量可以反映取样时林地地表的水分状况。本研究结果表明,不同树种间木条含水量差异显著,呈现火炬松>美洲山杨>毛白杨的顺序(图4A)。与密度损失类似,有机肥和生物炭对3 个树种木条含水量存在显著交互作用(表3)。其中,与未经处理的M0B0 相比,M3B2.5 处理显著提高了毛白杨和美洲山杨木条含水量;火炬松木条含水量在M3B2.5 处理条件下最高,但与M0B0 相比无显著差异(图4C)。皮尔逊相关性分析结果表明,表面毛白杨(r=0.32,P=0.001)和美洲山杨(r=0.43,P<0.001)木条含水量与密度损失存在显著正相关关系,而火炬松木条含水量与密度损失无显著相关性(r= -0.11,P>0.05)。
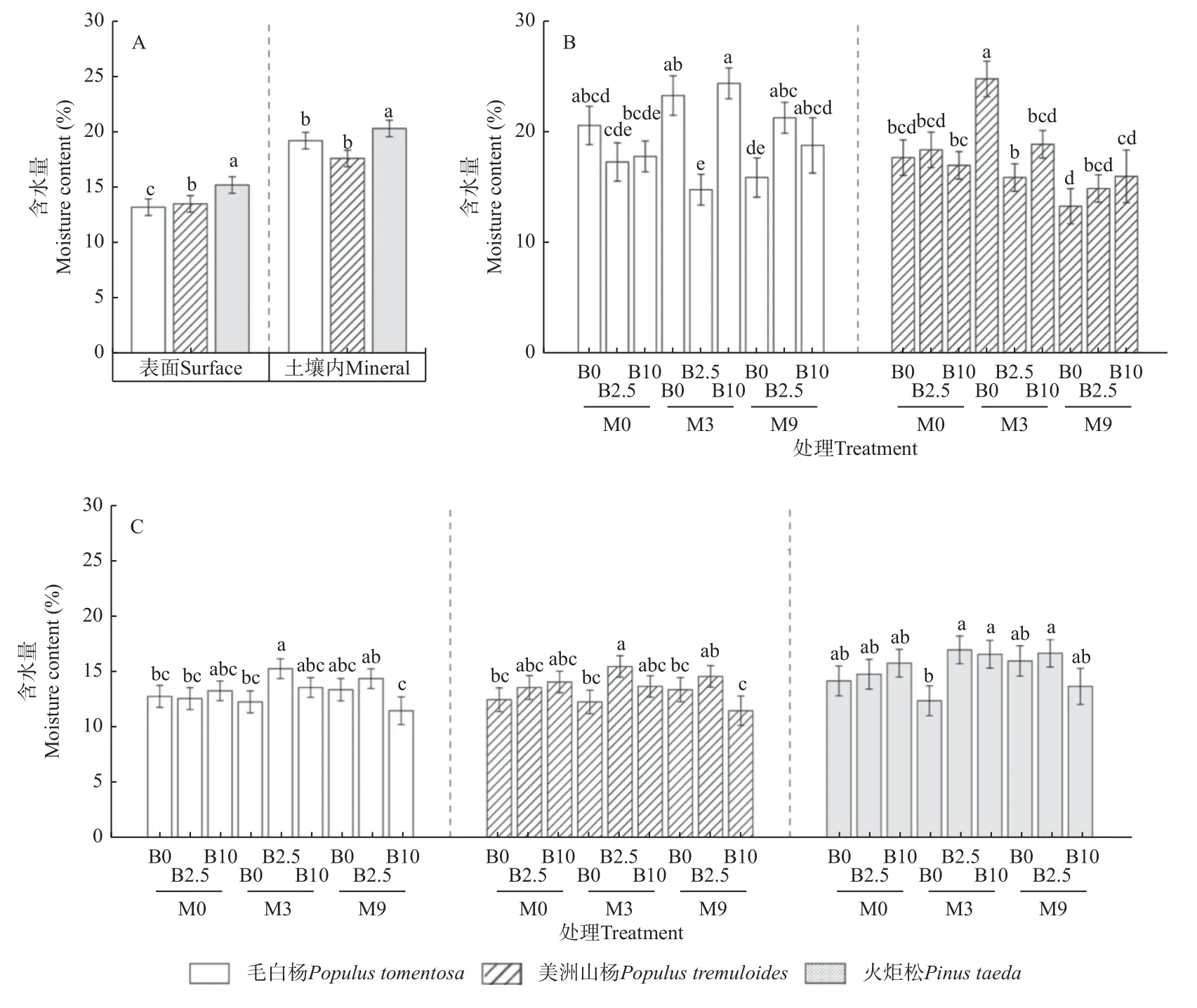
图4 表面和土壤内木条含水量(A);不同土壤处理条件下土壤内(B)和表层(C)木条含水量Fig.4 Surface and mineral stake moisture content under all soil treatments ( A ); mineral ( B ), and surface ( C ) stake moisture content under different soil treatments
在林地土壤内,木条含水量仍呈现火炬松>美洲山杨>毛白杨的顺序,但毛白杨和美洲山杨木条含水量无显著差异(图4A)。方差分析结果表明,毛白杨和美洲山杨木条含水量受土壤有机肥和生物炭显著交互作用影响,而火炬松木条含水量受土壤处理影响不显著(表3)。其中,相较于M0B0,M3B2.5 处理显著降低了毛白杨木条含水量;而M3B0 处理显著提高了美洲山杨木条含水量(图4B)。皮尔逊相关性分析结果表明,土壤内毛白杨木条含水量与密度损失呈显著 正 相 关(r=0.40,P<0.001),而 美 洲 山 杨(r=-0.22,P<0.05)和火炬松(r=-0.28,P<0.01)木条含水量与密度损失呈显著负相关。
3 讨论
生物炭和有机肥等土壤改良剂可以通过改善土壤物理、化学和生物学性质影响木质分解进程(Pingreeet al., 2022)。其中,微生物群落及其产生的胞外酶是许多土壤过程(有机质分解,养分循环等)的主要调节器,可反映生物炭等土壤添加剂对土壤养分循环过程的潜在长期影响(Elzobairet al., 2016)。本研究中,土壤处理2 年后,生物炭和有机肥显著提高了土壤养分循环相关酶(酸性磷酸酶、α-葡萄糖苷酶、β-葡萄糖苷酶、N-乙酰-β-D-葡萄糖苷酶和芳基硫酸酯酶)活性(图2),说明生物炭和有机肥具有长期提高人工林地土壤微生物活性的潜力。在林地土壤内,生物炭和有机肥处理促进了毛白杨和火炬松木条的分解(图3B)并提高了土壤含水量(图1)。与Zhang 等(2021)研究一致,生物炭等土壤添加剂可以通过改善土壤结构(尤其干旱、半干旱地区的粗质土壤)来提高土壤持水能力。本研究中,土壤处理显著提高了土壤内毛白杨和火炬松木条分解(图3B),但仅有M3B0 处理显著提高了火炬松木条含水量(图4B);同时,毛白杨木条密度损失和其含水量呈正相关,而美洲山杨和火炬松木条密度损失和含水量呈现负相关关系。以上结果结合土壤养分循环相关酶活性的提高(图2),可以推断本研究中土壤含水量可能不是制约土壤内木条分解的主要因素,生物炭和有机肥可能通过改变土壤微生物结构等加速了土壤内毛白杨和火炬松木条的分解。
在林地表面,生物炭和有机肥处理显著提高了3个树种木条分解(图3C)以及毛白杨和美洲山杨木条的含水量(图4C)。另外,结合林地表面毛白杨和美洲山杨木条密度损失和含水量的正相关关系(r分别为0.32 和0.43)和研究期间提高的土壤含水量(图1B),这表明本研究中生物炭和有机肥处理主要通过提高林地表面的含水量加速了木质分解进程(Adamset al.,2021; Page-Dumroeseet al., 2021b)。与前人(Lentzet al., 2019;Somervilleet al., 2020)的研究一致,生物炭和有机肥等土壤改良剂可以通过改变土壤的非生物特性来影响地下微生物的活性和随后的有机质分解。
在北半球,木条分解主要受当地气候条件下土壤温湿度的影响(Adamset al., 2021),土壤水分过低会限制真菌的定殖,但水分含量过高也会因为缺氧,限制微生物呼吸或抑制菌丝生长进而限制木质分解(Jaroszewiczet al., 2021)。在本研究中,土壤内木条较林地表面木条分解更快,这可能跟土壤内更适宜的温湿度有关(Wanget al., 2019; Bradfordet al., 2021)。另外,较大的日温湿度波动也可能是限制真菌在林地表面木条定殖的重要原因(Zhonget al., 2017; Wanget al., 2020)。此外,研究表明,落叶树种木质素含量、木质素∶氮和碳∶氮比值较针叶树种低,因此更容易受到腐生营养型真菌的定殖(Huet al., 2018;Perezet al.,2021)。本研究中木条分解表现为2 种杨树树种>火炬松。同时,杨树树种木条较高的孔隙率和较低的密度可能也是引起本研究中不同树种间分解差异的重要原因(Russellet al., 2015;Chaet al., 2017)。
4 结论
生物炭和有机肥可以用作改善土壤理化性质和微生物活性。本研究结果表明,生物炭和有机肥提高了土壤含水量和酶活性并促进了林地表面和土壤内木质分解。其中,林地表面木质分解的变化主要与含水量有关,而土壤处理可能通过提高微生物活动和酶活性加速了林地土壤内木质分解。标准规格的木质分解能灵敏反映土壤质量变化,然而,为了发挥生物炭和有机肥在提高林业土壤质量和缓解气候变化方面的作用并制定相应的林业土壤管理措施,需要进行更多的长期大田试验研究并阐明木条分解进程中其养分循环、真菌群落组成变化和土壤性质之间的关系。
猜你喜欢
学苑创造·B版(2022年2期)2022-01-28
新农民(2020年21期)2020-12-08
绿色科技(2019年19期)2019-11-26
数学小灵通(1-2年级)(2018年4期)2018-05-07
东坡赤壁诗词(2018年1期)2018-03-31
河南林业科技(2017年1期)2017-06-10
科学与财富(2017年10期)2017-05-09
广东农业科学(2016年7期)2016-10-08
黑龙江生态工程职业学院学报(2013年4期)2013-11-20
陕西林业科技(2010年2期)2010-08-15
