
树形调整对香榧成花和坐果的影响*
2023-12-11 03:00吴家胜仲嘉玥胡渊渊喻卫武俞晨良宋丽丽索金伟
林业科学 2023年11期
成 豪 吴家胜 马 爽 仲嘉玥 胡渊渊 喻卫武 俞晨良 宋丽丽 索金伟
(浙江农林大学 亚热带森林培育国家重点实验室 杭州 311300)
香 榧(Torreya grandiscv.Merrillii)是红豆 杉 科(Taxaceae)榧树 属(Torreya)榧 树(Torreya grandis)中的优良栽培类型,是我国特有的珍稀干果树种,属国家二级保护植物。其坚果风味香酥、余味浓郁,营养丰富,尤其是富含多不饱和脂肪酸和多种生物活性成分,具有很高的保健、药用和综合开发利用价值(Wuet al., 2018; Louet al., 2019; Suoet al., 2019)。近些年,香榧产业不断发展,截至到目前,香榧种植区域覆盖浙江、安徽、江西、贵州等全国8 个省市,面积已达10.67 余万hm2。然而,在香榧幼年林经营过程中发现,香榧幼年林分容易树体生长过旺,混合芽形成数量少、坐果率低、结实能力弱,呈现‘光长树不结果’现象,缺乏科学的栽培管理技术是导致此问题的主要原因之一。
树体结构调整是影响果树产量的关键技术,合理的骨干枝数目和拉枝角度选择是树体结构调整的核心内容。骨干枝数目直接决定了果树的树形,是影响树冠光照分布的主要因素。而拉枝对培养树体骨架结构、调整枝条空间分布、改善树体通风透光条件、改变枝条极性、促进或抑制枝条生长、平衡果树营养生长与生殖生长等方面都具有重要作用。研究发现,苹果(Malus pumila)树形改造过程中,与单主枝的主干疏层形相比,多主枝开心形树冠大而扁,树冠透光率高;并且多主枝开心形树体的短果枝占结果枝的60.68%,中果枝占28.67%,利于连年丰产,坐果率高(谢鹏等, 2018; 白岗栓等, 2021)。同样,无花果(Ficus carica)、 蓝 莓(Vacciniumspp.)、 板 栗(Castanea mollissima)等树形培养中也发现,合理的骨干枝数目选择,有效提高了果实单果质量、可溶性固形物含量等,明显提高了产量。此外,张满让(2013)对苹果树拉枝研究表明,拉枝可以缓和树势,较小的拉枝角度处理下,光合产物主要向上运输,有利于树体的生长;随着拉枝角度加大,光合产物向下运输增加显著,抑制树体营养生长,新生枝条的顶梢尖总糖含量和碳氮比值(C/N)增加,而顶梢尖的总氮含量降低,有利于花芽分化(李长亮等, 2018; 刘元媛等, 2021)。在苹果、‘翠冠’梨(Pyrus pyrifoliacv.‘Cuiguan’)、核桃(Juglansregia)等研究中也发现,适宜的拉枝角度可以有效影响枝条内源激素含量,促进成花,但不同果树适宜的拉枝角度不同,例如,核桃拉枝90°处理下,枝条中赤霉素、生长素与脱落酸比例下降,玉米素与生长素比例显著増大,利于核桃花芽分化(李晓龙等, 2015; 苗雅坤, 2020);当‘富士’(Malus pumilacv.Fuji)苹果拉枝角度为110°时,枝条顶芽IAA 和GA3含量减少,ABA和ZR 含 量 增 加,ABA/IAA、ABA/GA3、ZR/IAA 和ZR/GA3值也逐渐变大,花芽形成数量较对照明显提升(杨勇等, 2011; 余拱鑫, 2012)。
综上所述,科学合理的树体结构调整不仅有利于果树幼树树形培养,同时可显著促进开花结果,提高产量等。近年来,有关树体结构调整技术在核桃等坚果类树种中也有少量报道,但总体而言其在坚果类树种中并未得到广泛的推广应用。香榧作为我国南方特色干果,目前其高产栽培理论与配套树形调控技术仍不完善,关于树体结构调整如何影响其成花坐果的系列理论研究仍未见报道。因此,本研究针对香榧幼树树体结构调整展开研究,分别设置不同的骨干枝数目和骨干枝开张角度处理,分析其对树体光照强度、混合芽比例、内源激素含量、光合碳同化物含量等的影响,以揭示树体结构调整对香榧开花和坐果的影响,探索出香榧幼树适宜的骨干枝数目和开张角度,为开展香榧幼年林树形及树体结构管理,促进早实丰产提供科学依据。
1 材料与方法
1.1 试验地概况与试验材料
试验地位于浙江省杭州市临安区浙江农林大学潘母岗实验基地(30°14′N,119°42′E)。属亚热带季风气候,温暖湿润,雨量充沛,日照充足;全年平均气温17.8 ℃,平均相对湿度70.3%,年降水量1 454 mm,年日照时数1 765 h。试验材料为2007 年种植的香榧树,株行距4 m×5 m,树龄19 年,平均地径为12.2 cm,平均树高为1.8 m 的初结果香榧幼年林。试验区内0~20 cm 土壤中全氮质量分数为1.2 g·kg-1,全磷质量分数为1.1 g·kg-1,全钾质量分数为15.71 g·kg-1。基地采用统一的常规施肥管理措施,每年施用复合肥平均2~3 次,约750~1 000 g/株/年,有机肥每2 年施用1 次。
1.2 试验方法
1.2.1 试验处理 树体结构调整包括骨干枝数目和骨干枝开张角度2 个单因素试验,并在单因素基础上进行交互试验;骨干枝数目试验:共设置单主枝自然圆头形(对照)、3 主枝和4 主枝开心形3 个处理,分别标记为CK、N3、N4,每个处理5 次重复;骨干枝开张角度试验:共设置30°(对照)、60°和90° 3 个处理,分别标记为A30°、A60°和A90°,每个处理5 次重复。选择长势一致的香榧树,将主干枝先进行简单揉压,使用拉枝绳在主干枝条的基部进行抻拉,缓慢拉动直到调节至合适角度。在以上单因素基础上,选择效果较好的单因素进行交互处理:骨干枝数目保留CK、3 骨干枝和4 骨干枝处理,骨干枝开张角度保留30°和60°处理,交互试验各处理分别标记为CK-A30°、N3-A30°、N4-A30°、CK-A60°、N3-A60°、N4-A60°,采用双因素随机区组试验,每个处理5 次重复。
1.2.2 花果发育、混合芽比例及坐果率调查 香榧雌雄异株,通常混合芽于3 月底开始萌动,出现形态分化,4 月中旬左右传粉滴出现并授粉,授粉后雌配子体逐渐发育成熟,至9 月中旬左右开始受精(刘志敏等, 2017);同时,受精后的幼果次年4 月初开始启动发育,逐渐突破种鳞,5 月初完全突破种鳞即成功坐果(图1)。因此,于每年3 月分别从东南西北四个方位中选择结果枝组40 个进行挂牌标记,调查并计算混合芽比例、叶果比、叶花比和坐果率。混合芽% = 混合芽数/芽总数×100%;叶花比 = 叶片总数/花总数;叶果比 = 叶片总数/幼果总数;坐果率% = 总坐果数/总雌花数×100%。
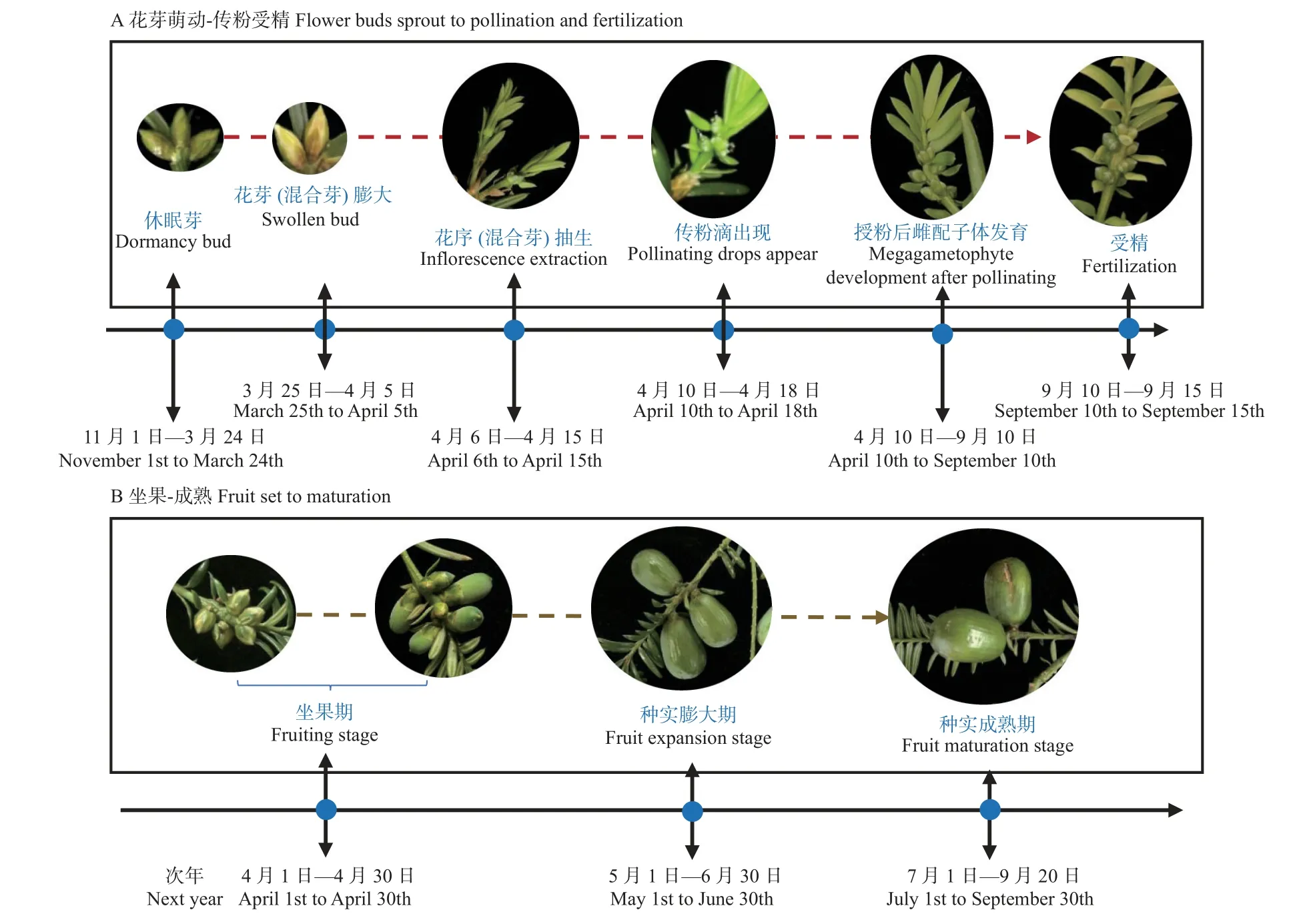
图1 香榧成花坐果规律Fig.1 Survey of flower bud development and fruit setting of Torreya grandis cv.Merrillii
1.2.3 树冠光照度和叶片净光合速率测定 为了探究树形调整对香榧树冠光照和叶片光合能力的影响,分别使用TES-1339R 照度计和Li-6 400 便携式光合仪(LI-COR Biosciences, Inc.)对各处理和对照树进行光照强度和净光合速率测定。选择天气晴朗且无云的天气,测量时间为上午9:00—11:00。光照度测量以树干为中心,将树干分为东南西北四个方向,从树干垂直方向由上至下分为上层、中层、下层,每层间隔50 cm。TES-1339R 照度计量程在(0.01~999)×100 lux之间,精确度在 ± 3%(符合NIM 标准)。测定温度范围在0~50 ℃之间,测量数据为屏幕显示数据(单位:lux)。净光合速率测定,光合有效辐射(PAR)设定为1 200 μmol·m-2s-1,空气流速设置为400 μmol·s-1,叶室温度设定为25 ℃。
1.2.4 内源激素含量测定 为了探究树形调整对香榧成花坐果期混合芽和幼果中内源激素含量的影响,分别于4 月5 日和4 月30 日采集骨干枝开张角度处理(A30 干、A60°、A90°)条件下香榧混合芽和成功坐果的幼果进行内源激素含量测定。激素含量测定采用高效液相色谱及质谱联用方法,具体参考Zhao 等(2019)的方法,每个样品4 次生物学重复。
1.2.5 光合碳同化物含量测定 为了探究树形调整对香榧成花坐果期混合芽和幼果中光合碳同化物含量的影响,于4 月5 日采集骨干枝数目调整(CK、N3、N4)条件下香榧的混合芽,于4 月30 日分别采集骨干枝数目调整(CK、N3、N4)和交互处理(CK-A30°、N3-A30°、N4-A30°、CK-A60°、N3-A60°、N4-A60°)条件下香榧成功坐果的幼果,进行光合碳同化物含量测定。葡萄糖、果糖、蔗糖等含量测定采用蒽酮比色法(曹健康等, 2019),简要过程如下:取鲜样组织样品0.2 g,无菌水振荡混匀后80 ℃水浴30 min,过滤后定容至2 mL,上清液50 ℃水浴3 min,冷却后于620 nm 和640 nm下分别测量吸光值;将测得的吸光度值代入标准曲线计算光合碳同化物含量,每个样品4 次生物学重复。
1.2.6 碳氮比分析 为了探究树形调整对香榧混合芽碳氮比的影响,于4 月5 日采集骨干枝开张角度调整(A30°、A60°、A90°)条件下香榧的混合芽进行总碳和总氮含量测定。测量仪器选用美国元素分析仪(Flash SMART)。样品在锡或银容器中称量,然后放入样盘中,通过球阀将样品自动进样至燃烧管。使用惰性气体吹扫样品以去除进样时带入的空气干扰。样品在高温下燃烧或裂解,待测元素转化为气体产物,用程序升温脱附(TPD)分离分析气体,在TCD (或可选的IR)中检测。利用检测器信号,样品质量和储存的校准曲线,计算出元素浓度。
1.3 数据分析
试验结果用至少3 次重复的平均值±标准误差表示。采用IBM SPSS Statistics 25 软件对数据进行单因素分析,采用 Microsoft Excel 2013 进行图表绘制。
2 结果与分析
2.1 树形调整对香榧成花坐果的影响
2.1.1 骨干枝数目调整 光照度测定发现,骨干枝数目调整对香榧树体光照度影响较大,随着香榧骨干枝数目的增加,树体透光能力更强,与对照相比,3 和4骨干枝显著增加了树冠中层和下层的光照强度,例如,3 骨干枝中层的平均光照强度为214.57×100 lux,约是对照的1.5 倍;而3 和4 骨干枝下层的平均光照强度达到132.96×100 lux 和126.15×100 lux,较对照分别提高了13%和7%(图2)。
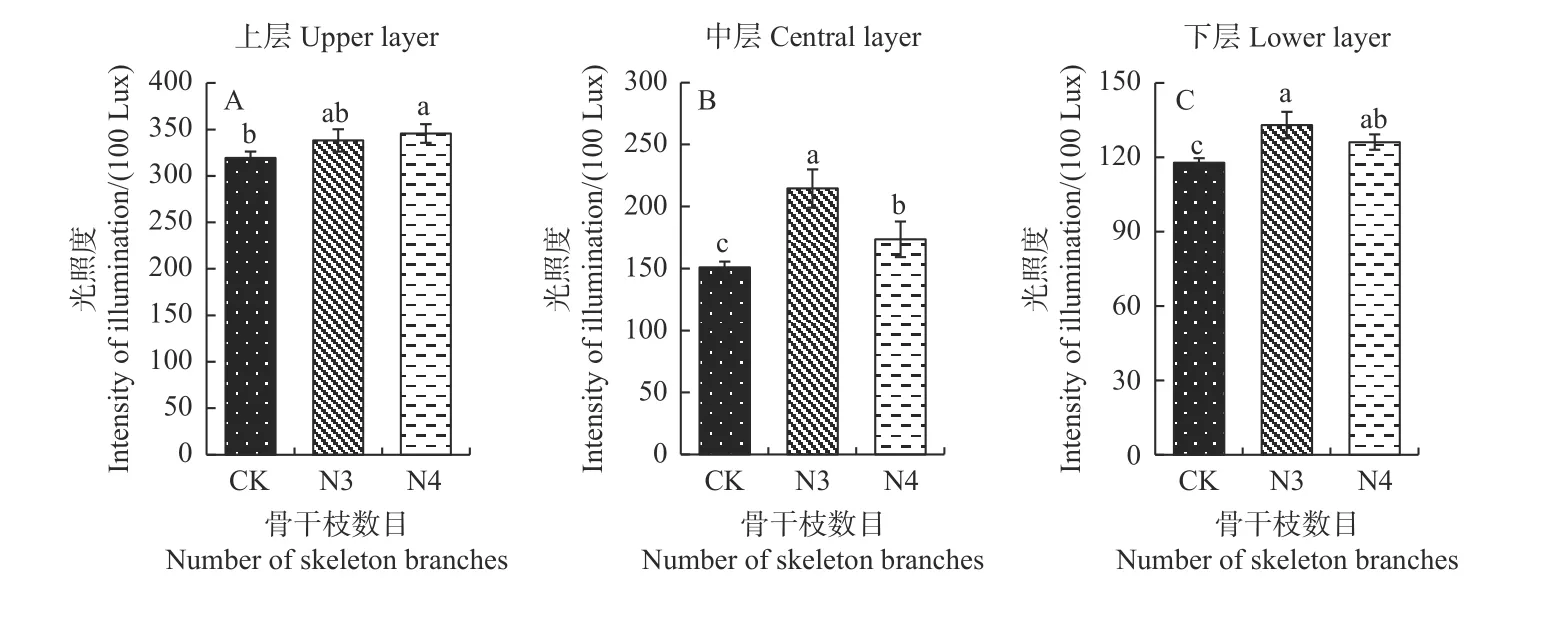
图2 骨干枝数目调整后香榧树冠光照强度变化Fig.2 The change of light intensity of T.grandis after setting skeleton branch number
骨干枝调整后,坐果率均明显提高。其中,3 骨干枝和4 骨干枝处理下,坐果率较对照(8%)分别提高了1.2 和1.3 倍(图3)。此外,随着骨干枝数目的增加,香榧混合芽比例也明显提高,3 骨干枝和4 骨干枝处理下混合芽比例由CK 的45.74%分别提高到53.51%和54.37%(图3)。
2.1.2 骨干枝开张角度调整 由图4 可知,骨干枝开张角度处理对香榧树体成花和坐果均有较大影响。其中,较对照而言,开张角度60°处理后坐果率显著增加,达到12.55%;开张角度60°处理也显著提高了香榧树体的混合芽比例,增加到60%左右。90°处理下,香榧树体坐果率与对照相比差异不显著,但显著降低了香榧树体的混合芽比例,由对照的54%降低到38%。表明骨干枝开张角度的适当增加,有利于香榧成花坐果,而开张角度过大则会抑制成花坐果(图4)。
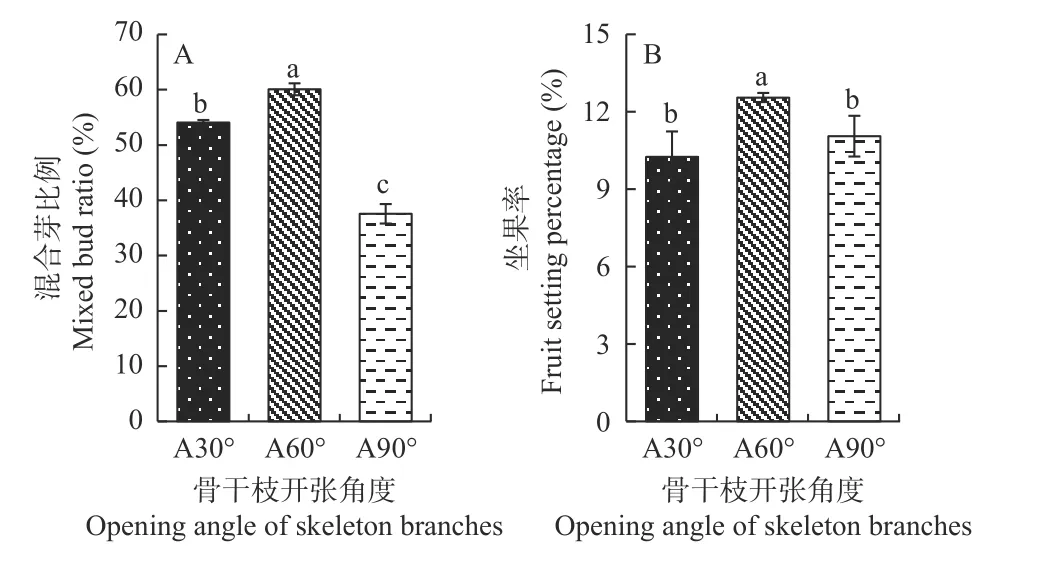
图4 骨干枝开张角度调整后香榧混合芽比例及坐果率分析Fig.4 The change of mixed bud proportion and fruit setting rate of T.grandis after setting skeleton branch opening angle
进一步对混合芽及幼果中的内源激素和光合碳同化物含量分析发现,开张角度60°处理一定程度上增加了香榧混合芽中ABA、细胞分裂素(tZR 和DHZ)等含量,但与对照(A30°)相比差异不显著;开张角度60°处理显著降低了IAA、GA1和GA3含量,其中GA1和GA3含量与对照相比分别下降了48.03%和26.73%(表1);开张角度90°处理显著降低了ABA 含量(P<0.05),但显著增加了tZ、tZR、DHZ 含量,同时也显著增加了IAA 含量(P<0.05)。混合芽中不同内源激素间的比值分析发现,开张角度60°处理下混合芽中ABA/IAA 和ABA/GA1+3比值明显升高,CTKs/IAA和CTKs/GA1+3比值与对照相比也略有升高(P<0.05)(表1)。此外,幼果中激素含量分析发现,开张角度60°处 理 显 著 增 加 了 幼 果 中GA1、GA3、DHZ 含 量,GA1+3/ABA 和CTKs/ABA 比值显著提高(P<0.05)。开张角度90°处理对幼果中GA1和GA3含量无显著影响,但显著降低了IAA 含量,同时显著提高了幼果中的ABA 含量(表2)。
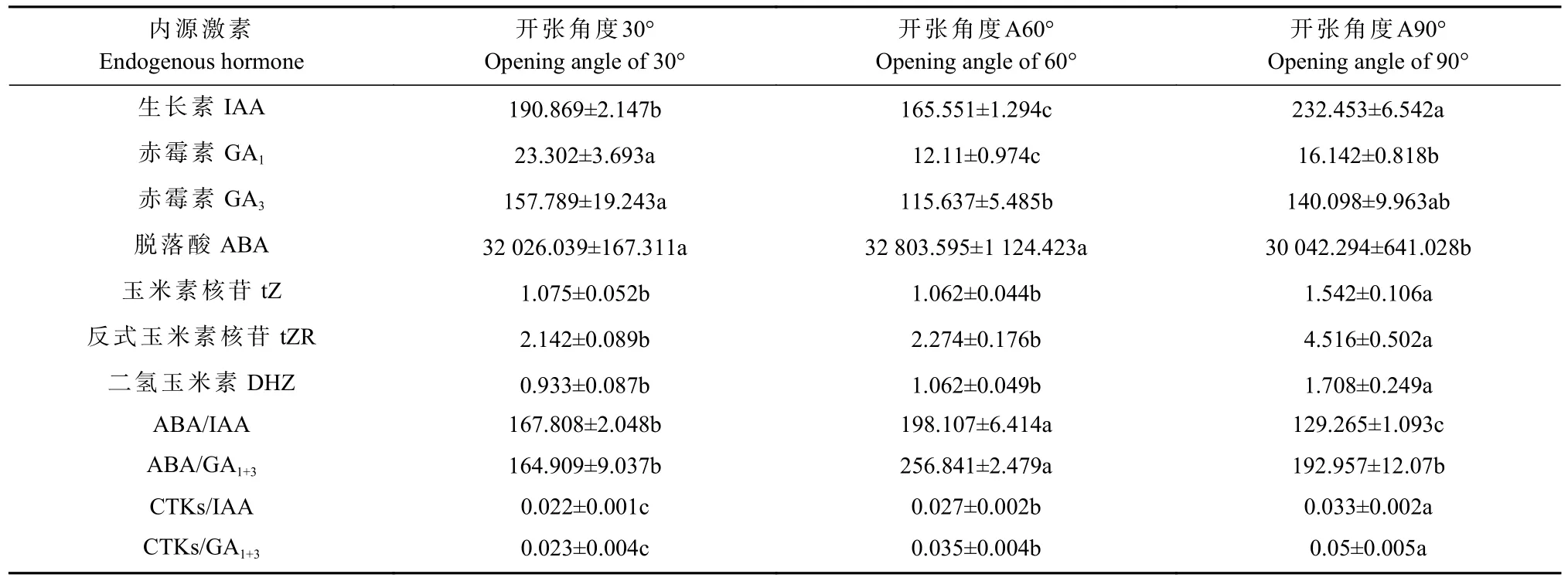
表1 骨干枝开张角度调整后香榧混合芽中内源激素含量分析①Tab.1 Analysis of endogenous hormone conte nt inmixed buds of T.grandis after setting skeleton branch openingangle ng·g-1
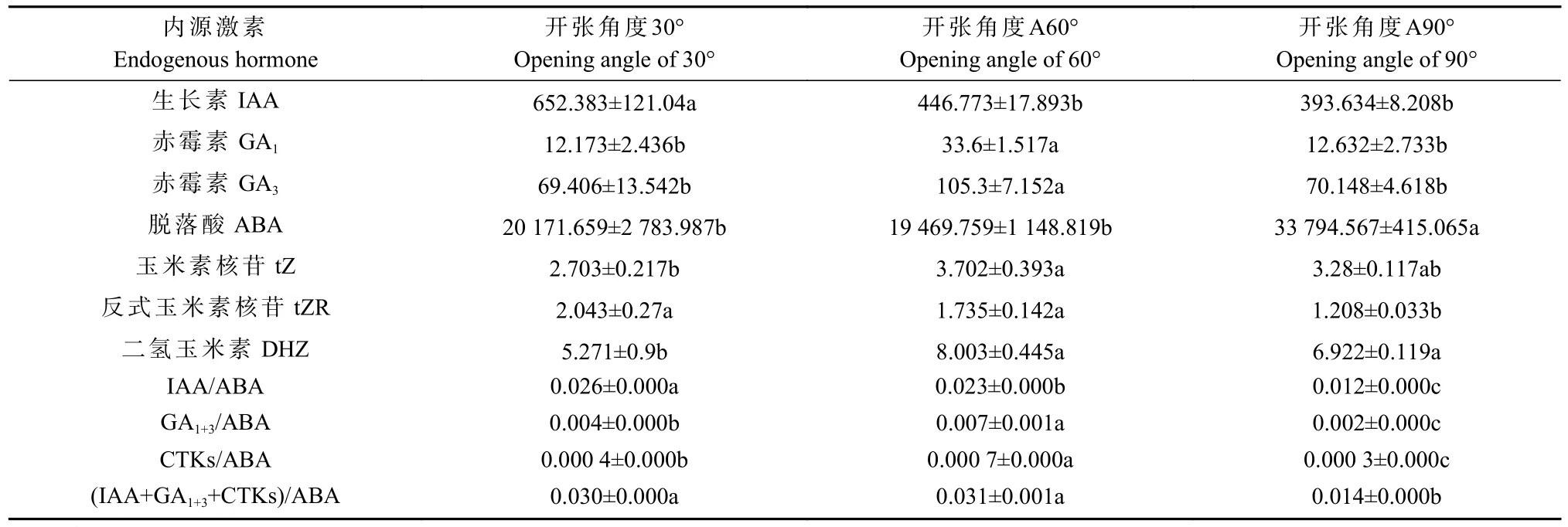
表2 骨干枝开张角度调整后香榧幼果中内源激素含量分析①Tab.2 Analysis of endogenous hormonecontentin young fruits of T.grandis after setting skeletonbranch opening angleng·g-1
光合碳同化物和碳氮比分析发现,开张角度60°处理一定程度上增加了混合芽中葡萄糖和蔗糖含量,显著增加了混合芽中果糖、总可溶性糖和淀粉含量(P<0.05);90°处理下混合芽中光合碳同化物总体呈现降低趋势(图5A-B)。此外,开张角度60°处理显著增加了幼果中的光合碳同化物含量,与对照相比,果糖、葡萄糖、蔗糖分别提高了39.5%、40.43%和18.64%,可溶性糖和淀粉含量分别提高了40.68%和15.42%;开张角度90°处理下幼果中果糖、葡萄糖、蔗糖及淀粉含量与对照相比无显著变化(图5C-D)。C/N 比分析发现,开张角度60°处理显著增加了混合芽中总碳含量,并显著降低总氮含量,因此显著提高了C/N 比例(P<0.05),而开张角度90°处理与对照相比无明显变化(图6)。
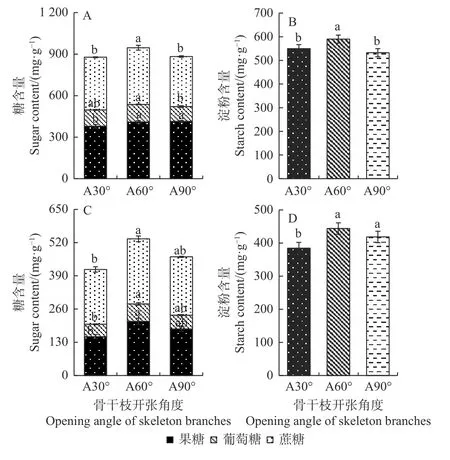
图5 骨干枝开张角度调整后香榧混合芽(A、B)和幼果(C、D)中光合碳同化物含量分析Fig.5 The change of photosynthetic carbon assimilate content after setting skeleton branch opening angle in mixed bud (A, B) and young fruit (C, D) of T.grandis

图6 骨干枝开张角度调整后香榧混合芽碳氮比分析Fig.6 The change of carbon to nitrogen ratio after setting skeleton branch opening angle in mixed bud of T.grandis
2.1.3 骨干枝数目和开张角度交互处理 在单因素试验基础上,选出效果较好的3 和4 骨干枝与开张角度60°单因素进行交互处理。研究发现,开张角度60°的所有处理中,香榧树体上层、中层、下层光照强度均显著高于其他处理。其中,3 骨干枝和4 骨干枝与开张角度60°交互处理显著提高了中层和下层光照强度,例如,N3-A60°和 N4-A60°处理下中层光照强度达到25 000~27 000 lux,下 层 光 照 强 度 达 到11 000~14 000 lux,分别是对应单因素试验的1.3 倍和1.5 倍左右(表3)。
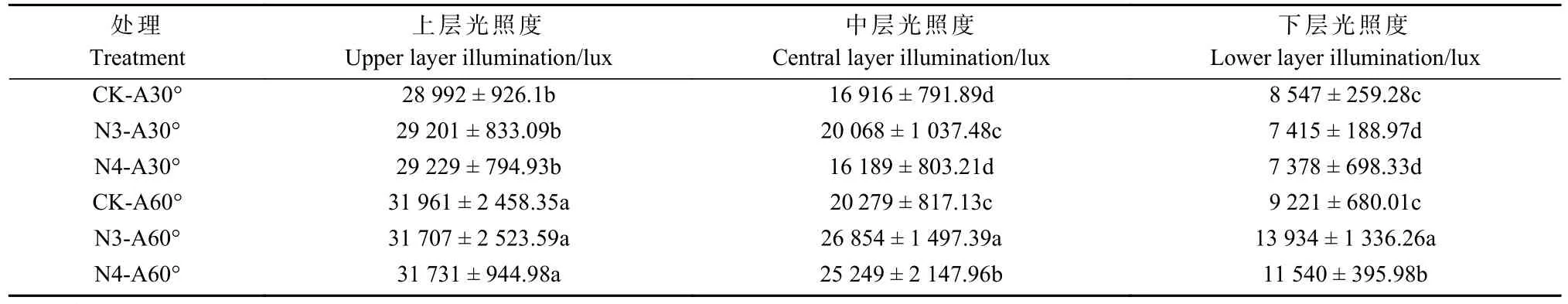
表3 骨干枝数目和开张角度交互处理对香榧树冠光照强度的影响①Tab.3 The change of light intensity of T.grandis after setting skeleton branches number and opening angle
光合碳同化物含量分析发现,交互处理下香榧坐果期的幼果中光合碳同化物含量有所提高。其中,N3-A60°和N4-A60°处理下葡萄糖、果糖、总可溶性糖、淀粉含量比单独3 骨干枝和4 骨干枝处理显著提高(P<0.05)(表4);此外,叶果比和叶花比调查发现,骨干枝数目和开张角度的单因素和交互处理下叶果比和叶花比均差异不显著,叶果比在7.6∶1~8.3∶1 之间,叶花比约为56.2∶1~63.2∶1(图7 A-B)。而净光合速率分析发现,开张角度A60 合以及交互处理下净光合速率明显高于其它处理,其中N3-A60°和N4-A60°处理下净光合速率分别达到9.07 和9.70 μmol CO2·m-2s-1(图7C)。坐果率分析发现,骨干枝和开张角度的交互处理显著提高了香榧坐果率,其中,N3-A60°和N4-A60°处理下的坐果率分别达到12.5%和15.6%,而对照和其他单因素处理下的坐果率不足10%(图7D)。
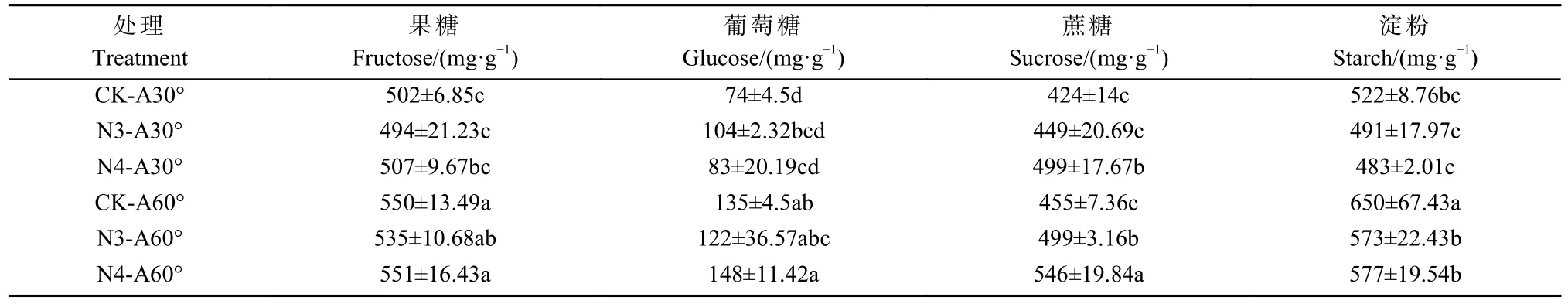
表4 骨干枝数目和开张角度交互处理对香榧幼果光合碳同化物含量的影响①Tab.4 The change of photosynthetic carbon assimilate content of T.grandis after setting skeleton branches number and opening angle
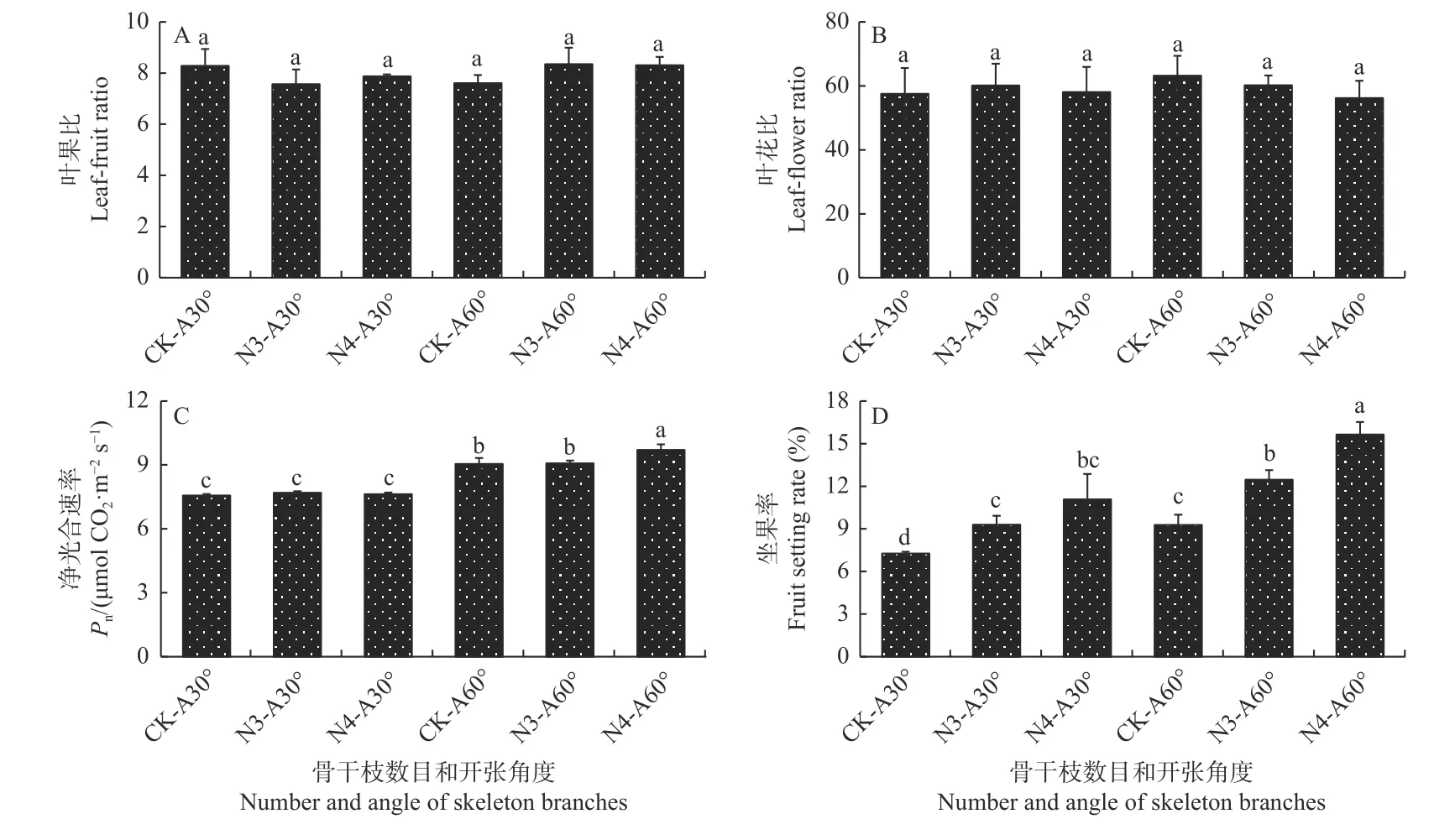
图7 骨干枝数目和开张角度交互处理对香榧叶果比(A)、叶花比(B)、净光合速率(C)和坐果率(D)的影响Fig.7 The change of T.grandis leaf-fruit ratio (A), leaf-flower ratio (B), net photosynthetic rate (C), and fruit setting rate (D) after setting skeleton branch number and opening angle
3 讨论
香榧是浙江最具特色的珍稀干果树种,也是高产优质的木本油料树种,成年香榧亩产值超万元,栽培效益居山地经济林之首。然而,香榧栽培管理过程中发现,幼年林分的香榧树体容易生长势过旺,成花及坐果率低(叶淑媛, 2020)。在前期的物候调查中也发现,香榧成花和坐果过程在时间上存在一定重叠,二者同时作为库器官竞争源叶运输的光合同化物,导致营养竞争大(图1)。而不合理的树体结构可能更加加剧了这种竞争,对成花和坐果产生不利影响。在‘富士’系‘富姬酷’(Malus pumilacv.Fujiko)苹果树中也普遍存在树体旺长,成花难的问题(马小龙等,2020)。澳洲坚果树体营养生长和生殖生长极不平衡,自然情况下坐果率仅有0.3%左右(陈贞洁等, 2022)。甜樱桃(Prunus avium)通常在幼树时期表现出生长过旺,而盛果期后又较易衰弱,不易成花坐果(朱天华等,2012)。因此,需要通过人为干预来调节树体的生长发育、开花结果,以实现早实丰产的栽培目标。
3.1 树形调整对香榧成花的影响分析
果树的成花过程除了与光照、温度等外部环境条件有关外,还主要受到植物体内碳水化合物,激素,矿质元素等内在营养物质变化的影响(陈晨等, 2020)。合理的树形结构调整,如骨干枝数目的增加并辅以合适的开张角度,能够改善树体光环境,增加树体的透光能力和光合效率,有利于成花(蔡虎等, 2022)。无患子骨干枝数目调整后发现,3 骨干枝和5 骨干枝处理树体内膛光照度达到(160~300)×100 lux,是对照的2 倍左右,且明显提高了花序数量(高媛等, 2016)。鲜黄梨(Pyrus pyrifoliacv.‘Sunhwang’)研究中也发现,树势过旺会对树体内部造成遮荫,导致冠内光照分布不均匀,通过树形调整可有效改变树冠光环境,充足的光照有利于花芽形成(赵瑾, 2010; Wertheimet al.,2001)。文冠果(Xanthoceras sorbifolium)幼树树形培养中发现,多主枝丛生形的花序数为33 个,显著高于其他处理(兖攀等, 2021)。本研究中也发现,3 骨干枝和4 骨干枝处理下显著增加了香榧树冠中层和下层的光照强度,且3 骨干枝和4 骨干枝处理后香榧树体成花率达到55%左右,较对照显著提高(图2-3)。
此外,大量研究发现果树骨干枝开张角度增加可以降低树体极性,调节激素、碳水化合物等分配及平衡,降低果树顶端优势,调控树体的营养生长与生殖生长平衡(杨勇, 2010; 李荳, 2016)。苹果拉枝处理通过提高芽中营养物质(蔗糖、果糖等),C/N 比,以及生长素,细胞分裂素、脱落酸含量,有效提高了树体成花率(邢利博,2016)。杨勇等(2011)的研究也表明‘富士’110°拉枝处理和‘嘎啦’(Malus pumilacv.‘Gala’)90°拉枝处理有效解决了苹果成花难的问题。赵通等(2020)对杏树(Prunus armeniaca)进行研究,结果表明拉枝处理可以提高花芽中ABA、CTKs 含量以及ABA/IAA、ABA/GA 比例,促进杏树的花芽分化。刘元媛(2021)研究发现,随着苹果树开张度的增大,IAA 和GA3含量逐渐降低,ABA 含量逐渐增加,C/N 比也有所增加,成花率明显提高,表明经过拉枝处理,树体内部生长中碳氮平衡与激素分配发生变化,由营养生长转化为生殖生长。在本研究中,开张角度60°处理虽然对香榧混合芽中细胞分裂素CTKs(tZR 和DHZ)、ABA 含量没有大幅增加,但显著降低了IAA、GA1和GA3含量,同时提高了ABA/IAA、ABA/GA1+3,CTKs/IAA 以及CTKs/GA1+3比值,也明显增加了混合芽中光合碳同化物蔗糖,淀粉等含量和C/N 比,显著提高了树体混合芽比例(图4-6,表1)。
综上表明,香榧树体结构调整有利于香榧成花,有效提高了混合芽比例,究其根本原因,一方面骨干枝数目的增加以及合适的骨干枝开张角度增加了树体的通风透光能力,尤其是明显增强了垂直方位上树体的透光能力,显著增加了中层和下层的光照度,促进了光合碳同化物的积累;另一方面,抑制了混合芽中GA1、GA3、IAA 的积累,进而改变了ABA 和CTKs与IAA、GA1、GA3等的比例以及光合碳同化物的分配,调节了C/N 平衡,最终促进了香榧花芽的形成。
3.2 树形调整对香榧坐果的影响分析
成花与坐果过程往往相辅相成,然而香榧坐果周期较长,大约每年9 月下旬—10 月上旬受精后一直持续到次年的4 月底,更容易受到多种环境因素及内部养分等的影响。长期的野外调查发现,每年平均坐果率不足10%,而部分林分坐果率仅5%左右(叶淑媛,2020)。本研究发现,通过香榧树体结构的调整,可以有效提高香榧坐果率,单独的骨干枝数目和骨干枝开张角度增加,香榧树体坐果率较对照均明显提高,二者交互处理(N3-A60°和 N4-A60°)下坐果率提高到12.5%~15.6%左右(图7)。进一步分析发现,香榧骨干枝数目和骨干枝开张角度的交互处理有效提高了树体的光照强度,N3-A60°和N4-A60°处理下中层光照强度达到25 000~27 000 lux,下层光照强度达到11 000~14 000 lux(表2);叶果比分析发现,骨干枝数目和开张角度单因素及交互处理对叶果比没有显著影响,但N3-A60°和N4-A60°处理显著提高了叶片净光合速率和幼果中蔗糖和总可溶性糖等含量(图7,表4),表明N3-A60°和N4-A60°处理主要是通过提高光合能力来促进幼果中光合同化物的积累。
此外,开张角度A60°处理还明显提高了幼果中赤霉素GAs 和部分细胞分裂素的含量(表2)。在梨(Pyrus spp.)、苹果、桃(Amygdalus persica)、蓝莓等多种果树上均发现,拉枝以及不同骨干枝数目调整能够控制树体旺盛生长、调节树体激素的平衡,提高坐果及产量(王磊等, 2010; 吴鲜亮等, 2008)。‘库尔勒’香 梨(Pyrus sinkiangensiscv.‘Korla Fragrant’)和 枣(Ziziphus jujuba)中研究也发现,果实中GA3含量增加能明显提高可溶性糖含量,显著提高坐果率(袁振杨,2019; 樊丁宇, 2022)。细胞分裂素和赤霉素处理后油茶(Camellia oleifera)坐果率分别为26.82%和20.02%,较对照明显提高(杨钧钰, 2022)。以上研究表明,香榧树体结构调整一方面通过提高香榧树体对光能的利用效率,增强了叶片光合能力;另一方面通过调节树体内部激素平衡,促进幼果中营养物质的积累与分配,最终提高坐果率。
4 结论
骨干枝数目和骨干枝开张角度调整均能促进香榧幼年林成花坐果。其中,3 骨干枝和4 骨干枝处理明显提高了香榧树冠中层和下层的光照强度,有利于提高树体光能利用率,提高叶片净光合速率,促进坐果。开张角度60°处理可以通过降低树体极性,调节混合芽内源激素含量及不同激素间平衡,调整光合碳同化物运输分配,影响C/N 比等促进成花坐果,但开张角度过大(90°)则会抑制成花坐果。综合分析,3 骨干枝和4 骨干枝与开张角度60°的交互处理对促进香榧幼年林成花坐果的效果最佳。
猜你喜欢
恋爱婚姻家庭(2023年1期)2023-02-15
金山(2021年10期)2021-11-02
今日农业(2021年10期)2021-07-28
河北果树(2020年4期)2020-11-26
热带生物学报(2019年1期)2019-04-18
基层中医药(2018年8期)2018-11-10
浙江林业科技(2018年6期)2018-03-13
现代园艺(2017年23期)2018-01-18
绿色中国(2017年19期)2017-05-25
食品工业科技(2014年7期)2014-03-11
