
核磁共振技术在沼生栎种子失水过程中水分相态变化*
2023-12-11 03:00朱铭玮解志军张中会李淑娴
林业科学 2023年11期
袁 鸣 朱铭玮 解志军 康 真 张中会 李淑娴
(1.南京林业大学南方现代林业协同创新中心 南京 210037;2.南京市六合区人民政府横梁街道农业服务中心 南京 211515;3.襄阳市林业科学技术推广站 襄阳 441022;4.苏州纽迈分析仪器股份有限公司 苏州 215163)
核磁共振(nuclear magnetic resonance,NMR)技术作为一种快速、无损的新型测定技术,能通过测得样品氢质子密度与分布来反映样品中的水分相态含量及迁移情况(谢小雷等,2014;宋平等,2016;Xiaoet al.,2017)。近年来,核磁共振波谱(nuclear magnetic resonance spectroscopy)技术已广泛应用于食品、生物学等领域(石芳等,2017)。宋珊珊等(2020)利用核磁共振技术发现板栗(Castanea mollissima)种子贮藏期间,不易流动水可以转化为结合水、自由水以稳定其内部结构。汪楠等(2017)利用核磁共振技术对稻谷(Olyza Sativa)的干燥特性与水分迁移规律进行研究,结果表明,干燥过程中3 种状态水分的核磁反演图谱向左迁移,结合水和束缚水的峰面积显著减少,但自由水的峰面积变化较小。
沼生栎(Quercus palustris)是壳斗科(Fagaceae)栎属(Quercusspp.)落叶乔木,原产美国,广泛分布在北半球温带区域(段丽君等,2020)。沼生栎树形优美,叶缘有齿裂,新叶亮红色,熟叶深绿色,秋季叶色逐渐转变为橙红色或铜红色(格瑞,2004)。沼生栎喜光照,抗风性强,抗霜冻,能适应城市环境污染,在我国北京、泰安等地区均有引种栽培,是良好的城市园林、工业区绿化树种(李云龙,2001;黄利斌,2007)。沼生栎以种子繁殖为主,但该种子对失水较敏感,属顽拗性种子,这给其运输、贮藏带来诸多不便(Dickieet al.,2002;Xiaet al.,2012a)。水分流失的高度敏感是影响顽拗性种子贮藏和长期保存的主要因素,失水过程中水分相态的变化及各相态的水分在脱水过程中的作用还鲜有研究。鉴于此,本文拟利用核磁共振技术探究沼生栎种子失水过程中种子内部水分相态及含量的变化,同时测定失水过程中种子发芽率的变化,探明其半致死和致死含水量,为沼生栎种子的运输、贮藏提供指导,也为其他顽拗性种子的研究提供参考。
1 材料与方法
1.1 试验材料
沼生栎种子于2018 年秋季从美国北卡罗来纳州达勒姆进口。2019 年春天购于上海瑶琪园林有限公司,水选去除虫害粒,选取无机械损伤的种子用于试验。
1.2 试验方法
1.2.1 新鲜种子含水量测定及试验材料处理 随机数取新购(对照)沼生栎种子5 粒,重复3 次,将每粒种子切成4 部分,用千分之一天平称重,按照ISTA(2020)的要求,采用低恒温烘干法测定新购种子的含水量。
种子处理:随机数取一定数量的新购种子,称重后将种子置于实验室台面上自然风干,每天对种子进行称重并计算其含水量。当种子含水量降至30.0%、25.0%、20.0%、15.0%、10.0%时,取一定数量各失水阶段的种子用于各试验,以新购买的种子作对照。
1.2.2 种子发芽率测定 沼生栎种子具有浅休眠特性,因此发芽率测定前需先将1.2.1 中处于不同含水量阶段的种子放在4 ℃的冷库中,湿沙层积15 天解除休眠。层积结束后,随机数取不同含水量的种子25 粒,3 次重复,将种子置于沙床中,在25 ℃的光照培养箱中进行发芽试验。3 天后记录发芽种子的粒数,每2 天统计1 次,胚根长度达到0.5 cm 时即认为种子已萌发。按照ISTA(2020)的规定,连续3 天发芽粒数不超过1%时即认为发芽结束。发芽结束后,计算对照及各处理种子的发芽率。
1.2.3 种子半致死含水量的确定 根据试验结果绘制含水量—发芽率的关系曲线,再利用Excel 拟合,得出符合该曲线的回归方程(李佳琦等,2021),计算沼生栎种子的半致死含水量。
1.2.4 种子失水过程中各水分相态的测定 选取外表完好的沼生栎种子10 粒,3 次重复,将其置于玻璃试管中,放置在低场核磁共振仪(苏州纽迈电子科技有限公司,PQ001,0.5 T;射频脉冲频率:12.2 MHz;探头线圈直径:25 mm)中进行测定。测定时,采用CPMG(Carr-Purcell-Meiboom-Gill)脉 冲 序 列,并 利 用SIRT(联合迭代重建技术)算法反演操作,得到T2弛豫图谱。CPMG 脉冲序列的主要参数:主频SF=21 MHz,偏移频率O1=239 082.32 Hz,90°脉冲射频脉宽P1=5.20 μs,180°脉冲射频脉宽P2=10.80 μs,信号采样点数TD=79 998,重复采样等待时间TW=2 000 ms,重复采样次数NS=16,回波时间TE=0.1 ms,回波个数NECH=4 000。随后将种子置于实验台面自然风干,每天称重并计算其含水量。当含水量达到预定值时,再将种子置于核磁共振仪中重复上面的操作,直至种子含水量降为10.0%为止。
1.2.5T2弛豫谱峰面积与沼生栎种子含水量关系的构建 NMRT2弛豫谱总峰面积与样品中氢质子的数量成正比,故可以用T2弛豫谱总峰面积反映种子中水分的含量。参照宋平等(2015)的方法,将传统的质量信号换做核磁信号量,再通过回归分析,确定T2弛豫谱总峰面积与种子含水量的回归方程,通过R2检验对回归方程的拟合效果进行校验。
2 结果与分析
2.1 失水对沼生栎种子发芽率的影响
失水过程中,沼生栎种子发芽率的变化见表1。沼生栎种子初始含水量为38.0%,发芽率为96.00%。随着种子的不断失水,发芽率呈下降趋势。当含水量下降至30.0%时,种子的发芽率较CK 下降6.95%,但二者差异不显著(P<0.05);含水量降至25.0%时,发芽率显著下降至81.33%,较CK 下降15.28%;含水量下降至20.0%时,发芽率下降至74.67%,较CK 下降22.22%,种子的外部形态也发生明显变化,即种皮变硬,颜色变浅,切开后发现子叶与种皮已明显分离;含水量下降至10.0%时,种子完全丧失萌发能力,发芽率降为0。上述结果表明,沼生栎种子不耐脱水,属于顽拗性种子,种子完全致死的含水量为10.0%左右。
2.2 沼生栎种子含水量与发芽率之间的拟合曲线
分别采用多种模式对失水过程中沼生栎种子含水量与发芽率之间的关系进行拟合,发现采用“多项式”拟合模式,其R2最接近1。栎种子含水量与发芽率的拟合曲线见图1,关系方程为y= -0.170 7x2+11.601x-100.15,R2= 0.971 5,其中y为沼生栎种子发芽率,x为种子含水量。根据此方程计算发芽率下降至50.0%,即半致死状态时,种子含水量为17.39%,远高于正常性种子的安全含水量(10.0%),说明沼生栎种子的耐脱水性较差。
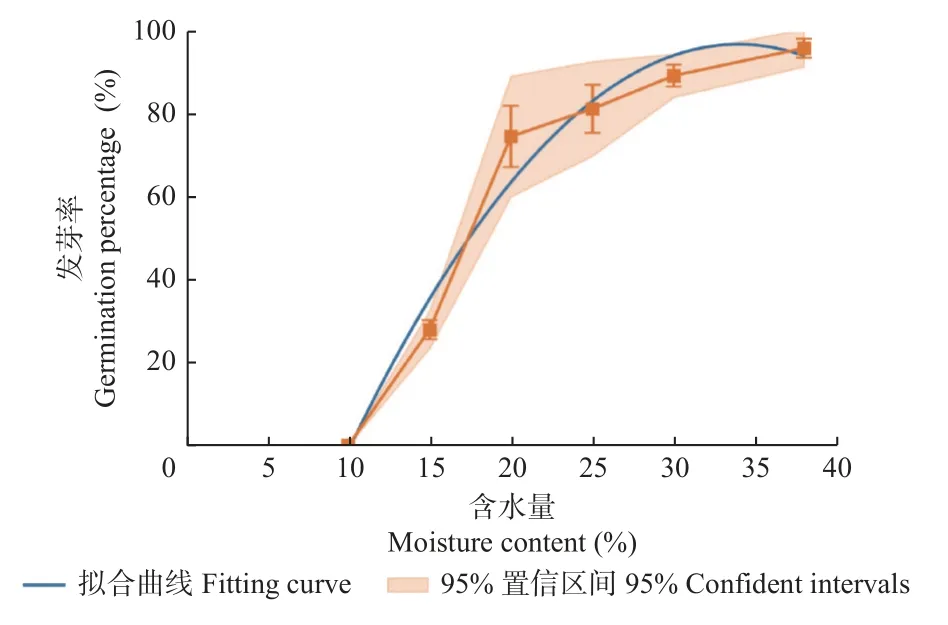
图1 沼生栎种子发芽率与含水量的拟合曲线Fig.1 Fitting curve of moisture content and germination percentage of Quercus palustris seeds
2.3 失水过程中T2 弛豫时间与水分相态的划分
根据核磁技术原理,横向弛豫时间T2的长短反映了样品中水分的自由程度,因此根据T2弛豫谱中波峰位置的差异可以区分样品中水分的相态。弛豫时间越短,水分的自由度越低,在T2谱上峰位置靠左,反之,弛豫时间越长,水分自由度越高, 在T2谱上峰位置靠右。由图2 可知,沼生栎种子失水过程中T2的变化范围主要在0.01~1 000 ms。横向弛豫时间T2图谱有3个明显的波峰,从左向右各峰值的T2弛豫时间分别为T21、T22、T23。其中弛豫时间较短的T21为束缚水(bound water),该状态的水主要被淀粉、蛋白质等大分子包围或是在细胞内通过氢键被大分子物质吸附,流动性很差(宋平等,2015);弛豫时间较长的T22为不易流动水(immobile water),其流动性虽然受到一定限制,但略高于束缚水;自由水(free water)T23弛豫时间最长,该相态水为游离状态,在细胞内可自由流动,不会被生物大分子吸附,是良好的溶剂,能够参与细胞的物质代谢。
中外运-敦豪国际航空快件有限公司近日在其北京总部举行媒体发布会,宣布其珠海口岸正式落成并投入使用,成为落户珠海口岸国际快递监管中心的首家国际快递公司。此外,DHL正式宣布将持续加大在中国的战略投资,对外公开了今年以来的一系列投资举措。DHL称,借助港珠澳大桥带来的高效物流通道,DHL珠海口岸的建立将大幅提升珠江西岸地区国际物流的快递效率。随着2018年10月24日港珠澳大桥正式通车,由珠海口岸清关的国际快递转运至其香港转运中心(DHL全球三大转运中心之一)的时间将从原来的4小时缩短为45分钟,大大提升了转运时效。而这对于专业做国际限时快递服务的DHL来说,尤为重要。
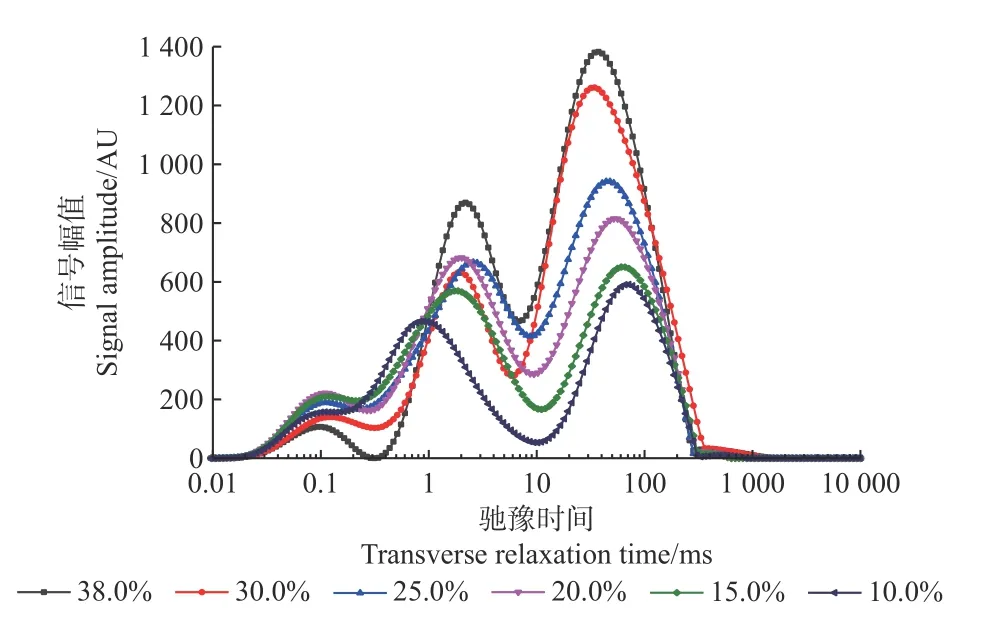
图2 失水过程中沼生栎种子的T2 反演图谱Fig.2 Spectrum inversion of transverse relaxation time T2 of Q.palustris seeds during desiccation
2.4 失水对沼生栎种子内水分相态的影响
沼生栎种子失水过程中T2反演谱结果见表2,由各组分水的峰起始点和结束点可以得到其弛豫范围。失水过程中,束缚水的弛豫范围呈减小趋势;含水量下降至25.0%时,弛豫范围由原来的0.34 ms 显著减小至0.26 ms(P<0.05);含水量下降至10.0%时,弛豫范围再次显著减小至0.17 ms。失水过程中自由水的变化较束缚水复杂,其弛豫范围呈先增大(38.0%~30.0%)后显著减小(30.0%~25.0%)再小幅波动的变化趋势(25.0%~10.0%):当种子含水量下降至30.0%时,自由水的弛豫范围由对照的757.23 ms 显著增加至997.97 ms,增加了31.79%;当含水量下降至25.0%时,弛豫范围明显减小至308.91 ms,较对照下降了59.21%。失水过程中不易流动水弛豫范围的变化趋势与自由水相反,呈先减小(38.0%~30.0%)后增大(30.0%~15.0%)随后小幅波动(15.0%~10.0%)的变化趋势,但变化幅度明显小于自由水。当含水量下降至20.0%时,其弛豫范围由对照的6.41 ms 显著增大至10.26 ms。总的来说,沼生栎种子失水过程中,氢质子自由度逐渐降低,种子内束缚水、自由水的弛豫范围均明显减小,说明失水过程中水分与营养物质的结合能力在逐渐变强。

表2 失水过程中沼生栎种子T2 反演结果①Tab.2 T2 inversion of Q.palustris seeds during desiccation
沼生栎种子失水过程中各峰顶点时间的变化见图3。失水过程中,束缚水的峰顶点时间变化较小,维持在0.11 ms 左右,各结果间差异不显著。不易流动水的峰顶点时间总体呈减小趋势,即峰顶点向左偏移,特别是失水至15.0%时,峰顶点由对照的2.26 ms 显著向左偏移至1.72 ms;当含水量下降为10.0%时,峰顶点又继续向左显著偏移至1.04 ms,水分流动性明显减弱。失水过程中,自由水峰顶点时间整体向右偏移,但失水初始阶段(38.0%~30.0%),峰顶点变化不显著;当含水量下降至25.0%时,峰顶点时间由对照的39.59 ms显著增加至60.76 ms,增加了53.47%;当含水量下降为10.0%时,峰顶点达到最大值77.69 ms。为减缓水分的散失,沼生栎种子内的不易流动水会使细胞结构变得更紧密,使水的结合能力变强;而自由水由于具有较好的流动性,较容易散失。
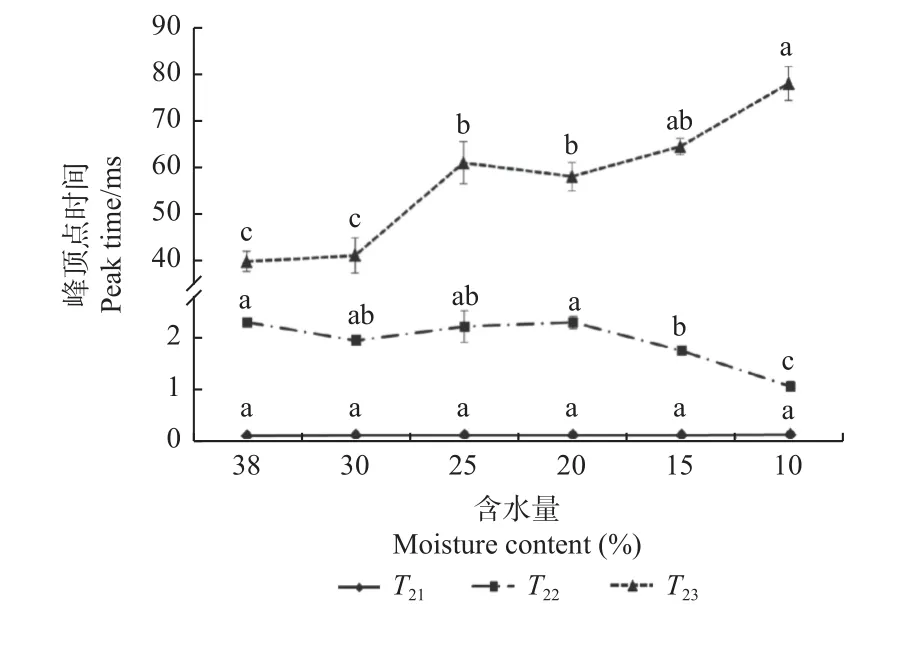
图3 失水过程中沼生栎种子各组分峰顶点时间变化Fig.3 Changes of peak time of each phase of water of Q.palustris seeds during desiccation
2.5 核磁共振T2 弛豫谱峰面积与沼生栎种子含水量的关系
沼生栎种子核磁共振T2弛豫谱峰面积与含水量的拟合曲线见图4。在显著水平α=0.05 进行回归分析,发现沼生栎种子含水量与峰面积之间具有明显的线性关系,其线性回归方程为y= 21 132x+ 698.05,其中,y为核磁共振T2弛豫谱峰面积;x为样品的含水量,相关系数R²=0.999 6。
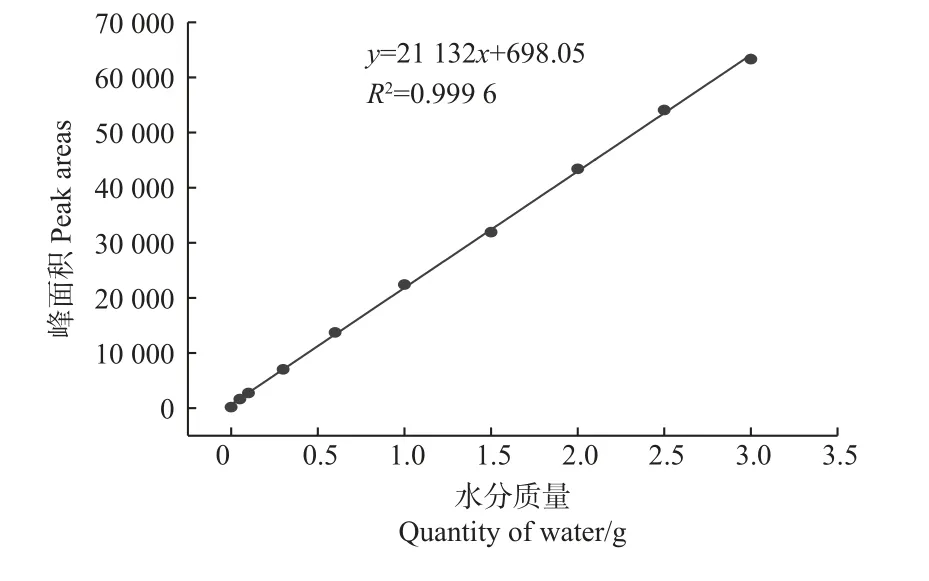
图4 沼生栎种子核磁共振T2 弛豫谱峰面积与含水量之间的回归关系Fig.4 Regressive relationship between NMR peak area of T2 relaxation spectrum and moisture content of the Q.palustris seeds
2.6 沼生栎种子失水过程中各峰面积及其比例的动态变化
沼生栎种子失水过程中3 种相态水分的峰面积及峰面积比例的动态变化见图5,其中T21、T22、T23所对应的峰面积分别为S21、S22、S23,S为3 种相态水含量的总和。随着含水量的下降,总峰面积呈下降趋势,且各处理间差异显著。当含水量下降至10.0%时,总峰面积由原来的62 908.25 降至31 569.89,下降了49.82%,但失水过程中各个相态水分峰面积的变化趋势不同:随着失水时间的延长,自由水峰面积总体呈下降趋势:含水量由38.0%下降至30.0%时,自由水峰面积下降不显著,其余各处理间差异显著;含水量下降至25.0%时,峰面积由对照的41 766.89 下降至28 325.78,下降了32.18%;含水量下降至10.0%时,自由水峰面积仅为14 930.85,较对照下降了26 836.04(64.25%),而此时总峰面积下降了31 569.89,自由水下降的量占总水分的85.01%,说明自由水散失较多。与自由水变化趋势不同,失水过程中,束缚水峰面积呈先上升(38.0%~20.0%)后下降(20.0%~10.0%)的变化趋势:含水量下降至20.0%时,束缚水峰面积由对照的2 692.78 显著上升至4 989.20,上升了85.28%;当含水量下降为10.0%时,束缚水峰面积显著下降至3 008.11,但仍略高于对照。不易流动水峰面积变化较为复杂,失水过程中其峰面积呈先下降(38.0%~30.0%)后上升(30.0%~25.0%)再下降(25.0%~10.0%)的变化趋势:含水量下降至30.0%时,不易流动水峰面积由对照的18 286.33 显著下降至13 391.27,下降了26.77%,下降幅度远高于同阶段的自由水,这可能是由于失水时,部分不易流动水向束缚水转化,使得束缚水增多;随后不易流动水峰面积上升,当含水量降为25.0%时,峰面积上升至最大值20 039.86,这可能是由于失水时部分自由水向不易流动水转化;当种子继续失水时,种子内不易流动水的峰面积再次显著下降。
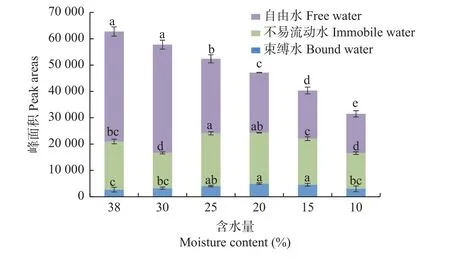
图5 失水过程中沼生栎种子峰面积变化Fig.5 The changes of peak area of Q.palustris seeds during desiccation
失水过程中,沼生栎种子各相态水分比例的变化趋势各不相同(图6)。新鲜种子中,自由水比例最高,占66.39%,而束缚水和不易流动水比例远低于自由水,分别占4.28%、29.07%。随着失水时间的延长,自由水峰面积比例总体呈下降趋势,当含水量下降至10.0%时,峰面积比例显著下降至47.29%;但在含水量由38.0%下降至30.0%的过程中,自由水的比例稍有上升。失水过程中,不易流动水的比例呈先下降再上升的变化趋势:含水量由38.0%下降至30.0%的过程中,不易流动水的比例下降;而当含水量从30.0%下降至25.0%时,该比例大幅增加至38.18%;当含水量降为10.0%时,不易流动水的比例缓慢增加至42.98%。这可能是种子在受失水胁迫时,为减缓水分的散失,不易流动水与大分子物质结合能力变强。失水过程中,束缚水由于本身性质较为稳定,且失水过程中部分不易流动水也会向束缚水转化,因此,含水量由38.0%下降至15.0%的过程中,峰比例一直呈上升趋势,由对照的4.28%上升至11.27%;随后峰比例下降至9.53%(含水量10.0%),束缚水比例的动态变化也说明,由于束缚水与营养物质间具有较强的结合能力,才有效地减缓了水分的散失速率,从而降低失水对种子的伤害。
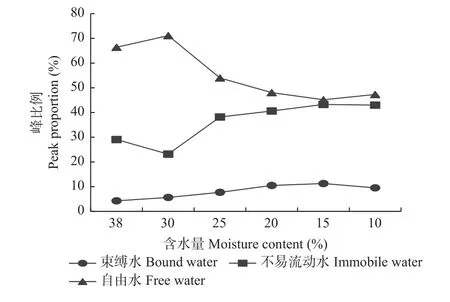
图6 失水过程中沼生栎种子峰比例变化Fig.6 The changes of peak proportion of Q.palustris seeds during desiccation
3 讨论
3.1 失水对沼生栎种子发芽率的影响
栎属植物种子成熟时未经历脱水阶段,所以散落时仍具有较高的含水量和代谢活性(Ntuliet al.,2014),表现出顽拗性种子的特征,具有脱水敏感性(Xiaet al.,2012b)。程继铭等(2019)在研究脱水和低温对辽东栎(Q.wutaishansea)种子萌发的影响中发现,无论快速脱水还是缓慢脱水,种子的萌发率和活力指数都随脱水时间的延长而下降。温祺(2010)也发现栓皮栎(Q.variabilis)种子的生活力随风干时间的延长而不断下降。本研究发现,沼生栎种子在自然失水过程中发芽率显著下降。当含水量降低至25.0%时,种子的发芽率由对照的96.00%显著下降至81.33%;当含水量进一步下降至10.0%时(正常性种子的安全含水量),沼生栎种子完全丧失发芽能力,说明含水量的降低对沼生栎种子质量影响较大。因此,沼生栎种子表现出脱水敏感性,为顽拗性种子,在贮藏运输过程中,需使种子维持较高的含水量。
3.2 沼生栎种子失水过程中各相态水分T2 弛豫时间的变化
失水过程中随着种子含水量的变化,各相态水分的结合能力也会发生改变。本研究发现沼生栎种子失水过程中,自由水弛豫时间范围先增大后减小,峰顶点整体向右偏移,且T23在含水量为10.0%时达到最大值;不易流动水的弛豫时间范围总体变大,峰顶点在含水量下降至20.0%后显著向左偏移;束缚水性质较为稳定,弛豫时间范围和峰顶点变化不大。孙旭等(2019)研究认为,银杏种子干燥过程中自由水变化较为显著,干燥220 min 时,自由水的峰顶点时间达到最大值;束缚水的峰顶点时间变化不大,这与本研究结果相似。宋姗姗等(2020)研究发现,板栗种子在相对湿度 70%, 25 ℃条件下贮藏过程中,前14 天束缚水的横向弛豫时间不断向左移而后稳定;而不易流动水的横向弛豫时间则是右移;自由水横向驰豫时间先向右移动后左移。不同顽拗性种子在失水或干燥过程中种子各相态水分的变化规律不同,这可能与种子中营养物质的种类、贮藏条件以及失水方式不相同等有关。由于自由水在细胞内呈游离状态,可自由流动,因此失水过程中,沼生栎种子内自由水峰顶点整体向右偏移;随着失水时间的延长,种子内淀粉、蛋白质等营养物质浓度的升高,种子为维持细胞膜的稳定性,减少失水胁迫带来的伤害,会使不易流动水和束缚水峰整体向左偏移,从而使水的结合能力增强。
3.3 沼生栎种子失水过程中各相态水分峰面积及比例的动态变化
失水过程中,沼生栎种子中各相态水分峰面积及比例也在不断变化。当含水量下降至25.0%时,自由水峰面积较对照显著下降了32.18%,而此时不易流动水、束缚水的峰面积较对照却显著增加了9.60%、50.20%,这可能是由于自由水向外散失的同时,有部分自由水向不易流动水转化,同样束缚水峰面积的显著增加也可能是由于部分不易流动水转化为束缚水造成;当含水量继续下降至20.0%时,束缚水峰面积及比例仍是持续增大,但不易流动水峰面积却在减小,说明此阶段仍有一部分不易流动水向束缚水转化。由此可推测,即使是顽拗性种子,为减缓水分的散失,会采取措施使水分结合能力变强以维持细胞膜的稳定性。宋姗姗等(2020)在板栗种子失水的研究中同样发现,贮藏前14 天,胚为了保持自身活性,与淀粉、蛋白质等紧密结合,使束缚水所受约束力不断增强;随后束缚水相对稳定,部分不易流动水向自由水转化。
4 结论
1)随着含水量的下降,沼生栎种子发芽率不断下降,含水量下降至25.0%时,发芽率显著下降。含水量降为10.0%时,种子完全死亡。根据含水量与发芽率的拟合曲线得出其半致死含水量为17.39%。沼生栎种子持水能力较弱,不耐脱水,不能像正常种子那样长时间储存。
2)沼生栎种子含水量与核磁共振T2驰豫谱峰面积有显著的线性关系。
3)T2弛豫图谱中各峰从左至右依次为为束缚水T21、不易流动水T22以及自由水T233 种相态。
4)沼生栎种子失水过程中,自由水峰面积及峰比例持续降低,峰向右偏移;不易流动水峰面积呈先下降后上升再下降的变化趋势,峰比例呈先下降后上升的趋势,峰顶点整体向左偏移,与水的结合能力增强;束缚水本身性质较稳定,且失水过程中不易流动水向束缚水转化,水的结合能力变强。
猜你喜欢
中外文摘(2022年15期)2022-11-21
艺术品鉴(2019年8期)2019-09-18
种子科技(2018年11期)2018-09-10
许昌学院学报(2018年8期)2018-09-05
小学生优秀作文(低年级)(2017年11期)2017-10-18
上海铁道增刊(2017年2期)2017-04-18
材料科学与工程学报(2016年4期)2017-01-15
高原山地气象研究(2016年3期)2016-02-28
天津农林科技(2015年1期)2015-12-30
中国塑料(2015年4期)2015-10-14
