
基于潜热效应的活立木冻融检测传感器设计与实验
2020-04-01 08:11:10赵燕东
农业机械学报 2020年3期
田 昊 高 超 赵 玥,3 郑 焱,4 赵燕东,4
(1.北京林业大学工学院,北京 100083;2.北京工商大学计算机与信息工程学院,北京 100048;3.城乡生态环境北京实验室,北京 100083; 4.林业装备与自动化国家林业局重点实验室,北京 100083)
0 引言
植物冻融特性对于植物的存活及生长具有重要作用,是评价植物抗冻能力及进行植物选育的重要依据[1-3]。植物冻融时会引起植物组织结冰,产生栓塞,进而导致木质部导管内气体和水分含量的变化及细胞间气体水分含量的变化。同时,冻融过程中存在热能交换,在结冰及融化时伴随有放热及吸热现象[4]。国内外学者对冻融发生的机理已经开展了广泛的研究,相关研究表明,冻融过程中冰晶产生的位置决定细胞是否死亡[5],细胞内冰晶会导致细胞死亡[6-9],而细胞外冰晶可以暂时保护细胞[5,10]。因此,在植物冻融过程中检测植物体内冰晶的形成及变化具有重要意义。早期研究中,主要通过采集植物样本、在实验室进行解剖、观察植物组织的内部结构变化、分析植物组织的生化指标来判断植物的冻融情况,这种方法存在滞后性,难以实时检测[11-13]。随着科学技术的发展,红外光谱法、核磁共振成像法、阻抗图谱法等被用于植物冻融检测[14-16],这些方法具有测量快速、图像精度高、便于直观分析的优点,但是对于农林业生产实际来说,难以实现实时检测,并且设备价格昂贵。安装简便、价格低廉、可以实时在线检测的植物冻融检测装置成为一种迫切的需求。KLEPPER等[17]研究了一种LVDT传感器,通过无损检测茎干直径的变化实时监测植物的冻融情况,LVDT传感器在植物冻融研究中得到了广泛使用[18-21]。SEVANTO等在2012年研究了一种Ultrasonic Emissions传感器,用于监测植物冻融过程中的空化。文献[22-25]基于介电原理,在植物冻融监测上开展了相关传感器的研究,通过检测植物茎干的水分变化分析茎干体积含冰量变化,实现对植物冻融变化的分析。但是,植物冻融变化时,不仅冰水含量的变化影响植物体内水分的占比,植物自身生命活动也会影响植物体内水分含量,基于植物水分研究植物含冰量及冻融变化存在一定误差。因此,本文基于冻融过程中的潜热效应[26],设计一款活立木冻融检测传感器,以实现同步实时监测植物冻融过程中体积含水率及热量变化,基于潜热效应判断植物冻融过程中的吸热点和放热点,并在此基础上,结合植物体积含水率变化计算植物体积含冰量及冻融深度,对植物冻融情况进行分析。
1 传感器设计
1.1 传感器测量原理
潜热是物质从一个相变化到另一个相吸收或放出的热量,这是物质在固、液、气三相之间以及不同的固相之间相互转变时具有的特点之一。植物在冻融过程中体内水分存在液态水与固态冰之间的相互转化,因此必然伴随潜热效应的发生,基于潜热效应进行温度监测,判断植物冻融期间的热量交换及冻融点,同时基于驻波参量同步测量活立木茎干体积含水率变化,进而实现植物冻融的准确判断,并根据植物体内体积含水率计算植物冻融期间植物体内体积含冰量的变化,避免植物自身生命活动引起体积含水率变化而造成的计算误差,测量原理如图1所示。

图1 传感器测量原理图
当植物茎干发生冻融时,茎干内的水分发生固液转换,潜热效应引起茎干温度的变化,进而导致插入茎干内部的铂电阻温度的变化,铂电阻的内阻会随温度线性变化,恒流源向铂电阻的两端提供稳定幅值的电流,通过电压放大器观察铂电阻两端的电压,即可等效得到铂电阻的内阻,进而计算得到铂电阻的温度,铂电阻测温等效电路如图2所示。其中U1、U2为放大器。
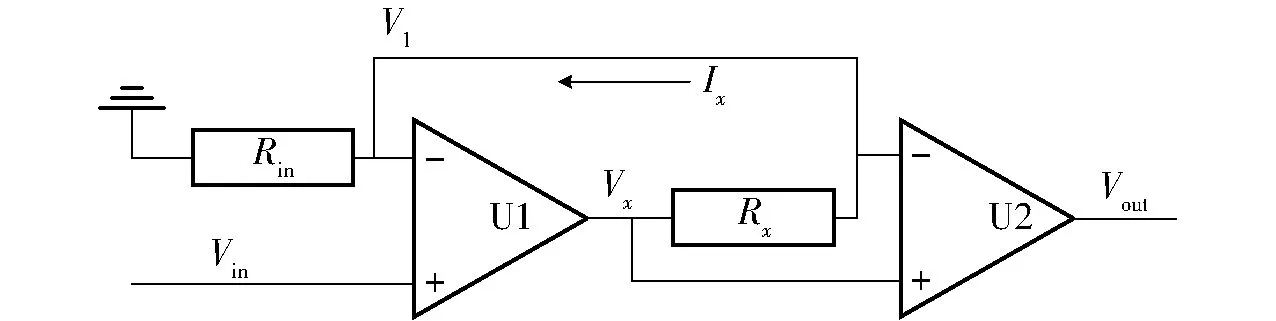
图2 铂电阻测温电路示意图
根据运算放大器的“虚短”原理可知
Vx=A1(Vin-V1)=A1(Vin-IxRin)
(1)
式中Vx、Vin、V1——电路各点电压
Rin——输入电阻
Ix——流过铂电阻的电流
A1——U1的开环放大倍数
当A1=+∞时,式(1)可化简为
Ix=Vin/Rin
(2)
运放U2输出的电压为
Vout=A2RxIx
(3)
式中Vout——运放U2输出的电压
Rx——铂电阻的电阻
A2——U2的运算放大倍数
而铂电阻温度与电阻的关系为
T=KRx+B
(4)
式中T——铂电阻温度
K、B——温度与电阻的关系系数
通过式(2)~(4)可得
(5)
当电路及温度测量范围确定时,式中K、B、Rin、A2和Vin为固定值,因此铂电阻温度T与电压Vout呈线性关系,因此式(5)化简得
(6)
其中K′=KRin/(A2Vin),B′=B,经过标定后即可得到K′与B′,通过测量运放U2输出的电压Vout并将其代入式(6)即可计算得茎干温度。
基于潜热效应,通过检测植物茎干温度变化可以得到植物茎干发生冻融的时刻,进而实现植物冻融的准确判断,由于水和冰的介电常数分别为81和3(25℃),当茎干发生冻融时,茎干内冰水比例随之变化,通过检测发生冻融时茎干体积含水率及冻融过程中茎干体积含水率的变化可以计算得到茎干中冰的含量。茎干体积含水率检测如图1所示,当高频信号源产生的电磁波(100 MHz正弦波)沿着同轴传输线传输至环形探头处时,由于茎干中水冰含量的变化会使得茎干介电常数发生变化,进而使得探头处阻抗与传输线阻抗不匹配,电磁波在探头处发生反射,反射波和入射波叠加在一起形成的驻波的电压幅值与探头处阻抗密切相关,而探头处阻抗与茎干介电常数相关[27],因此通过测量同轴传输线的驻波可以得到茎干的体积含水率[22]。茎干体积含水率测量等效电路示意图如图3所示。

图3 茎干体积含水率测量等效电路示意图
同轴传输线两端的电压为
U=2A(ZP-Z)/(ZP+Z)
(7)
式中U——同轴传输线两端的电压
A——同轴传输线两端驻波的电压幅值
ZP——探头阻抗
Z——传输线阻抗
传输线阻抗Z为50 Ω,电压幅值A和传输线阻抗Z为恒定值,U只与ZP相关,而ZP与介电常数相关,因此通过测量U即可表征探针处茎干的介电常数,进而得到茎干体积含水率。
1.2 传感器机械结构
1.2.1探头设计
由于植物的生长,茎干也会逐渐变粗,现有的相关研究中用于茎干体积含水率测量的环形探头安装后,探头的直径便固定,在一段时间生长后探头会对茎干产生压迫并在茎干上形成凹痕,最终会影响测量结果。因此本文提出并设计了新型的环式弹片探头(图4a),探头为环形结构,使用304不锈钢弹片(图4b)制作,钢片厚度为0.2 mm,环形探头可以自动收缩,同时又具有一定的弹性,这样探头在紧密贴合茎干的同时也可以随着茎干变粗而变化,避免对茎干造成压迫。同时在环式弹片探头上设计并安装了铂电阻用于温度同步测量,铂电阻为PT100,直径1 mm,长度可根据安装茎干的直径进行定制,复合设计探头如图4c所示,探头整体由3个环式弹片探头组成,上下两个负极,中间的为正极,负极复合有铂电阻,正极为单独的环式弹片探头,探头整体如图4d所示。

图4 传感器探头
1.2.2硬件电路设计
传感器硬件系统构成如图5所示,主要包括传感器和采集器两部分。采集器部分包括供电单元、分时供电单元、模数转换(Analog to digital converter, ADC)采集单元、时钟单元和数据通信单元;传感器部分包括茎干体积含水率检测单元和茎干温度检测单元。分时供电单元控制茎干体积含水率检测单元和温度检测单元的供电,以避免同步检测时存在干扰引起的测量误差,采集器通过ADC采集单元采集茎干水分检测单元和茎干温度检测单元输出的模拟信号并换算得到对应的茎干体积含水率及温度,并基于茎干冻融时潜热效应引起的温度变化判断吸热和放热点,结合茎干体积含水率计算得到茎干体积含冰量,之后通过时钟单元和数据通信单元将数据按设定的时间进行输出,数据通信单元包含GPRS(General packet radio service)无线传输模块和RS485传输模块,因此数据既可以无线传输,也可以通过有线方式读取,便于不同应用场景的使用。

图5 硬件系统构成框图
1.2.3整体结构设计
传感器整体结构由测量探头、硬件电路和外壳组成。探头的组成如1.2.1节中所述,两个探头环之间间隔为10 mm,通过探头环上的螺纹连接件实现与硬件电路的连接。硬件电路由两层测量电路组成,底层为传感器电路,顶层为采集器电路。外壳使用SolidWorks软件设计并采用3D技术打印制作,材料为光敏树脂,打印公差为0.2%。传感器结构组成及实物如图6所示。

图6 传感器结构组成及实物
2 传感器性能分析
2.1 传感器标定
2.1.1茎干温度检测单元标定
式(6)中K′和B′可以通过理论计算得到,但是电路温漂及焊接会对测量造成影响,因此需要对茎干温度传感器进行标定,消除误差。标定中将传感器放置在GDJ-1500B型高低温交变试验箱内,设置温度从-30~80℃逐渐增加,记录传感器输出电压,同时将DS18B20型温度传感器固定在茎干温度传感器探头处,记录DS18B20型温度传感器测量的温度,使用最小二乘法拟合测量值,结果如图7所示。由图7可看出,拟合决定系数为0.999 6,高于0.99,传感器输出电压与温度具有良好的线性关系,满足使用要求。
2.1.2茎干体积含水率检测单元标定
从活体植株上截取一段茎干,选取的植株为瓜栗,用溢水法测量茎干的体积(106 cm3),然后将传感器安装在茎干上并放入干燥箱中干燥(45℃),每隔6 h取出一次称量并记录传感器的输出电压,直至茎干完全干燥,通过干燥法计算得到对应的体积含水率,对测量结果进行拟合,拟合结果如图8所示,拟合决定系数为0.997 6,高于0.99,说明茎干体积含水率检测单元输出电压与茎干体积含水率具有良好的相关性,可以用于表征茎干体积含水率。

图8 茎干体积含水率检测单元标定曲线
2.2 静态特性分析
2.2.1测量范围
从2.1节中可以看出,温度传感器测量范围为-30~80℃,茎干体积含水率测量范围为0~68.67%。
2.2.2分辨率
分辨率表示传感器能有效辨别的最小示值差,数字仪表分辨率通常决定于ADC的位数精度。本文设计的传感器ADC采样模块的位数为12,分辨率为0.000 8 V,体积含水率从0~68.67%变化时茎干体积含水率检测单元输出电压为0.392~1.750 V,可知茎干体积含水率检测单元输出电压每改变1 V代表体积含水率变化50.57个百分点,茎干体积含水率检测单元输出电压通过ADC采集后输出至单片机,因此传感器体积含水率检测分辨率为0.04%。同理对于茎干温度检测单元,温度-30~80℃变化时茎干温度检测单元输出电压从1.969 V增加至2.905 V,可知茎干温度检测单元输出电压每改变1 V温度变化128.21℃,因此温度传感器的检测分辨率为0.10℃。
2.2.3稳定性
为了测试传感器的稳定性,将传感器探头分别置于空气中(体积含水率默认为0)、水中(体积含水率默认为100%)和安装在湿茎干上(用保鲜膜包裹防止茎干水分流失),在室温下每2 s记录一次传感器输出结果,共记录2 050次,茎干体积含水率测量结果如图9a所示。同时将传感器空载放置在恒温箱中,温度分别设置为-30、22、50℃,每2 s记录一次传感器输出结果,共记录2 050次,温度测量结果如图9b所示。
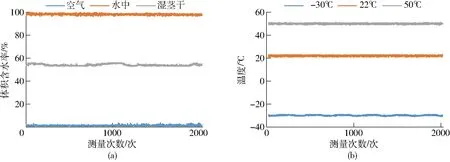
图9 稳定性测试结果
从图9可以看出,在2 050次测量中,传感器输出体积含水率和温度波动很小,满足测量要求,为进一步分析传感器输出稳定性,对2 050次测量结果进行统计分析,结果如表1所示。可以看出,体积含水率标准差小于1%,误差波动小于2%,稳定性满足体积含水率测量要求;实验室用恒温箱控温精度为±1.5℃,而温度测量结果标准差小于1℃,误差波动小于1.5℃,温度波动与恒温箱一致,表明稳定性满足温度测量要求。
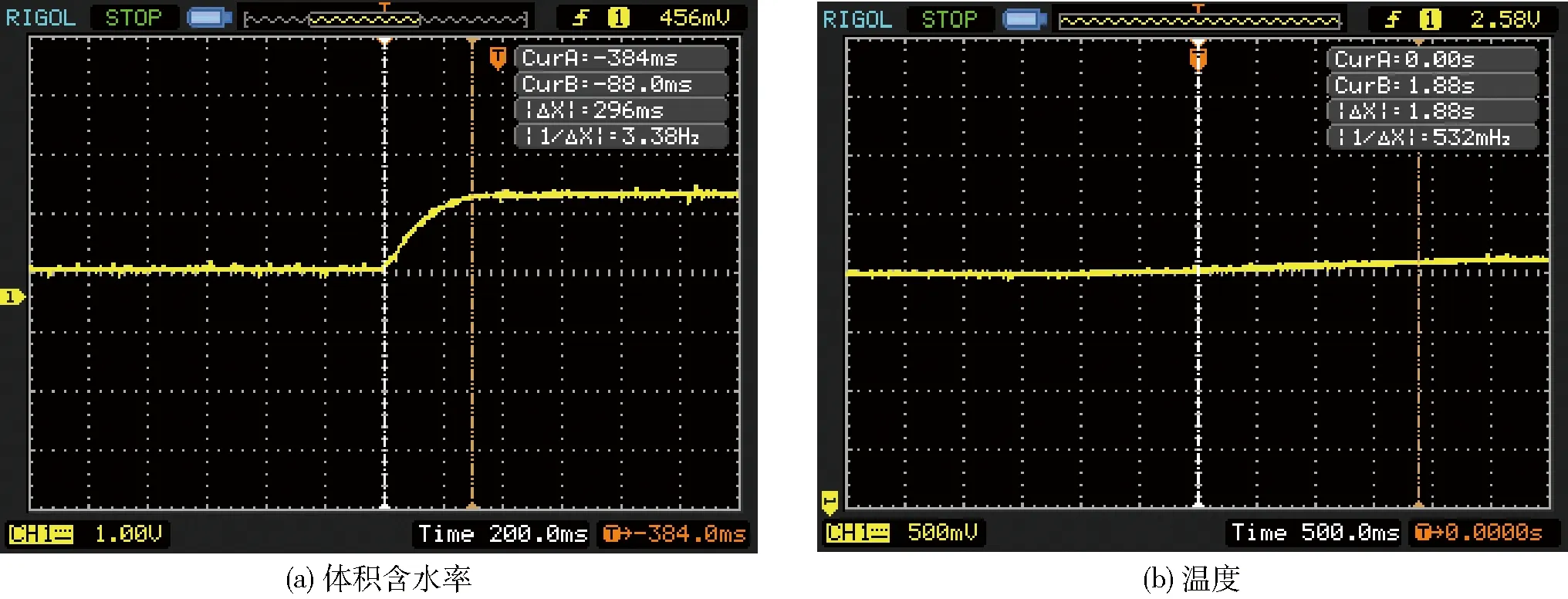
图10 传感器动态特性曲线
2.3 动态特性分析
动态特性指传感器对随时间变化的输入量的响应特性,将传感器探头置于空气中,待传感器输出稳定后,在一个500 mL的烧杯中装满热水,将传感器探头迅速放入水中,将这一过程的温度与体积含水率视作输入信号,此时输入为一阶阶跃信号,通过测量输出随输入的变化得到动态响应特性,用示波器观测传感器输出信号变化,测量结果如图10所示。可以看出,体积含水率的动态响应时间为0.296 s,温度的动态响应时间为1.88 s,动态响应时间均小于2 s。
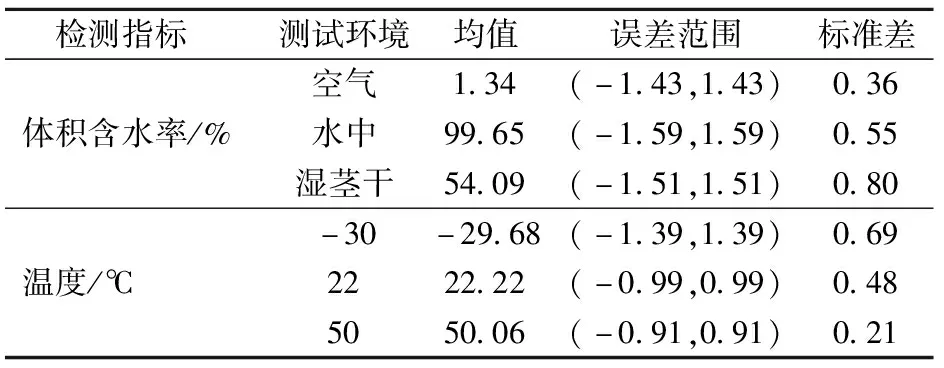
表1 稳定性分析
3 基于潜热效应的冻融检测
3.1 茎干冻融点
液态水和固态冰在转换过程中由于潜热效应会吸收或者放出热量,吸热及放热会影响茎干温度变化速率,通过检测茎干温度及温度变化速率可以对茎干冻融点进行判别。本文选取瓜栗(直径6 cm、株高40 cm)为实验对象,在实验室环境下进行茎干冻融模拟实验,将传感器安装在瓜栗的茎干上,植株放入低温冰箱(DW-40W100型,海尔)中,设置温度为-30℃,当茎干完全冻结后关闭冰箱电源并打开冰箱,让植株自然融化,记录整个冻融过程中冰箱环境温度、茎干温度及茎干体积含水率变化,结果如图11所示。

图11 茎干冻融过程
从图11可以看出,在冻结过程中,随着冰箱中环境温度不断下降,茎干温度也随之下降,当茎干温度下降至A点时,茎干到达结冰点,茎干开始冻结,水由液态转变为固态放出热量,导致茎干温度下降速率明显变缓,同时茎干体积含水率开始下降,随着水完全结冰,放热结束,茎干温度又开始快速下降。而在融化过程中,茎干温度随着冰箱环境温度的上升而上升,当茎干温度上升至B点时,茎干中的水开始融化,水由固态转变为液态吸收热量,导致茎干温度上升速率明显变缓,同时茎干体积含水率开始上升,随着茎干中冰融化完成,吸热停止,茎干温度又开始快速上升。可以看出A和B分别为茎干冻融过程中的冻结点和融化点,由于茎干温度传感器测量探针只插入茎干直径的1/3处,因此在放热和吸热结束时,茎干体积含水率依旧保持下降或上升一段时间后稳定。为了准确判断冻融点,本文提出用茎温变化率Tt表征茎干温度的变化速率,即
(8)
式中 ΔT——茎干温度差
Δt——茎干温度测量时间差
T2、T1——茎干温度
t2、t1——茎干温度T2、T1对应的测量时间点
冻融过程中的茎干温度变化率如图12所示,在茎干放热过程(A到A′)和吸热过程(B到B′)中茎干温度变化率很小(小于0.05℃/min)且茎干温度在-5~0℃之间,因此可以通过检测茎干温度及茎干温度变化率判断植物是否处于冻融过程,发生冻融时的起始时间点即为茎干冻融点,图12中A点为冻结点(10:22),B点为融化点(21:02)。

图12 冻融过程中茎干温度变化率和茎干温度的变化曲线
3.2 茎干体积含冰量
当检测到冻融点时,表明茎干开始结冰或者融化,根据结冰点的茎干体积含水率及冻融期间茎干体积含水率可以计算得到冻融过程中体积含冰量的变化。根据冻融过程中的茎干体积含水率的变化计算得到茎干中冰的质量为
Mice=(θA-θx)Vstρw
(9)
式中θA——结冰点的茎干体积含水率
θx——冻融过程中的茎干体积含水率
Vst——茎干体积
ρw——水的密度
进而可以计算得茎干体积含冰量为
(10)
式中Vice——茎干中冰的体积
ρice——冰的密度
3.3 茎干径向冻融深度

图14 茎干冻融体积含冰量及冻融深度的变化曲线
由于茎干温度传感器测量探针只插入茎干直径的1/3处,因此茎干温度有效探测范围为茎干直径的1/3,从图11中可以看出在传感器检测到放热和吸热完成时,茎干体积含水率依旧保持下降或上升并在一段时间后稳定,说明冻融过程结冰或融化是沿着茎干的径向方向从外向内进行的,越靠近茎干外部,结冰或融化越先发生,因此本文提出基于茎干体积含冰量的茎干径向冻融深度检测,茎干径向冻融模型如图13所示,可得
D=R-r
(11)
Vst=πR2H
(12)
(13)
式中D——冻融过程中冻结茎干或融化茎干的径向深度
R——茎干半径
r——冻融过程中未冻结或未融化部分茎干的半径
H——传感器测量的茎干长度
根据式(10)可得
(14)
进而有
(15)
代入式(11)求得
(16)

图13 茎干径向冻融模型
基于潜热效应通过检测茎干温度及茎干温度变化率可以对茎干冻融过程中的冻结点及融化点进行判断,进而在此基础上实现茎干体积含冰量及径向冻融深度的计算。通过式(10)和式(16)可以得到实验中瓜栗在冻融过程中的茎干体积含冰量及径向冻融深度,结果如图14所示,可以看出茎干中含冰量越高,茎干冻融深度越大。
3.4 野外实验
野外实验在黑龙江省哈尔滨市帽儿山东北林业大学实验林场(127.58°E,45.27°N)进行,将传感器安装在实验林场的香杨树上,如图15所示,香杨树高7.5 m,传感器安装高度为3 m,安装处茎干直径为7 cm,每10 min采集1次数据,监测香杨树从秋天到春天(2018年10月1日—2019年4月20日)的茎干冻融情况,验证传感器在野外长期测量的稳定性和可靠性,测量结果如图16所示。

图16 香杨树的茎干冻融变化曲线
从图16可以看出,从秋天到冬天的过程中,随着环境的变化,植物生命活力下降,香杨树茎干体积含水率逐渐下降,同时茎干温度越来越低,在入冬时节茎干开始出现冻融,植物进入休眠状态,并随着深冬的到来,茎干体积含冰量逐渐上升,茎干径向冻融深度也不断增大;在由冬入春的阶段,茎干开始进入萌芽期,温度升高,植物生命活力逐渐增强,茎干体积含水率开始增大,植物茎干体积含冰量逐渐下降,植物体内冰开始融化,到春末,植物的生命活力恢复,茎干体积含水率恢复到较高水平。同时茎干体积含冰量每天呈单波峰单波谷的周期变化。结果表明本文设计的传感器可以有效检测植物冻融变化,为植物冻融研究提供了一种新的监测手段。
4 结论
(1)设计了一种基于潜热效应的植物冻融检测传感器,可以实现植物茎干冻融过程中茎干体积含水率、茎干温度、茎干体积含冰量及径向冻融深度的测量,为植物冻融监测及研究提供了一种有效的技术手段。基于潜热效应检测植物冻融过程中因吸热和放热引起的温度变化,进而实现植物冻融的有效判断,同时复合茎干体积含水率测量,在对植物冻融有效判断的基础上,根据得到的冻融点,结合茎干体积含水率进行茎干体积含冰量及径向冻融深度的计算。
(2)提出了基于茎干温度及茎干温度变化率的冻融点的检测,通过判断茎干温度的变化范围及茎干温度变化率可以有效识别植物冻融点,并有效消除只根据茎干温度进行判断时由于温度波动异常造成的误差。针对以往茎干体积含水率测量传感器探针存在的缺点,设计了一种环式弹片探头,可以有效避免固定式环形探头长时间安装在茎干上对植物产生压迫而造成凹槽。
(3)标定结果表明,设计的传感器测量结果与真值拟合决定系数高于0.99,通过静态特性及动态特性实验,表明传感器温度和体积含水率测量范围分别为-30~80℃、0~68.67%,分辨率分别为0.10℃、0.04%,传感器稳定性良好,温度测量动态响应时间为1.88 s,体积含水率测量响应时间为0.296 s,动态响应时间均小于2 s,表明传感器具有良好的静动态特性,满足实际使用要求。
(4)通过监测室内瓜栗的冻融变化和室外香杨树从秋到春期间的冻融变化可知,传感器能够有效监测植物冻融过程中茎干体积含水率及温度变化,在冻结过程中茎干体积含冰量增加,径向冻结深度变大;在融化过程中,茎干体积含冰量减小,径向融化深度变大。野外实验表明,长期冻融过程中植物茎干体积含冰量具有单波峰、单波谷的周期变化。说明传感器能够有效监测植物冻融期间水分生理参数的变化,可以作为植物冻融研究的有效监测手段。
猜你喜欢
南京林业大学学报(自然科学版)(2022年1期)2022-11-29 10:10:16
林业机械与木工设备(2022年5期)2022-05-27 09:28:56
林业科学(2021年5期)2021-07-13 07:53:36
中国粉体技术(2021年1期)2021-01-04 02:19:18
种子(2019年5期)2019-07-02 11:37:44
长江科学院院报(2018年12期)2018-12-19 09:52:02
中国计划生育学杂志(2017年3期)2017-06-01 12:10:12
水利科技与经济(2017年2期)2017-04-22 02:34:14
电子制作(2016年1期)2016-11-07 08:42:56
天然产物研究与开发(2016年11期)2016-06-15 20:29:15
