
不同生境中多树种生长对干旱胁迫的敏感性评价*
——以德国萨克森州为例
2023-12-11 03:00乌尔里希皮特扎卡安德烈亚斯罗洛夫张德顺
林业科学 2023年11期
刘 鸣 乌尔里希·皮特扎卡 安德烈亚斯·罗洛夫 张德顺
(1.南京工业大学 南京 211816;2.德国德累斯顿工业大学 塔兰特 D-01737;3.高密度人居环境生态与节能教育部重点实验室(同济大学) 上海 200092)
在气候变化影响下,欧洲中部的持续干旱严重影响树木生长,枝条和根部普遍发育滞缓,树木死亡率显著上升(Gillneret al., 2013a; Kunertet al., 2020;Laryschet al., 2021; Liet al., 2015)。为应对干旱,选择具有高抗旱性的树种来替代干旱敏感性树种,日益受到城市林业和园林绿化管理部门的关注(Blackmanet al., 2019; Eilmannet al., 2012; Gillneret al., 2014;Rahmatiet al., 2015)。如何简便、快速、有效地评价不同树种在多样化生境中的生长反应和敏感性,是亟待解决的实际科学问题。
以往关于树木生长的研究主要关注树木年轮的测量,即通过年轮宽窄对干旱的响应来判断该树种是否敏感(Gillneret al., 2014, 2013b; Nitschkeet al., 2017;Riglinget al., 2002)。但是,测量年轮需要从树干中抽取树芯,会对树木造成直接伤害,且病虫害的潜在侵入也会对树体产生间接性危害,故不容易获得城市林业管理部门的审批许可。生理生化试验(叶片)等检测方法也常用于树木生长活力的评估,通常需购置昂贵的仪器设备,实地操作也会因采样的差别造成检测结果的差异(王斌等, 2013; 董鹏等, 2018; 马斌等,2020)。生理生化试验指标一般只能反映当年树木状况,若需要评价树木对环境的长期动态变化就需要多年长期重复监测(Callowet al., 2018)。近年来,叶片膨压损失点水势(leaf water potential at turgor loss point,Ψtlp)作为相对简便的方法,被广泛用于检测各树种的生理与生态抗旱性(Bartlettet al., 2012a, 2012b;Blackman, 2018; Sjömanet al., 2018; Zhuet al., 2018),若Ψtlp为较小的负值,则表示该树木在干旱胁迫下具有较强的气孔和水分导度,能维持光合作用速率,具有较强抗旱性。但是,单一指标并不能完全反映树木在不同生境的抗旱能力(Farrellet al., 2017)。一些树木形态评估指标,如目视生命力指数(visual vitality index)、树冠透明度(crown transparency)、树冠形态(crown morphology)等,也可用于评估树木的生长活力,但这些方法主要为感性的经验估计,评价结果既不精确也不可靠(Callowet al., 2018; Dobbertin, 2005;Roloff, 1987)。
各树种有不同的干旱胁迫应对策略,如等水和非等水行为(isohydric and anisohydric behavior)、干旱逃避与干旱耐受策略(drought-avoiding and droughttolerating strategy)、水节约与水消费(water savers and water spenders)等(Litvaket al., 2012; Ogasaet al., 2014;Sadeet al., 2012; Weiet al., 2019; 罗丹丹等, 2017)。此外,各树种不同木质部结构也会影响其抗旱能力,在同等条件下特别是极端干旱条件下,散孔材树种因导水率低而比环孔材树种更敏感(Elliottet al., 2015;Hackeet al., 2006; Kunzet al., 2018; Tanedaet al., 2008;Vanhellemontet al., 2019)。因此,各树种抗旱机制十分复杂,其采取的抗旱策略也会随干旱程度和耐旱性差异而不同(Yiet al., 2017)。
树木枝条生长(shoot growth)是一种每年必发生的初级生长形式。与年轮相似,多年生树枝会形成节间(Cochardet al., 2005)。当新枝停止生长后,形成的节间长度(及各年的枝长)不会再变化(Roloff, 1987)。因此,通过测量树枝长度就可对树木生长进行精确评估,并且也可探知树木生长对环境的响应关系。目前,关于不同树种树枝生长对环境胁迫的响应的报道较少,其响应机制尚不清楚。
鉴于此,本研究通过测量不同生境中不同林龄的各树种年均枝长,评价各树种生长对干旱胁迫的敏感性,并通过对影响因子的分析,揭示不同树种对干旱胁迫生境的适应性反应特征。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于德国萨克森州中部地区(13°15′36″—13°30′05″E,50°59′11″—51°28′02″N),属 暖 温 带 气 候,年均气温8.5 ℃,年均降水量625 mm,7 月与8 月为最热月,年均最高温为23.6~23.7 ℃,7 月降水量最充沛,平均为82 mm(世界天气信息服务网,http://www.world weather.cn/)。地形地貌为平原和低山丘陵,土壤为砂质黄土。在研究区内共设样地14 个。其中12 个样地位于德累斯顿西北部的易北河河岸地带,均为开敞林和行道树,树木分布较为分散,全株几乎暴露于全光照之下。另2 个样地位于塔兰特(Tharandt)森林地带,为郁闭林,受高大乔木树冠遮蔽影响。在2020 年和2021 年秋冬季(10 月—次年3 月)2 次调查样地。总共测量了24 个不同树种-生境组合情况(表1)。按生境特点,将采样树木分为行道树(赋值=1)、开敞林树木(赋值=2)和郁闭林树木(赋值=3)。按树龄,分为幼龄树(5~8 年,赋值=1)、低龄树(9~15 年,赋值=2)和成年 树(16~30 年,赋 值=3)。参照木材数据库网站(InsideWood Database, https://insidewood.lib.ncsu.edu/)相关信息,各树种的木质部结构分为散孔材(diffuseporous,赋值=1)、半环孔材(semi-ring-porous,赋值=2)、环孔材(ring-porous,赋值=3)。
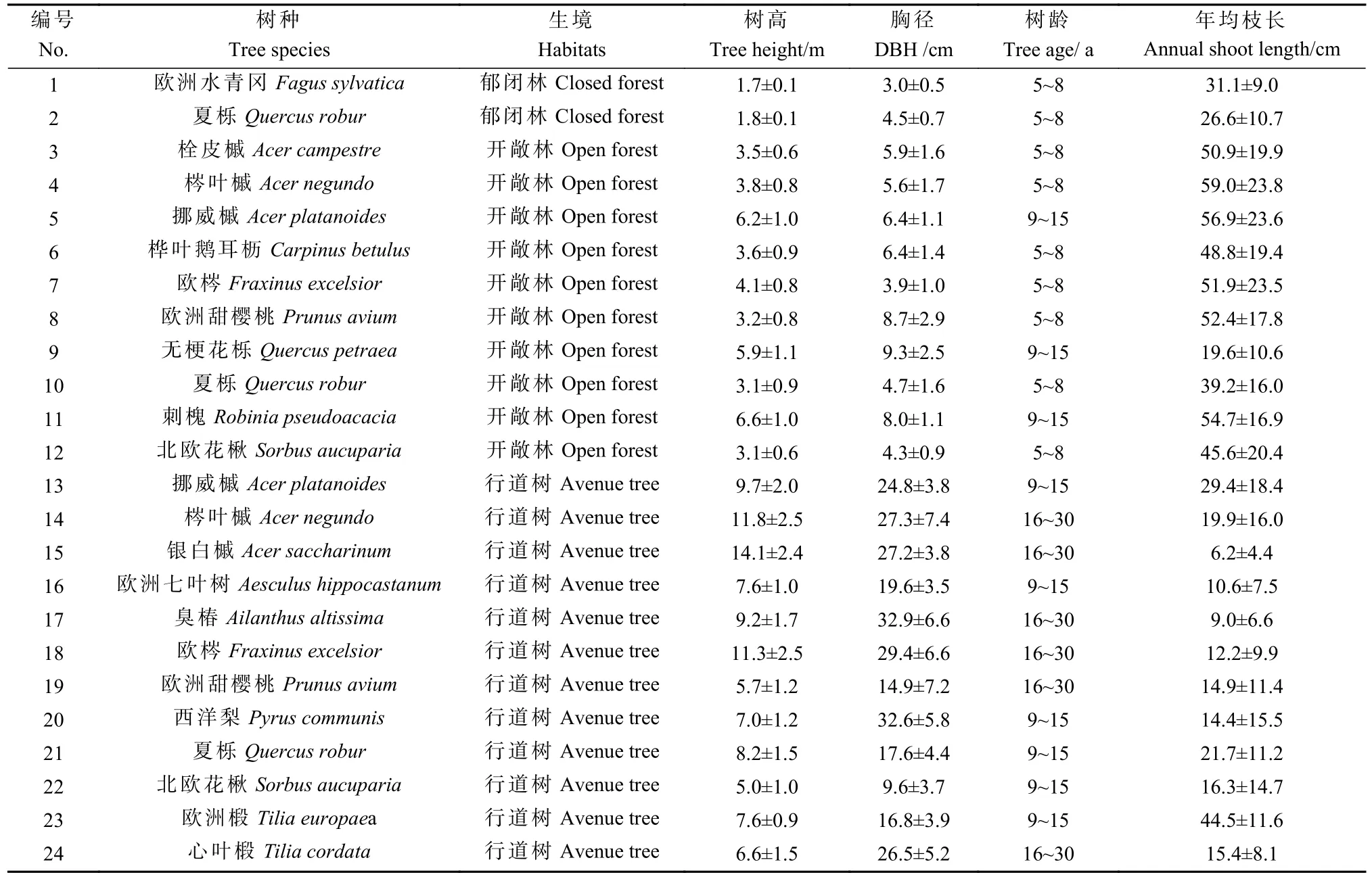
表1 调查采样的24 个树种–生境组合的有关信息Tab.1 Information of 24 surveyed combinations of tree species and habitats
1.2 标准化降水指数
本文采用标准化降水指数(standardized precipitation index,SPI)来指示当地气候干旱程度,并依此划分干旱与非干旱年份。SPI 是国际上广泛使用的衡量干旱程度的指数,其计算过程见参考文献(Brzosteket al., 2014; Caloieroet al., 2019; Łabȩdzki,2007; Nareshet al., 2009)。当SPI 低 于-1.0、-1.5 和-2.0 时,分别定义为中等干旱、严重干旱和极度干旱。获取德累斯顿克洛切(Dresden-Klotzche)气象站(51.1333 N; 13.7500 E)在1975—2020 年的各月降水数据,用于计算SPI(图1)。结果表明,2015—2017 年为非干旱年份,2018—2020 年为干旱年份。
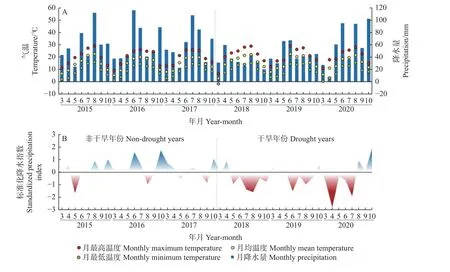
图1 在2015—2020 年生长季内(3—10 月)的各月气温、降水量(A)和标准化降水指数(B)Fig.1 The monthly air temperature, precipitation (A), and standardized precipitation index (B) during the growth season (March to October) in 2015—2020
1.3 各年年均枝长测量
本文采用年均枝长(annual shoot length,ASL)来评价各树种在干旱胁迫下的生长敏感性(Kusteret al.,2014; Roloff, 1987)。取每株树的任一主导枝,按枝节顺序依次测量节间距(首节长度为芽尖至第一枝节距离)作为各年枝长。每一树种不少于50 株样本,取其各年平均值作为该树种各年年均枝长。个别树种可能具有一年多次发枝的特性,例如栎树(Quercusspp.),则需要同时测量次轴方向上的侧枝长度来辅助鉴定(Kusteret al., 2014; Roloff, 1987)。本研究共统计了17个树种在24 个树种—生境组合条件下的1 200 个主导枝在6 年(2015—2020)间的各年年均枝长数据。为便于统计,将枝长长度分为5 个等级:极短(<5 cm)、短(5~10 cm)、中等(11~30 cm)、长(31~60 cm)、极长(>60 cm)。
1.4 生长敏感性评估
为评估各树种生长对干旱的敏感性,对各树种的各年平均枝长与当年(同年)及上一年的标准化降水指数进行Pearson 相关分析,其相关系数大小可指示对当年(同年)或上一年干旱的敏感性大小。通过系统聚类分析,可分为3 个生长敏感性梯度:不敏感(赋值=1)、中等敏感(赋值=2)、敏感(赋值=3)。
1.5 年均枝长相对增长率
为消除量纲和便于比较各树种生长应对干旱胁迫的响应大小,需对各树种的年均枝长进行标准化处理,计算年均枝长相对增长率(relative ASL increase rate,RAI),计算公式如下:
1.6 叶片膨压损失点水势反映的树种抗旱性
为探讨各树种的生长敏感性与抗旱性的关系,广泛收集已发表文献中多个树种在各种干旱胁迫情形下的Ψtlp数据(Bankset al., 2019; Bartlettet al., 2014;Fuchset al., 2021; Kunertet al., 2020; Leuschneret al.,2019; Löbbeet al., 2017; Marsalet al., 1997; Nardiniet al., 2012; Petruzzelliset al., 2019; Ranneyet al., 1990;Saviet al., 2016; Sjömanet al., 2015; Tsudaet al., 1997;Zhuet al., 2018)(表2),以此来确定各树种的抗旱能力大小。通过对各树种的Ψtlp平均值和极小值进行系统聚类分析,将各树种抗旱性分为4 类:低抗旱性(赋值=1)、中等抗旱性(赋值=2)、较高抗旱性(赋值=3)、高抗旱性(赋值=4)。
1.7 数据分析
本文所有数据分析均在SPSS 22.0 中进行。
2 结果与分析
2.1 年均枝长的种间差异
在郁闭林和开敞林中的幼龄树和低龄树的年均枝长呈中等(11~30 cm)和长(31~60 cm)的等级(图2),而成年行道树的年均枝长主要呈极短(<5 cm)和短(5~10 cm)的等级。开敞林中的无梗花栎(Quercus petraea)每年均生长有短枝。行道树欧洲椴(Tilia europaea)以长枝为主(图3)。大多数树种在干旱年份的短枝比例在增加,而长枝比例在下降。表现最差的树种是行道树西洋梨(Pyrus communis),其在干旱年份的极短枝比例逐年增加,在2020 年发现有56.0%的枝条已枯死。
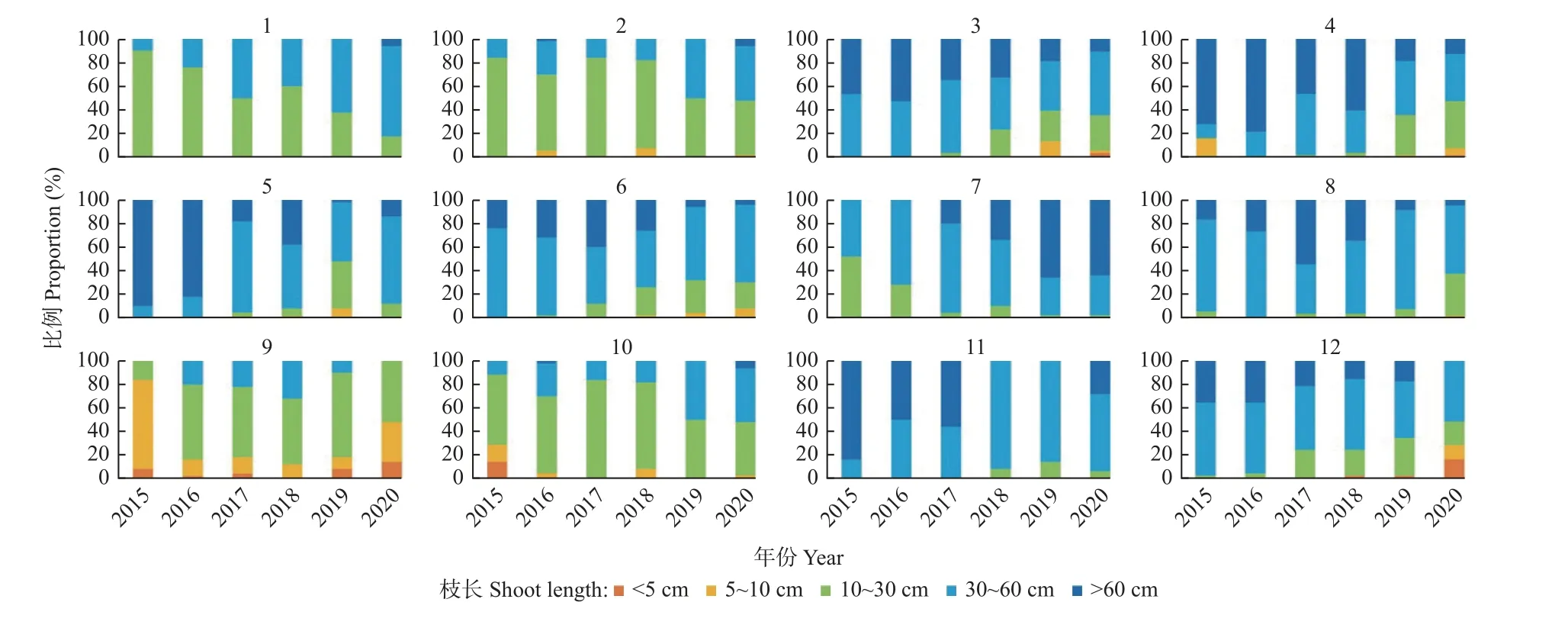
图2 郁闭林和开敞林中各树种的枝长分类Fig.2 Categories of annual shoot lengths of trees in closed and open forests
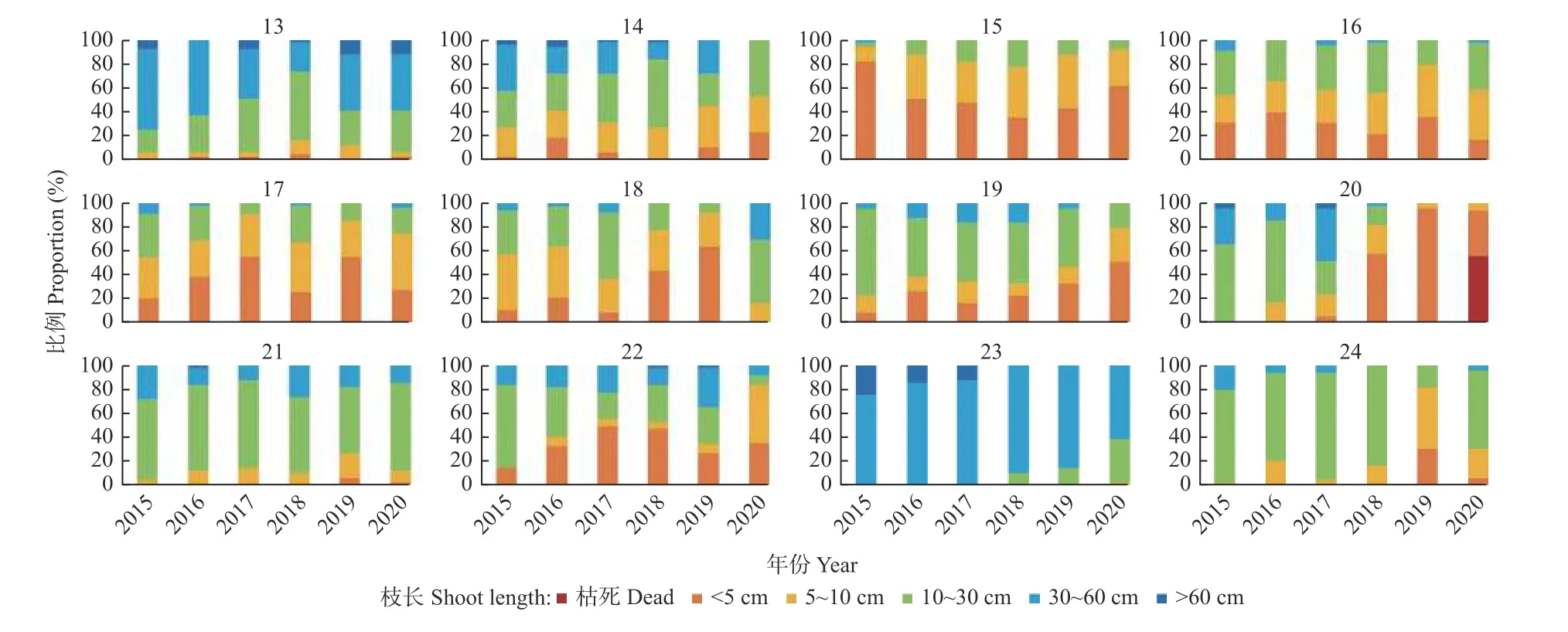
图3 行道树的枝长分类Fig.3 Categories of annual shoot lengths of avenue trees
2.2 年均枝长的年际动态变化
郁闭林中欧洲水青冈(Fagus sylvatica)和夏栎(Quercus robur)的年均枝长在干旱年份呈增长趋势(图4)。开敞林中,除欧梣(Fraxinus excelsior)和夏栎外,其他树种的年均枝长在干旱年份均表现为减小趋 势。 挪 威 槭(Acer platanoides)和 刺 槐(Robinia pseudoacacia)的年均枝长因受干旱影响而减小,但在2020 年的年均枝长却有增长趋势。绝大部分行道树的年均枝长变化较稳定,而行道树中挪威槭、欧梣和心叶椴(Tilia cordata)在2020 年的年均枝长有增加趋势(图5)。
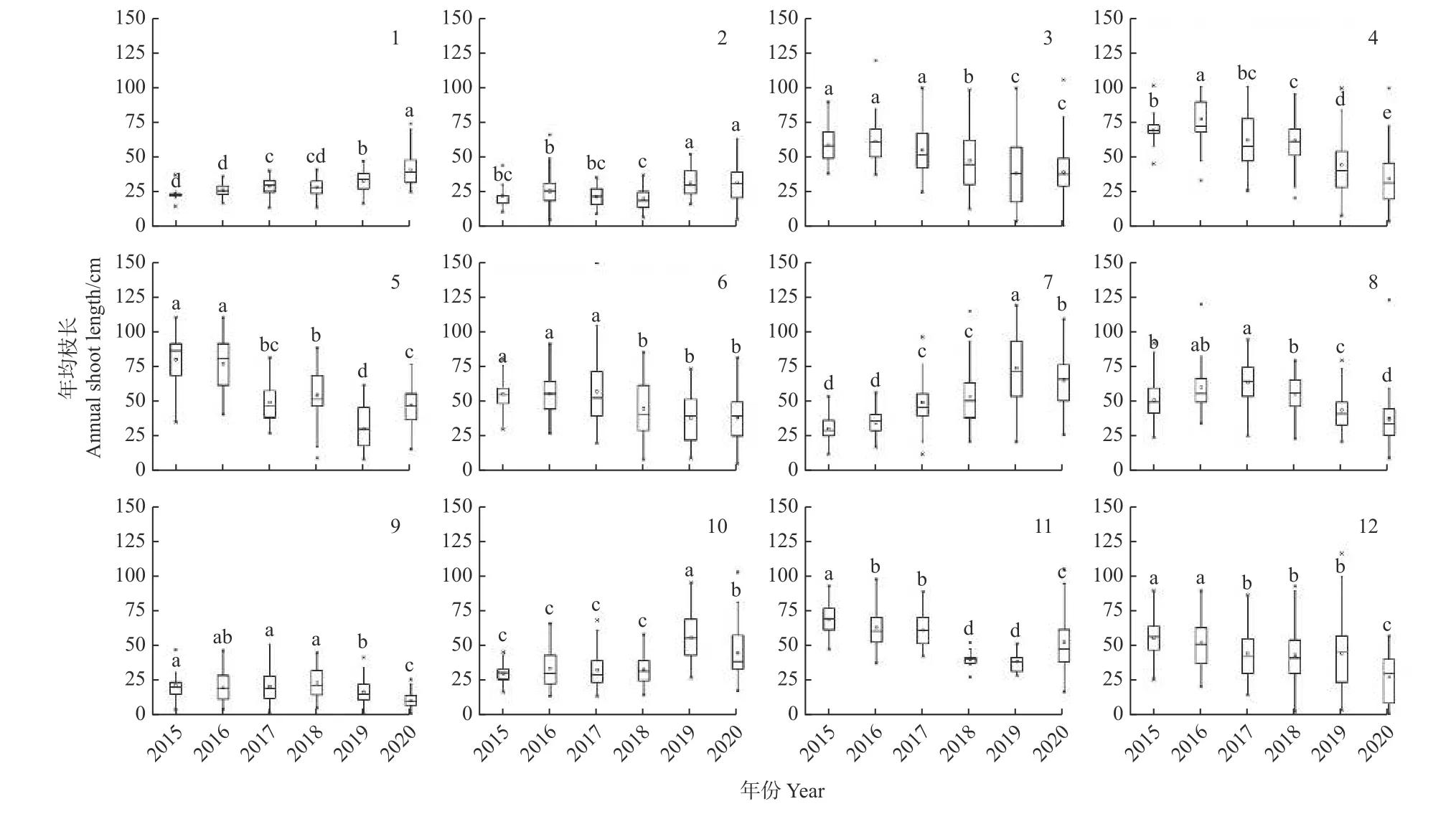
图4 郁闭林和开敞林中各树种的年均枝长变化Fig.4 Variation of annual shoot length of trees in closed and open forests
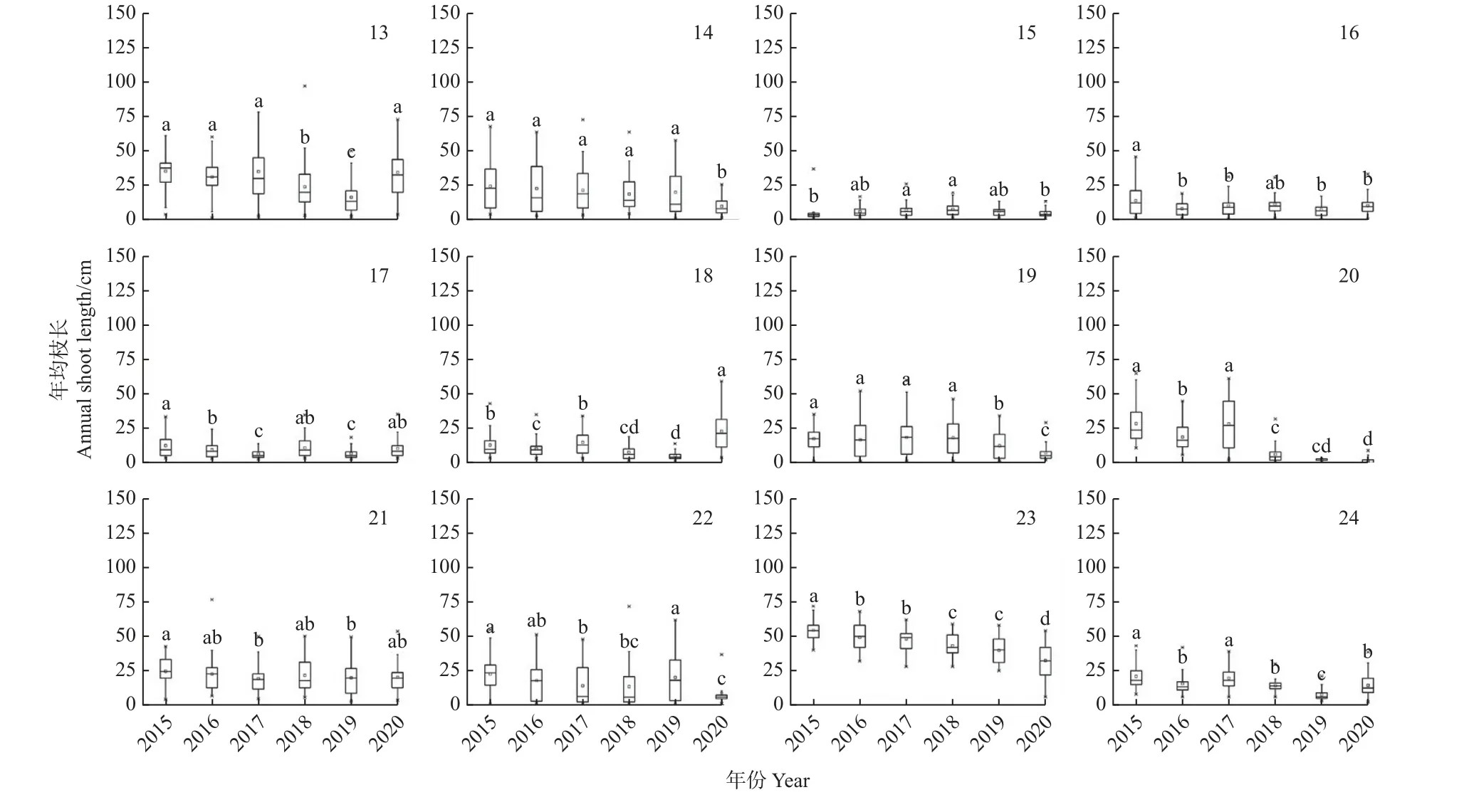
图5 行道树中各树种的年均枝长变化Fig.5 Variation of annual shoot length of avenue trees
2.3 各树种生长对干旱的敏感性
各树种的年均枝长与当年和上一年的SPI 都有较强的相关关系(r2=0.687,P<0.01)(图6)。郁闭林中的欧洲水青冈和夏栎,以及开敞林中的欧梣和夏栎的年均枝长与当年和上一年的SPI 均为负相关,反映出其对干旱胁迫不敏感。行道树中的银白槭(Acer saccharinum)、欧洲七叶树(Aesculus hippocastanum)、臭椿(Ailanthus altissima)、欧梣、夏栎和北欧花楸(Sorbus aucuparia)的年均枝长与SPI 呈中等相关关系,为中等敏感。其他各树种均呈较高的正相关系数,说明对干旱敏感。
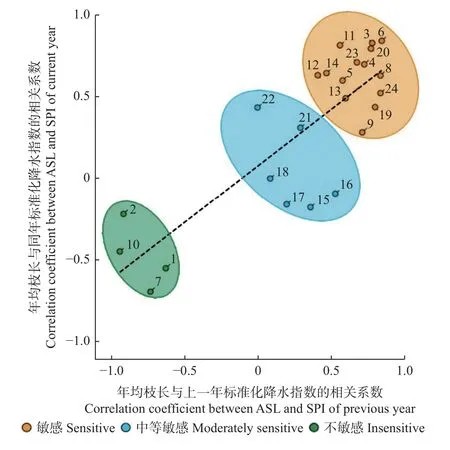
图6 各树种对干旱的生长敏感性Fig.6 Growth sensitivity of various tree species to drought stress
2.4 基于叶片膨压损失点水势的各树种抗旱性分类
各树种的Ψtlp的平均值与极小值具有正相关性(r2=0.549,P<0.01)(图7)。西洋梨和银白槭的Ψtlp最小,表明有很强抗旱性。栓皮槭(Acer campestre)、挪威槭、桦叶鹅耳枥(Carpinus betulus)、欧梣、无梗花栎和 夏 栎的Ψtlp平 均 值(-2.16~-2.73 MPa)和 极 小 值(-2.87~-3.09 MPa)均较低,表明具有较高抗旱性。而欧洲七叶树的Ψtlp平均值与极小值都最大,表明抗旱性最弱。
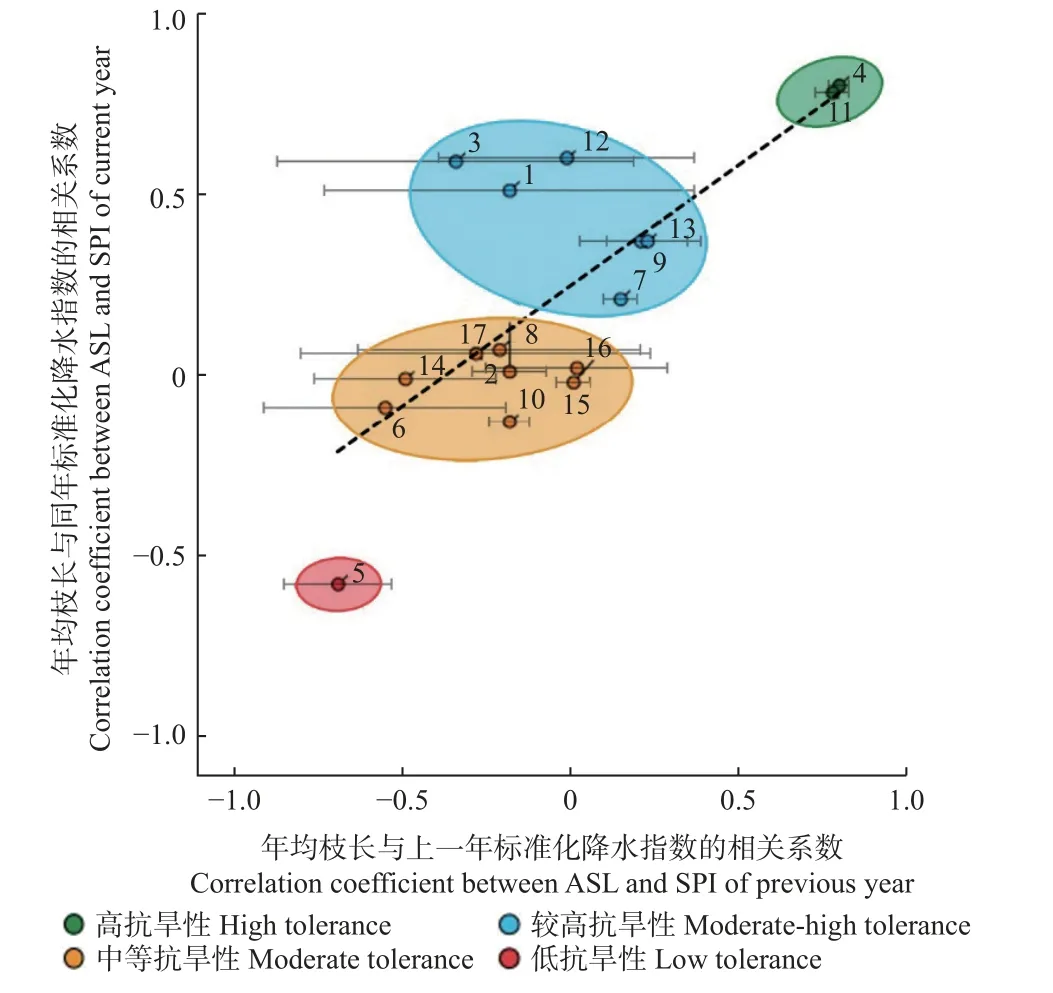
图7 各树种抗旱性分类Fig.7 Categories of drought tolerance of tree species
2.5 影响因子分析
利用各树种的RAI,可以更加清晰反映各树种年均枝长受干旱影响的差异(图8)。RAI 值越高,表明该树种枝条生长对干旱不敏感;相反,RAI 值越低,表明该树种对干旱胁迫越敏感。值得关注的是,各树种Ψtlp并不能反映在各种生境条件下的生长敏感性强弱。在不同生境下,郁闭林中的欧洲水青冈和夏栎的RAI均呈正值,但增长幅度却比开敞林中的欧梣和夏栎要低。除了行道树中的银白槭外,其他开敞林和行道树的各树种RAI 均为负值,表明受干旱胁迫影响强烈。其中,行道树中的西洋梨由于大量枝条死亡,其RAI最低。从树龄上看,幼龄树的RAI 均呈正值。除成年银白槭外,其他低龄树和成年树的RAI 均呈负值。从木质部结构上分析,在环孔材树种中除刺槐外的其他树种均表现出较高的RAI,而半环孔材树种的RAI 比散孔材树种高。
相关分析表明,RAI 与生长敏感性呈负相关关系(r=-0.853,P<0.01),说明RAI 能有效反映生长敏感性,RAI 越大时树木枝条生长对干旱的敏感性越小(表3)。按偏相关分析结果,其他因子对其相关性影响微弱(表4)。各树种的木质部结构与其RAI(r=0.553,P<0.01)和生长敏感性(r=-0.545,P<0.01)均呈显著相关关系,偏相关结果也显示,木质部结构对RAI 和生长敏感性均有显著影响。生境与树龄之间也有较高相关性(r=-0.771,P<0.01),但偏相关结果则表明,其他因子对树龄与生境之间的相关性没有影响。

表3 因子间Pearson 相关性分析①Tab.3 Pearson correlation analysis of factors

表4 因子间偏相关分析①Tab.4 Partial correlation analysis of factors
3 讨论
3.1 生物与非生物因子对年均枝长的影响
在干旱胁迫下,树木生长受生境条件和树种本身抗旱能力的共同影响(Castagneriet al., 2021; Gillneret al., 2015, 2014, 2013b; Pichleret al., 2007)。在生境方面,郁闭林下的欧洲水青冈和夏栎幼龄树的枝长受干旱直接影响程度较小,可能是因其所处林下小气候环境较为湿润,但可能由于光照不足,其枝长比开敞林中的欧梣和夏栎短。开敞林树木和行道树暴露于强烈太阳辐射之下,易受干旱的直接影响。相比而言,开敞林的生境条件比行道树要更加接近理想的自然生境条件,但是,在严重干旱胁迫下,当土壤中水分和养分资源不足时,开敞林中树木之间的竞争也会影响其年均枝长(Castagneriet al., 2021; Coomeset al., 2007;Dobbertin, 2005; Mradet al., 2019)。各树种抗旱性不同,年均枝长的反应也不同。
树龄是另一个重要影响因子(Eilmannet al., 2012;Pretzsch, 2020; Roloff, 1987)。各树种的生长遵循各自的树龄生长曲线(growth curve),在不同树龄阶段的生长过程和生长速率各有特点。幼树的枝条生长潜力旺盛,年均枝长较长。当进入成年和高龄阶段后,尤其是达到或超过生长极限点(culmination point)后,树木年均枝长变短,生长活力便会逐渐下降。本文结果表明,幼龄树和低龄树的年均枝长较长,而成年树的年均枝长普遍较短。树龄较大的行道树易受恶劣环境影响,例如有限的生长空间、土壤紧实度过高、土壤养分不足、空气污染以及冬季除冰盐使用等,都会额外加重对其正常生长的干扰,导致其年均枝长减小(Gillneret al., 2017, 2014; Sæbøet al., 2003; Sjömanet al., 2015)。在调查中,检测了行道树银白槭近15 年间(2006—2020)和欧洲七叶树近10 年间(2010—2020)的年均枝长变化,发现其生长势均呈下降趋势。故而认为,严重干旱以及长期恶劣的人工生境共同导致其“未老先衰”的结果。
此外,在严重干旱影响下,嫁接效应可能是导致行道树生长活力更低、年均枝长更短的原因之一。很多行道树是嫁接苗,例如,梣叶槭(Acer negundo)、银白槭、西洋梨、欧洲甜樱桃(Prunus avium)和欧洲椴等。嫁接苗在树干上能发现明显的瘤状嫁接斑痕,阻碍了导管中的水分和筛管中养分的上下传输(Gonçalveset al., 2007)。
3.2 年均枝长变化与种间抗旱性差异的关系
通过测量年均枝长,不仅可反映各树种在不同生境下的生长状况,也可以指示各树种自身抗旱性的强弱。例如,银白槭、欧梣和夏栎通常被认为具有较高抗旱性(Gillneret al., 2014; Kunzet al., 2018; Sjömanet al., 2015)。本文结果也证实行道树银白槭的年均枝长相对较长,不同生境下的欧梣和夏栎的年均枝长较长。相反,年均枝长表现较差的树种的抗旱能力也相对较弱。
本研究未发现Ψtlp与RAI 具有相关性。一方面,Ψtlp虽能反映叶片水平上的气孔导度调节和水分分配策略,但不能反映树木在逆境胁迫下的最终生长结果。另一方面,Ψtlp在指示不同生境上也有一定的局限性。在极干旱环境下,树木往往会被迫落叶以减少水分蒸腾。而年均枝长作为最终的生长结果,却可以反映树木对不同生境的综合抗逆反应。建议应慎重将Ψtlp作为判断树种抗逆性的单一指标,应重视测量年均枝长对树木抗逆性的初步评价。
各树种的木质部结构与其抗旱能力差异有关。例如,欧梣、夏栎都是环孔材树种,木质部中有相对较长和较宽的导管,能更有效地输送水分以应对干旱胁迫,因而表现出较强抗旱能力(Hackeet al., 2006;Leuschneret al., 2019; Liet al., 2015; Litvaket al., 2012;Percivalet al., 2006; Tanedaet al., 2008; Yinet al., 2017;Zimmermannet al., 2021; 李泽东等, 2021)。本文结果也表明,环孔材的年均枝长较长,半环孔材次之,散孔材最次。需特别注意的是,木质部结构与树种抗旱性的机制较为复杂。例如,刺槐和无梗花栎也是环孔材树种,但其年均枝长较短,原因可能与严酷干燥的生境条件有关,即极端土壤干旱可能已超越了刺槐和无梗花栎的抗旱生理极限(Boseet al., 2021; Cochardet al., 1992)。在调查中发现,刺槐种植在沙地上,无梗花栎种植在垃圾土上,缺乏养分和土壤贫瘠可能是造成其生长活力低下的原因之一。
虽然,近些年的研究表明,气候变化导致的干旱是造成各树种生长活力下降的主导因素,但其他因子也会对树木生长活力和年均枝长产生影响,包括高温、霜冻、风袭、虫害和病害等 (Bankset al., 2019; Ordóñezet al., 2015; 2014)。尽管通过测量各树种的年均枝长可反映树种的生长敏感性和抗旱能力,但仍需进一步探索其深层次的响应机制(Boseet al., 2021; Kusteret al., 2014)。
4 结论
通过测量和分析17 个树种在不同生境和干旱胁迫程度下的多年年均枝长,发现各树种生长活力与生境条件以及自身抗旱性紧密相关,受多因子的影响和制约。年均枝长可作为一种简便、快速、有效评价树木生长活力和生境适应的方法,可为城市林业和园林绿化部门初步筛选应对气候变化的抗逆树种提供科学依据。
猜你喜欢
公民与法治(2022年11期)2022-12-06
榆林学院学报(2022年4期)2022-08-02
家教世界·创新阅读(2021年12期)2021-01-13
计算机与生活(2018年8期)2018-08-15
现代园艺(2017年22期)2018-01-19
浙江农业学报(2017年1期)2017-05-17
甘肃林业(2016年2期)2016-11-07
西南农业学报(2016年4期)2016-05-17
理科考试研究·高中(2016年9期)2016-05-14
山西农经(2015年7期)2015-07-10
