
东北地区主要造林树种幼苗期生物量分配特征与异速生长模型*
2023-12-11 03:00叶尔江拜克吐尔汉何怀江张春雨赵秀海郝珉辉
林业科学 2023年11期
周 昊 叶尔江·拜克吐尔汉 何怀江 张春雨 赵秀海 郝珉辉
(1.北京林业大学 国家林业和草原局森林经营工程技术研究中心 北京 100083;2.新疆农业大学 林学与风景园林学院 乌鲁木齐 830052;3.吉林省林业科学研究院 长春 130022)
生物量是评价森林生态系统结构和功能的重要指标(罗云建等,2009),其准确测定为揭示生态系统的物质循环和能量流动等提供基础资料,并在预测生态系统生产力、稳定性、量化碳储量和碳汇功能等方面有重要意义(Fournieret al.,2003;Peichlet al.,2007;毛子军,2002)。目前生物量的研究集中在生物量分配和生物量估算两方面。生物量分配是指生物量在植物不同组织、器官或功能单位中的分配特征,反映了植物的生活史策略,是生态学研究的核心问题之一(McCarthyet al.,2007;Hendriket al.,2012),阐明生物量分配特征不仅可以揭示植物结构与功能,同时对气候变化背景下森林生态系统的碳循环研究起着基础性作用;生物量估算是指对区域内个体林木或整个林分的生物质量进行测算,是量化森林碳储量和生产力,预测森林生态系统功能的重要基础(Duncansonet al.,2015)。生物量估算的主要方法有直接法和间接法,其中直接法需对目标树种全株取样,准确性较高,但需耗费大量时间和精力,不适于大面积森林的生物量测定(Bond-Lambertyet al.,2002;唐守正等,2000);间接法则是通过建立生物量模型,使用少量样本,借助相对易测的指标(如胸径、树高等)对个体生物量进行量化,进而预测整个林分的生物量和生产力,是目前应用最广泛的生物量估算方法(Hossainet al.,2016;Ubuyet al.,2018;侯燕南等,2016)。总体而言,生物量分配体现的是植物在生长发育过程中不同组织、器官或功能单位对能量的累积,而生物量估算则主要描述了植物在个体或林分水平上对能量的累积,两者之间有重要联系。
20 世纪以来,全球范围内开展了大量生物量分配与估算的研究,建立了各种形式的生物量异速生长模型,成为估算区域或全球森林生物量和碳储量的重要工具(Alaback,1986)。然而,根据联合国政府间气候变化专门委员会(IPCC)提供的默认生物量参数,发现这些研究大都基于欧洲或北美的森林类型,对估算我国复杂多样的森林生态系统生物量并不适用(张小全等,2009)。国内有关森林生物量的研究始于20 世纪下半叶,针对我国森林生态系统结构复杂、类型多样的特点,构建了大量适用于不同森林类型的生物量异速生长模型(冯宗炜,1999;李晓娜等,2010;黎燕琼等,2010;费玲等,2016)。由于我国森林类型十分复杂,不同树种或同一树种在不同地区的形态结构、木材密度等存在很大差异,导致其生物量分配特征和生长模型不相同,采用通用方程估算特定林木或林分的生物量会产生较大误差(Roxburghet al.,2016)。目前,我国仍缺乏统一的模型参数使用标准和成熟的模型评价体系。
东北森林带作为我国“两屏三带”生态战略格局的重要组成部分,具有资源集中、碳储量高、生物多样性丰富等特点,不仅是东北地区的生态屏障,同时也在全国社会、经济建设中扮演着重要角色(王春梅等,2003)。近代以来,受到人口增长和过度采伐等因素影响,东北地区森林资源遭受了严重破坏。提高森林质量、恢复生态功能成为目前东北地区森林经营理论与技术研究的核心目标(张会儒等,2019)。作为评价森林生态功能最重要的基础性指标,森林生物量的研究受到了广泛的重视。目前,对东北林区生物量已开展了大量研究工作,针对不同区域林分特点构建了主要树种的生物量方程,如Wang 等(2006)、董利虎等(2015a;2015b;2015c)和何怀江等(2018)以成熟林木为对象,通过胸径、树高、冠幅等指标对生物量进行拟合,建立了特定树种的异速生长方程;张梦弢等(2012)研究了长白山云冷杉林中主要树种的幼树生物量,构建了天然更新状态下5 种幼树的生物量模型;范春楠等(2013)对长白山针阔混交林中14 种幼树,通过分析不同树种各器官及全株生物量,构建了幼树生物量模型。目前,东北地区人工林主要造林树种幼龄时期的各器官生物量分配特征还不够全面,常见树种的生物量模型研究还不够完善,基于特定树种构建的单树种生物量方程往往并不适合区域性生物量估算。此外,以往研究大多集中在地上生物量估算,地下生物量由于取样困难且需耗费大量人力、物力导致相关研究较少,基于单树种和全树种生物量模型来预测人工针阔混交林幼龄期生物量的研究仍相对缺乏。
本研究以东北地区10 种主要造林树种幼龄苗木为研究对象,分析叶片、根系、茎干等器官生物量分配特征,构建单树种和全树种生物量异速生长模型,并筛选最优生物量模型,研究结果以期提高东北林区不同树种个体和区域性生物量的估算精度,为东北地区人工林尤其是幼龄人工林的生物量和碳储量精确评估提供基础支持。
1 研究区概况与研究方法
1.1 研究区概况与样地设置
研究区位于吉林省舒兰市开原林场,地处长白山余脉向松辽平原的过渡地带,属温带大陆性季风气候,年均气温4.3 ℃,其中最热月平均气温(7 月)21.7 ℃,最冷月平均气温(1月)-18.6 ℃,气温年较差较大,年均降水量683 mm,平均相对湿度69%。研究区土壤类型为弱酸性山地暗棕壤,pH 值为5.5~6.5。
造林苗木来自吉林省蛟河林业试验区苗圃和江密峰苗圃的3~4 年生幼苗。2021 年4 月选取生长健康、干形良好的苗木,在退耕还林的缓坡区域建立生物多样性与生态系统功能控制试验样地。参照Tobner 等(2016)的 试 验 设 计:在 样 地 内 设置160 个5 m×5 m 的小样方造林,样方间隔1 m,每个小样方种植苗木81 株,每株间隔0.5 m。选取的10 个造林树种包括:色木槭(Acer mono)、白桦(Betula platyphylla)、水 曲 柳(Fraxinus mandshurica)、 胡 桃 楸(Juglans mandshurica)、怀槐(Maackia amurensis)、红松(Pinus koraiensis)、蒙古栎(Quercus mongolica)、紫椴(Tilia amurensis)、 红 皮 云 杉(Picea koraiensis)、 黄 檗(Phellodendron amurense)。
1.2 标准木取样方法
2021 年7―8 月对全部树种进行标准木取样,每个树种选取15~21 株树干通直、长势良好、无病虫害和机械损伤的个体作为标准木。测量并记录标准木的树高,基径等数据,然后对标准木进行全株挖掘,挖掘过程中确保植株根系的完整,并在不损伤根系的情况下清理根系上残留的土壤。全株取样后,将标准木分解为根系、茎干、叶片等器官,用标签记录树种名称、树高、基径等信息后装入自封袋带回实验室。样品清洗后于75 ℃的恒温烘箱中烘干至恒质量,再测定样品干质量。
1.3 生物量分配特征与异速生长模型构建
采取一元线性、二次函数、幂函数等形式的方程,对10 个树种的树高,基径与各组分生物量建立回归关系,共建立14 种方程(表1)。由于生物量模型存在异方差,因此对数据进行自然对数转化消除异方差后建立模型,转化形式为表1 方程Eq.1—Eq.4。
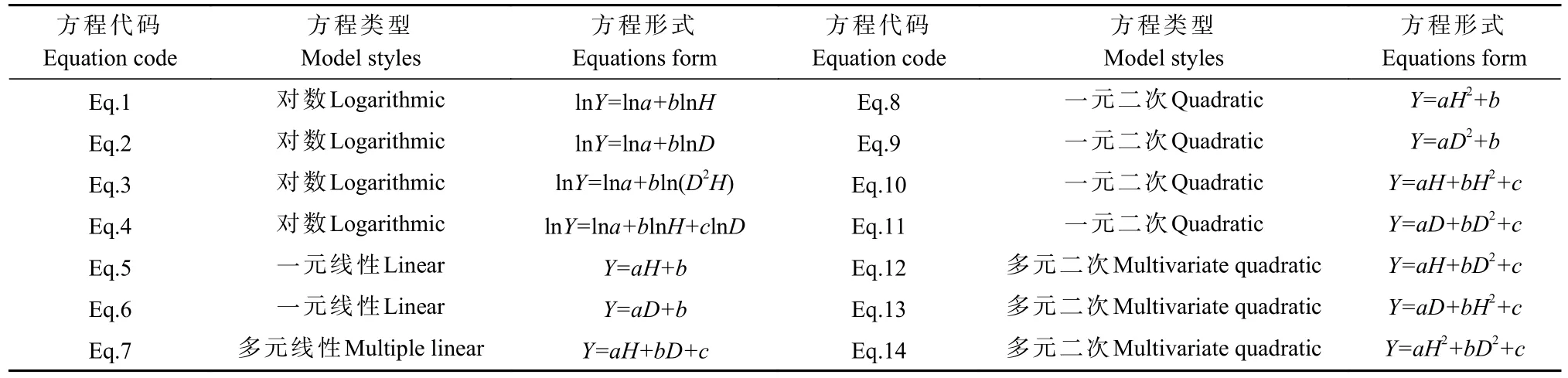
表1 生物量方程形式①Tab.1 Form of biomass equation
采用模型决定系数R2、参数的显著性(P<0.05)、均方根误差(RMSE)、赤池信息标准(AIC)等指标评价模型的拟合优度。由于通过自然对数对数据进行转化时会引入系统误差,因此在利用对数模型进行生物量拟合时,为确保拟合结果真实可靠,通过校正因子(CF)对模型进行校正处理,以消除系统误差。本研究所有数据分析均在统计软件R(4.0.3)中进行。
2 结果与分析
2.1 不同器官生物量分配特征
10 个树种的茎干、叶片和根系平均生物量占比分别为45.0%±10%、19.5%±10%和35.5%±12%,呈现为茎干生物量比例较高,叶片生物量较低的趋势(图1)。不同树种生物量器官分配比例不同,水曲柳、白桦等树种的茎干生物量占比最大(41%~62%);怀槐、蒙古栎等树种的根系生物量占比最大(50%~55%);红松、红皮云杉2 种针叶树的叶片生物量占比最大(31%~51%)。
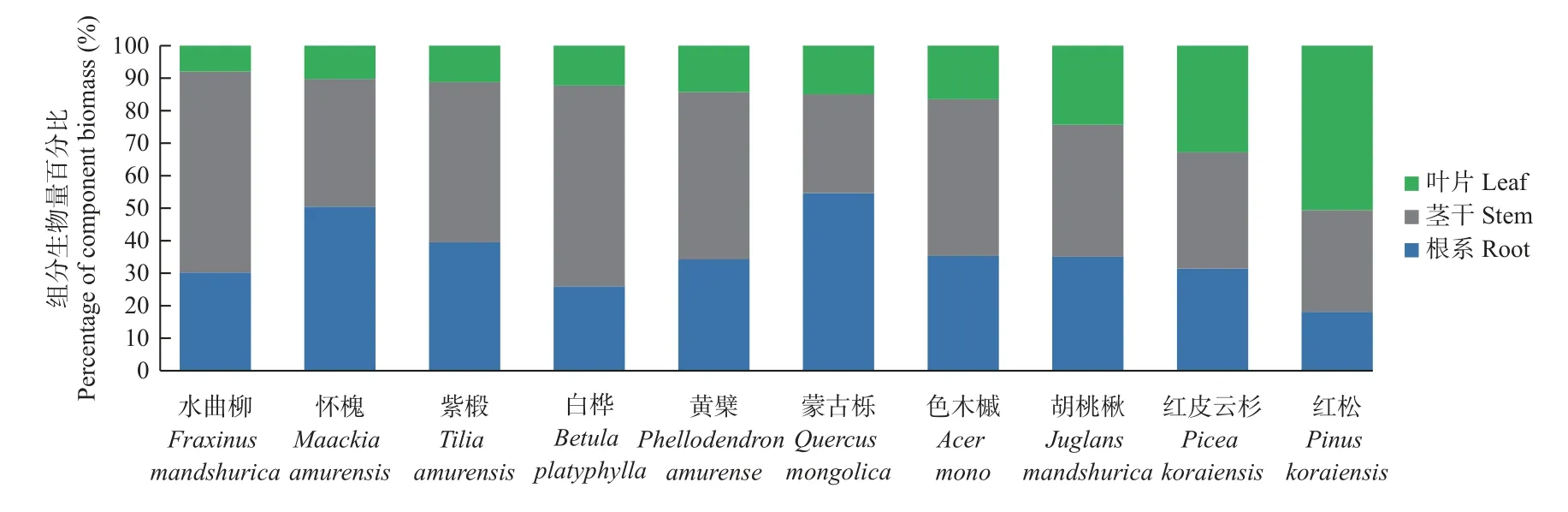
图1 不同树种各组分生物量分配格局Fig.1 Biomass distribution pattern of various components in different tree species
10 个树种各器官生物量随基径变化有所差异。随基径增加,叶片生物量分配比例呈下降趋势,茎干生物量分配比例呈上升趋势,根系生物量分配比例变化不明显,说明多数树种根系生物量在林木生长过程中较稳定(图2)。
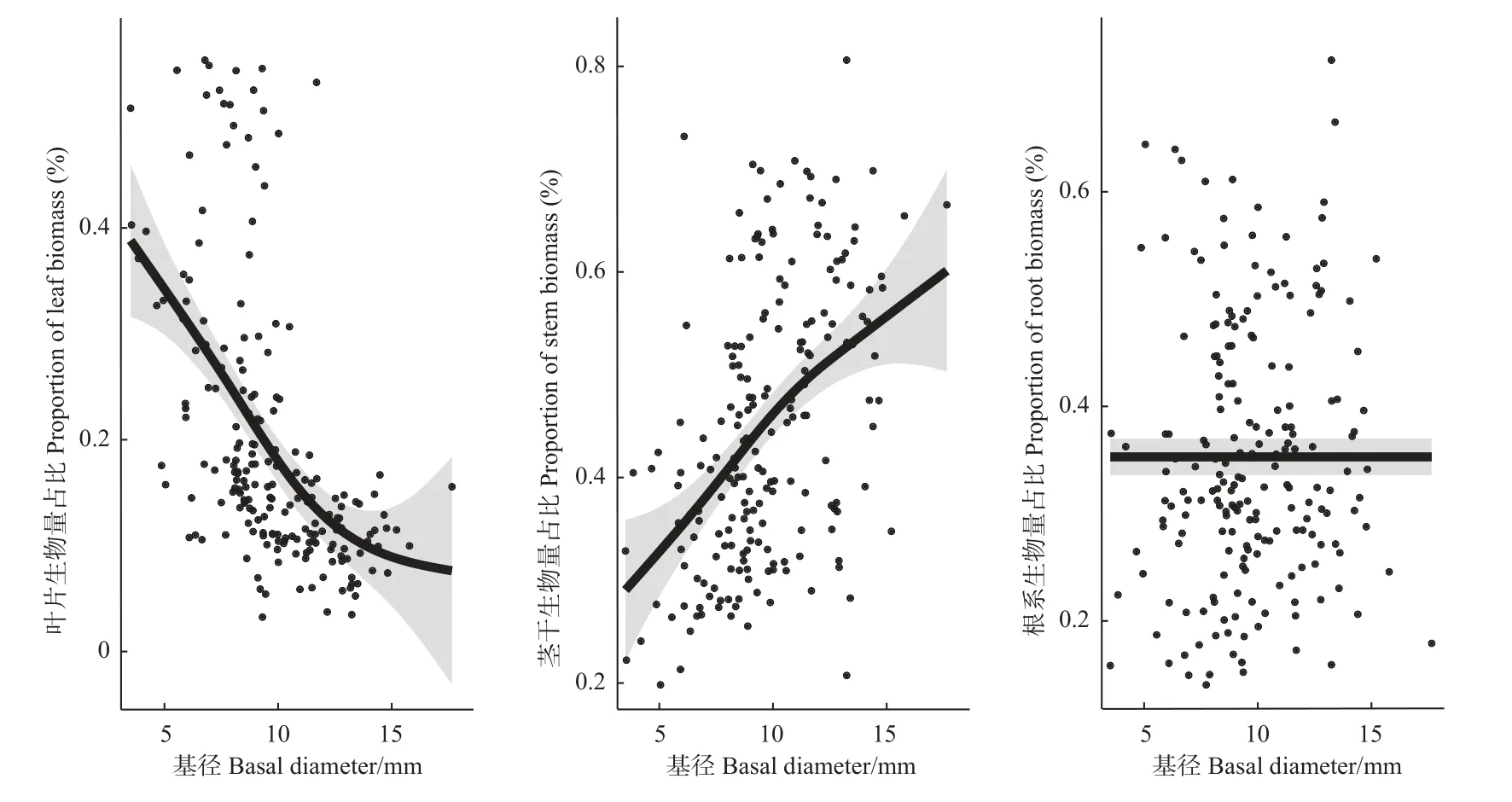
图2 各器官生物量分配比例随基径的变化趋势Fig.2 Variation of biomass distribution in each organ with base diameter
2.2 异速生长方程的构建与筛选
样地内10 个树种的叶片、茎干、地上、根系及整株生物量随树高和基径的变化趋势分别如图3、图4所示。
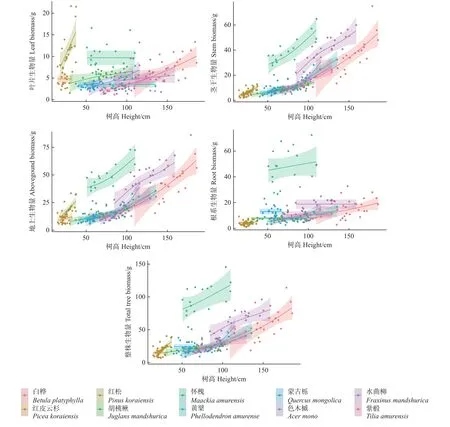
图3 各组分生物量随树高的变化趋势Fig.3 Variation trend of biomass distribution in different component with tree height
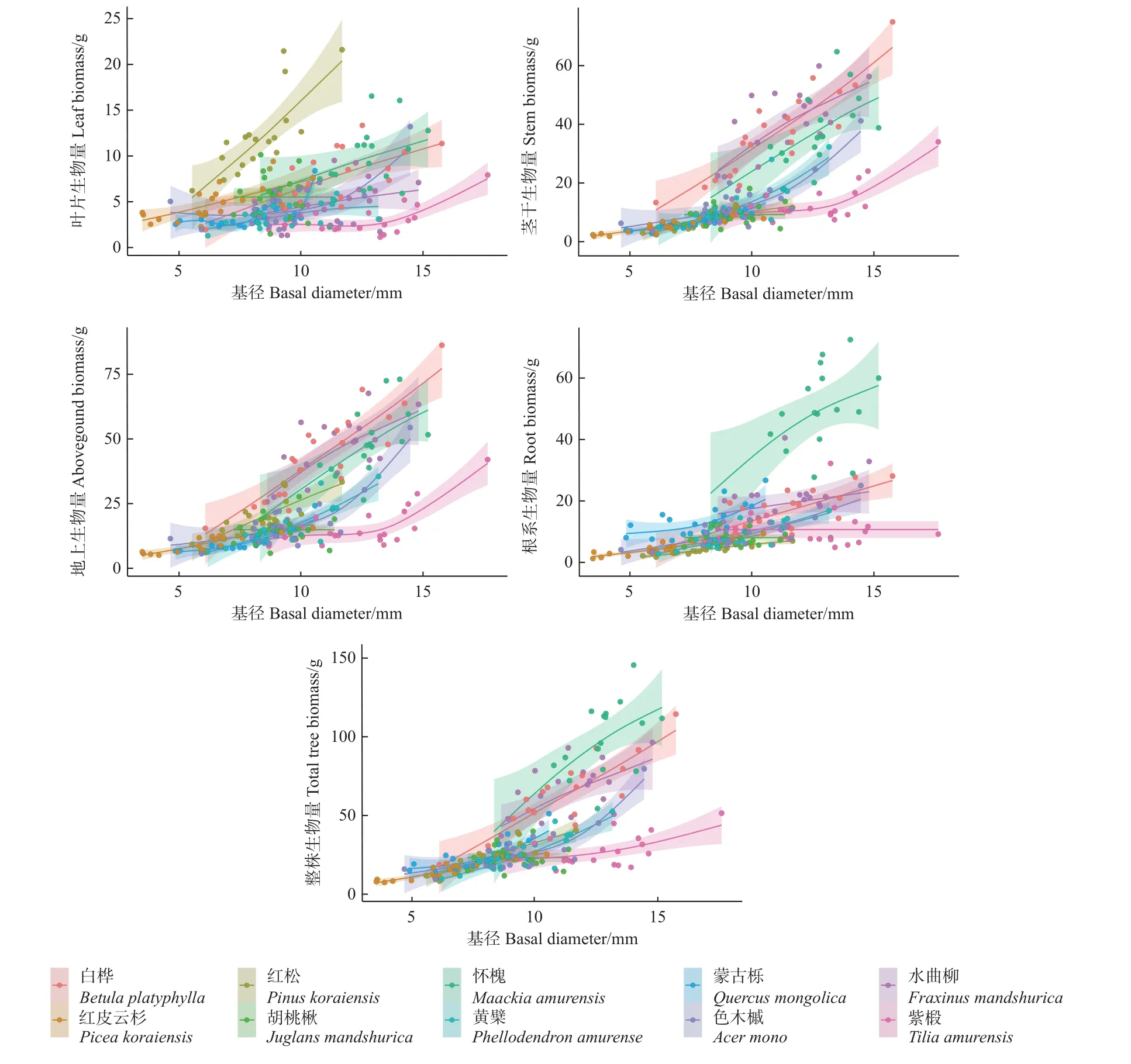
图4 各组分生物量随基径的变化趋势Fig.4 Variation trend of biomass distribution in different component with base area
以基径和树高为解释变量,整株、地上、根系、茎干、叶片生物量5 个部分的自然对数为响应变量,采用14 种形式的方程进行拟合,构建10 种幼树单树种和全树种的生物量方程,共拟合方程700 个。采用解释能力(R2)高、AIC 值小、方程形式简洁作为生物量方程筛选标准,通过计算幂函数、线性函数、二次函数和多元函数的R2和AIC 值筛选最优方程形式,综合所有单树种和全树种生物量方程的拟合结果来看,R2值以幂函数和多元函数最高,二次函数最低;AIC值为幂函数优于其他3 种形式函数,因此,确定幂函数为最佳拟合形式。限于篇幅,本文仅呈现幂函数的单树种和全树种最优生物量方程(表2)。
对茎干和地上部分的拟合效果整体优于叶和根系部分。对单树种最优生物量模型而言,以基径(D)或基径-树高组合(D2H)作为解释变量的拟合效果较好。对全部树种而言,整株、叶片、地上、根系部分均以树高、基径(D,H)分别作为2 个解释变量的形式为最优模型,而茎干部分则以基径-树高组合(D2H)为解释变量的形式为最优模型。对比不同解释变量的最优生物量模型可以发现,树高对生物量的整体拟合效果低于基径,树高(H)作为第2 解释变量加入模型后方程解释能力有所提高。对比各树种不同器官的最优生物量模型形式可以发现,整株、地上、茎干部分采用最多的形式为Y=a(D2H)b,根系使用次数最多的形式为Y=aDb,表明根系生物量与基径关系密切且随树高变化不显著。
为进一步说明单树种生物量模型和全树种生物量模型的适用性,采用单样本t检验比较了生物量最优模型的平均拟合优度(R2),表明单树种生物量模型的叶片、根系、茎干以及地上部分显著优于全树种生物量模型;但对整株生物量而言,单树种生物量模型与全树种生物量模型差异不明显(表3)。
3 讨论
3.1 生物量分配特征
如何有效分配生物量对林木个体的生存和繁殖有重要意义,植物可通过选择不同生物量分配模式以适应复杂多变的环境,从而最大效率地利用资源,提高其生存适合度与竞争能力(Xieet al.,2016;孙越等,2017)。根据平衡生长假说,为更好适应生境,当资源压力较大时,植物会优先将生物量分配给需要的组织或器官。一般情况下,喜光树种更倾向于将生物量较多地分配给茎干以快速占据林分上层空间,而耐阴树种则更倾向于将较多生物量分配给枝叶部分以增加树冠面积从而获取更多光照,实现最大速率的生长(赵厚本等,2022)。本研究所选的10 个树种表现出不同生物量分配策略:当处于幼龄期时,白桦、水曲柳、黄檗等阔叶树种将更多生物量分配给茎干从而实现较快的高生长,因此这类树种的茎干所占生物量最大,平均树高显著高于其他树种;红皮云杉、红松这类针叶树种相较于阔叶树种有着更高的叶生物量分配比例,这是由于红皮云杉和红松在发育初期高生长较为缓慢,对资源的竞争能力低于阔叶树种,更多的生物量会分配给叶片,有助于获取更多的光照、增强个体的生存能力。怀槐属于固氮树种,根部具有根瘤,喜肥沃湿润的土壤,发达的根系有利于提高植物的抗逆性和固氮能力;蒙古栎是营造防风林、防火林、水源涵养林的主要树种,自身具有耐贫瘠、耐干旱、萌生能力强的特点,发达的根系有利于提高对土壤养分的竞争能力和对环境胁迫的抗逆性(王淼等,2004)。由此可见,植物的生物量分配特征与生活史策略与其生态学特性密切相关。此外,在生长发育过程中,植物为适应环境变化和不同阶段的竞争需求,会调整自身生物量的分配策略。本研究结果显示:随着基径增加,茎干生物量分配比例呈上升趋势,而叶片生物量分配比例呈下降趋势,根系生物量的变化则不明显,该结果基本符合植物的平衡生长假说,即当资源稀缺时,生物量会优先分配给更需要的器官,当该器官获得收益时,其他器官发育将被延缓(Weiner,2004),这一生存策略有利于在竞争激烈的环境中提高自身竞争能力(赵厚本等,2022)。
3.2 生物量异速生长方程分析
利用树高、胸径或其他解释变量与生物量的回归关系构建异速生长模型,是估算林木和林分生物量简便有效的方法。生物量异速生长方程一般以幂函数形式为主,并且多数以树高和胸径分别作为解释变量或以树高—胸径组合作为解释变量(李晓娜等,2010;何怀江,2018)。本试验以东北地区10 种主要造林树种的幼苗为研究对象,构建并筛选了最优单树种和全树种生物量模型,为估算东北地区主要造林树种幼龄阶段的生物量提供了可靠的模型参考。拟合结果显示,幂函数的解释能力优于一元线性、二次函数、多元函数等其他形式的模型,因此,本研究最终结果均采用幂函数形式,并且单树种生物量模型以基径-树高组合作为单一解释变量和以基径作为解释变量的形式为最优模型;全树种生物量模型以基径、树高分别作为解释变量和基径-树高组合作为解释变量的模型为最优模型,这与前人研究结果基本一致(Wang,2006;代海军等,2013;何怀江,2018)。本研究以基径为单一解释变量和基径-树高组合为单一解释变量的模型均具有较高的R2和较低的AIC 值,说明对本地区幼苗、幼树而言,基径与生物量有着较强的内在联系。结合杨瑞等(2009)、范春楠等(2013)对幼龄林木生物量的研究发现,基径作为易被精准测量的指标,在生物量模型中广泛使用,并且以基径作为解释变量的生物量模型解释能力较高,是估算生物量的关键变量。构建的树高-生物量模型拟合优度整体低于基径-生物量模型,表明树高对生物量的影响低于基径,实际应用中树高通常作为第二参数加入生物量模型中以提高模型精度(赵厚本等,2022),本研究通过树高与基径相结合,以(D2H)为解释变量的茎干、地上和整株生物量方程都具有较高的解释能力和拟合优度。
以往生物量模型研究中,多数集中在地上部分生物量,对地下部分生物量研究较少(Bond-Lambertyet al.,2002;Ciencialaet al.,2006;代海军等,2013),其中最主要的原因是林木个体较大,根系的取样和清洗难度高,无法保证研究结论的可靠性,并且取样对森林环境破坏较严重(Chamberset al.,2001;宇万太等,2001;董点等,2014)。本研究以幼龄林为研究对象,根系取样工作量相对成熟木较少,且取样精度高,因此能很好的拟合林木地下生物量方程,弥补前人研究的不足。
3.3 单树种生物量模型和全树种生物量模型对比
单树种生物量模型以基径(D)和基径-树高(D2H)组合作为单一变量的形式最优。全树种生物量模型以树高和基径(D,H)分别作为2 个独立变量和以基径-树高(D2H)组合作为单一变量的形式最优。采用单样本t检验分析对比单树种和全树种的各器官生物量最优模型后发现,单树种生物量最优模型的根系、茎干、叶片以及地上部分显著优于全树种最优模型(P< 0.05),而整株生物量则没有显著差异。因此,在估算特定树种的各器官生物量时,单树种生物量模型比全树种生物量模型有更好的解释能力。全树种模型采用的解释变量较多,考虑的因素较为全面,在预测区域尺度的生物量时比单树种模型有更广泛的适用性,因此一般采用全树种模型估算大尺度范围的生物量(Wanget al.,2002;Chaveet al.,2005;林力,2011)。
10 个主要树种各器官生物量最优模型均具较高的解释能力,能很好估算各器官生物量,但对某些树种(水曲柳、胡桃楸和紫椴等)的根系生物量方程解释能力较低。这主要是由于水曲柳、胡桃楸和紫椴的根系十分发达,在前期植苗造林过程中,为减少窝根、保证成活率,对其部分个体进行了剪根处理,在标准木取样时此部分苗木的根系可能尚未完全恢复,因而影响了根系生物量方程的拟合精度。
3.4 树种最优异速生长模型选取
大部分单树种生物量方程的预测能力优于全树种生物量方程,所以在选择生物量方程时应尽量采用对应树种的特定方程(何怀江,2018)。当该树种未能拟合合适的生物量方程或解释能力较低时,可以考虑采用同属植物的生物量模型或全树种生物量模型代替。在调查区域性生物量时,可以采用全树种生物量模型对该区域生物量估算(Wanget al.,2002;Chaveet al.,2005;林力,2011),全树种生物量方程采用的解释变量较多,比单树种生物量方程有更好的解释能力和更广泛的适应性。但模型解释变量越多,调查取样难度就会变大,易产生数据误差。由于本研究树种处于幼龄期,树高易测量,但实际应用中考虑到方程使用的简便性和实用性,最优模型选取应在各项参数检验合格的情况下,选取解释能力高和形式简便的方程。
4 结论
本研究采用全株取样法,分析了东北地区10 个主要造林树种的各器官生物量分配特征,建立并筛选了单树种和全树种的生物量最优模型。总体而言,本研究所筛选的生物量最优模型均采用了幂函数形式,单树种生物量模型拟合精度较高,适用于特定树种生物量的精确估算,而全树种生物量模型对区域性生物量估算有更广泛适用性。本研究结果能够为预测东北地区的森林生物量尤其是幼龄林生物量提供可靠的模型参考。
猜你喜欢
南京林业大学学报(自然科学版)(2022年1期)2022-11-29
今日农业(2020年19期)2020-12-14
农业机械学报(2020年3期)2020-04-01
四川林业科技(2019年5期)2019-07-07
种子(2019年5期)2019-07-02
安徽农学通报(2019年1期)2019-02-14
湖北林业科技(2019年6期)2019-01-06
湖北林业科技(2017年2期)2017-05-25
天然产物研究与开发(2016年11期)2016-06-15
中国科技信息(2015年2期)2015-11-16
