
箭叶淫羊藿种子的休眠类型与萌发研究
2022-12-30 06:58吉浴芳宋松泉田向荣高家东戴嘉兴
广西植物 2022年11期
吉浴芳, 宋松泉, 田向荣, 高家东, 戴嘉兴, 刘 军*
(1. 吉首大学 生物资源与环境科学学院, 湖南 吉首 416000; 2. 广东省农业科学院农业生物基因研究中心, 广东省农作物种质资源保护与利用重点实验室, 广州 510640; 3. 中国科学院植物研究所, 北京 100093 )
种子休眠是活种子在适宜环境条件下暂时不能萌发的一种机制,对种子植物的生存和繁衍起到关键作用 (Gao & Ayele, 2014)。种子休眠是植物对环境适应的一个重要表现 (Donohue et al., 2010; Huang et al., 2010)。高水平的种子休眠会延迟萌发和萌发不整齐,从而导致生长季节的长度缩短和生长不一致;相反,低水平的休眠可能引起种子在有利的生长季节开始之前萌发,存在幼苗死亡的风险 (Donohue et al., 2010),从而降低作物的产量和质量。
休眠是一个复杂的性状,主要由遗传和环境因素决定,植物物种之间存在种子休眠的多样性和普遍性(Baskin & Baskin, 2014)。种子休眠可分为生理休眠 (physiological dormancy, PD)、形态休眠 (morphological dormancy, MD)、形态生理休眠 (morphophysiological dormancy, MPD)、物理休眠 (physical dormancy) 和组合休眠 (combinational dormancy) (Baskin & Baskin, 2004,2014; Iwasaki et al., 2022)。PD存在于具有充分发育和成熟胚的种子中,主要是由种子组分中抑制剂的存在和植物生长物质的缺乏所引起的 (Chen et al., 2010)。PD是最普遍的休眠形式,可分为深度、中度和浅生理休眠,广泛存在于裸子植物和所有主要被子植物中 (Baskin & Baskin, 2004, 2014; Finch-Savage & Leubner-Metzger, 2006)。MD 的种子胚小 (未发育完全),但已分化,种子的萌发不需要释放休眠的预处理,但需要较长时间让胚生长至足够体积,之后萌发。MPD的种子具有未发育完全和生理休眠的胚,其种子萌发需要释放休眠的预处理。MPD的种子胚的生长/胚根突破种皮需要的时间比MD的种子长得多。
植物激素脱落酸 (abscisic acid, ABA) 和赤霉素 (gibberellin, GA) 参与种子休眠和萌发的控制 (Weitbrecht et al., 2011; Iwasaki et al., 2022)。Dong等 (2012) 发现,氟啶酮 (fluridone,Flu, ABA生物合成的抑制剂) 和GA3显著降低莴苣种子萌发的热休眠,而烯唑醇 (diniconazole, ABA分解代谢酶ABA 8′-羟化酶的抑制剂) 和多效唑 (paclobutrazol, GA生物合成途径的抑制剂) 增加莴苣种子萌发的热休眠。在降低莴苣种子萌发的热休眠中,多效唑具有明显拮抗氟啶酮的作用,而氟啶酮具有显著增加GA3的作用。特别是ABA和GA水平之间的平衡及其各自的信号通路在诱导和维持休眠以及促进萌发的调控中起重要作用 (Finkelstein et al., 2008; Graeber et al., 2012; 宋松泉等, 2020a, b)。此外,种子休眠还被一些环境因素释放,如后熟、变温、冷/暖层积、交替光照等,具体取决于物种 (Baskin & Baskin, 2004, 2014; Black et al., 2006; Finch-Savage & Leubner-Metzger, 2006)。
淫羊藿属 (EpimediumL. ) 植物为小檗科 (Berberidaceae) 多年生草本植物,该属植物约55 种,我国有45 种,其中淫羊藿 (Epimediumbrevicornum)、箭叶淫羊藿 (E.sagittatum)、柔毛淫羊藿 (E.pubescens)、朝鲜淫羊藿 (E.koreanum) 和巫山淫羊藿 (E.wushanense) 为《中华人民共和国药典》所收载 (国家药典委员会,2020) 。淫羊藿为常用中药,最早记载于《神农本草经》,具有补肾壮阳、祛风除湿之功效;在临床上,淫羊藿常被用于治疗骨质疏松、更年期综合征、乳房肿块、高血压、冠心病等;另外,淫羊藿还具有增强免疫力、抗衰老、抗肿瘤、抗艾滋病等作用。淫羊藿含有多种生理活性很强的成分,主要包括黄酮类化合物、生物碱、多糖、木脂素和一些必需的微量元素 (Kang et al., 2012; 李聪聪等, 2020)。长期以来,种子萌发困难,淫羊藿的栽培大多采用挖取野生植株进行分蔸种植,但其繁殖系数低,并对野生资源造成了严重破坏。近年来,随着市场需求的大量增加,淫羊藿野生资源的蕴藏量逐年锐减,濒临枯竭。因此,利用种子进行种苗繁育已成为淫羊藿产业化栽培中亟须解决的关键问题。
淫羊藿属植物在果实成熟时,种胚发育不完全,需要经过一段时间的冷层积或冷暖层积才能萌发,具有明显的休眠现象 (樊家乙等, 2010; 付长珍等, 2012; 田向荣等, 2015; 苏贺等, 2016; Rhie & Lee, 2020)。GA3处理促进巫山淫羊藿 (苏贺等, 2016) 和朝鲜淫羊藿 (Rhie & Lee, 2020) 种胚的生长,氟啶酮处理能显著增加巫山淫羊藿种子的萌发率 (苏贺等, 2016)。田向荣等 (2015) 利用常规萌发试验以及直接测量和石蜡制片技术初步研究了箭叶淫羊藿种子的萌发特性。目前,箭叶淫羊藿种子的休眠和萌发特性仍不够清楚。本文以成熟的箭叶淫羊藿种子为材料,研究了种子对水分的吸收、种子的脱水耐性以及温度、层积和植物激素对种子萌发的影响,试图探明箭叶淫羊藿种子的休眠类型及其释放休眠和促进萌发的适宜方法,为产业化育苗提供参考。
1 材料与方法
1.1 材料
箭叶淫羊藿 (Epimediumsagittatum) 的成熟果实于2020年6月采自张家界八大公山 (110°07′ E、29°77′ N,海拔1 302 m),年平均温度小于9.9 ℃,最冷月为1月,平均温度3.5~4.3 ℃,最热月为7月,平均温度26.3~28 ℃;年均降雨量2 300 mm,春夏季 (4—6月) 降雨最多,约占48%。将收集后的果实置于(25±3)℃和(50±5)%相对湿度下自然干燥,待种子从果荚中脱落后,干燥种子含水量到(9.7±0.49)%,一部分种子用于耐脱水性分析,另一部分种子在4 ℃中贮藏备用。
玉米‘郑单958’ (Zeaways‘Zhengdan 958’) 和水稻‘日本晴’ (Oryzasativa‘japonica’) 种子由广东省农业科学院农业生物基因中心种质资源实验室提供。
1.2 耐脱水性分析
将含水量为9.7%的种子置于筛网中;将筛网置于盛有干燥硅胶的干燥器中,分别脱水0、2、4、8、12、24、36 h后,一部分种子用于测定含水量,另一部分种子则在4 ℃下层积60 d后在10 ℃和黑暗条件下萌发。
1.3 含水量测定
种子含水量的测定参照国际种子检验协会的方法 (International Seed Testing Association, 1999) ,以鲜重的百分比表示。
1.4 种子对水分的吸收
箭叶淫羊藿种子、玉米种子和水稻种子分别播种在垫有两层滤纸的培养皿中,加入7 mL 蒸馏水,于25 ℃和黑暗中分别吸胀不同时间后测定种子的含水量。种子的含水量用鲜重百分比来表示。
1.5 层积处理
层积基质为珍珠岩。首先,将珍珠岩在自来水中除去粉尘,121 ℃中灭菌30 min,冷却后装入自封袋中并加入适量的无菌水,室温下放置20 min后,用吸管将多余的水吸掉。其次,将含水量为9.7%的箭叶淫羊藿种子与湿润的珍珠岩 (V/V=1∶10) 在自封袋中充分混合,然后,分别在4、10、(4+10)、(10+4)、(4+10+4) ℃和黑暗条件下层积。最后,层积不同的时间组合后,取样。一部分种子利用OLYMPUS (SZ61) 体式显微镜观察胚的形态变化,拍照、计算胚/种子(embryo/seed,E/S)的比例 (胚的长度/种子的长度);另一部分种子做萌发试验,测定种子的萌发速率和萌发率。
1.6 种子萌发
经不同处理的种子用0.1% (V/V) NaClO灭菌15 min后,播种在垫有两层滤纸的培养皿中,加入6 mL 蒸馏水或不同浓度的植物激素GA3或氟啶酮,之后在注明温度和黑暗的条件下萌发。以胚根突破种皮2 mm作为萌发的标准。
1.7 统计分析
所有数据均使用SPSS 20.0软件的单因素方差分析模型 (one-way ANOVA model) 进行分析,平均数差异的显著性利用Student-Newman-Keuls (S-N-K)检验确定(P=0.05)。
2 结果与分析
2.1 种子对水分的吸收
为了解种子是否具有物理休眠,将收获的新鲜箭叶淫羊藿、玉米和水稻种子在25 ℃和黑暗中分别吸胀0~72 h。我们观察到,箭叶淫羊藿种子具有2个阶段的水分吸收,即阶段Ⅰ (0~4 h) 的快速吸水和阶段Ⅱ(4~72 h)的缓慢吸水直到平台期。在阶段Ⅰ,种子的含水量从9.7%增加到52%;在阶段Ⅱ,种子的含水量从52%增加到56.7%。玉米和水稻种子的吸水过程可分为3个阶段,即阶段Ⅰ的快速吸水和阶段Ⅱ的缓慢吸水以及阶段Ⅲ种子萌发后的迅速吸水 (图1:A)。
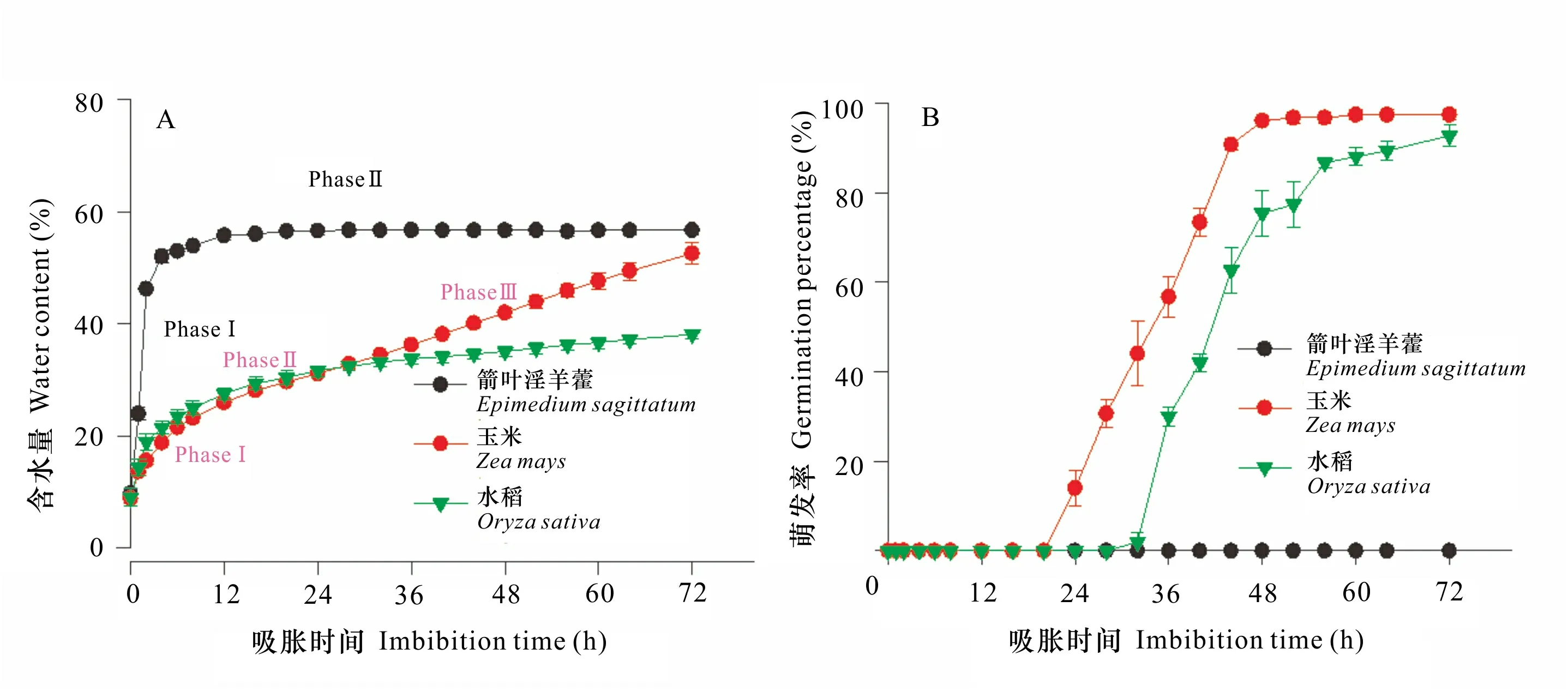
干种子在25 ℃和黑暗中吸胀不同时间后,种子的含水量和萌发率被测定。胚根伸出种皮2 mm作为种子完成萌发的标准。所有的数值均为100粒种子重复3次的平均值±标准误。Water content and germination of seeds were determined after dry seeds were imbibed at 25 ℃ and darkness for different time. A radicle length of 2 mm was used as the criterion for germination completion. All values were of three replicates for 100 seeds each.图 1 箭叶淫羊藿、玉米和水稻种子的含水量 (A) 与萌发率 (B)Fig. 1 Water content (A) and germination percentage (B) of Epimedium sagittatum, Zea mays and Oryza sativa seeds
在吸胀的72 h内,未观察到箭叶淫羊藿种子萌发;相反,玉米和水稻种子随着吸水量的增加逐渐萌发。玉米和水稻种子达到10%萌发率的时间分别为22 h和33 h,达到50%萌发率的时间分别为34 h和42 h。当吸胀72 h时,玉米和水稻种子的萌发率分别达到97%和93% (图1:B)。
2.2 脱水对种子萌发的影响
为研究箭叶淫羊藿种子的耐脱水性,将含水量为9.7% (鲜重为基础) 的箭叶淫羊藿种子置于干燥硅胶表面在不同的时间进一步脱水。脱水初期,种子的含水量下降较快,之后缓慢下降。如脱水2 h时,种子的含水量下降了18.9%;从脱水8 h至脱水12 h,含水量仅下降了4.2% (图2:A)。

a. 含水量的变化; b. 萌发率的变化。种子在干燥硅胶表面被分别脱水不同的时间后, 测定种子的含水量和萌发率。脱水后的种子先在4 ℃下层积60 d,然后在10 ℃和黑暗条件下萌发72 d。胚根伸出种皮2 mm作为种子完成萌发的标准,所有的数据为50粒种子4次重复的平均值 ± 标准误。下同。条形图上相同的大写字母表示不同处理之间没有显著性差异 (S-N-K, P=0.05)。a. Change of water content; b. Change of germination percentage. Water content and germination percentage of seeds were determined after seeds were dehydrated at surface of dry silica gel for different time. Dehydrated seeds were first stratified at 4 ℃ for 60 d, and then germinated at 10 ℃ and darkness for 72 d. A radicle length of 2 mm was used as the criterion for completion of germination. All values are of four replicates of 50 seeds each. The same below. The same uppercase letters above bars represent no significant differences among different treatments (S-N-K, P=0.05).图 2 箭叶淫羊藿种子对脱水的反应Fig. 2 Response of Epimedium sagittatum seeds to dehydration
脱水4 h和8 h,当含水量从9.7%下降到7.3%和5.7%时,种子 (经4 ℃层积60 d后) 的萌发率增加;随着进一步脱水,种子的萌发率稍微下降,但仍高于对照,与对照没有显著性差异 (图2:B)。脱水后,种子的萌发速率加快,如未经脱水的种子在萌发处理后155 d时开始萌发,而经过脱水的种子在萌发处理后105 d时开始萌发 (未表明数据)。
2.3 温度对种子萌发的影响
为发现箭叶淫羊藿种子的适宜萌发温度,将采集的新鲜干燥种子分别在4、10、15、20、25 ℃和黑暗条件下吸胀30 d,其萌发率为零 (未表明数据)。但是,在4 ℃和10 ℃的条件下,种子分别在吸胀的115 d和145 d 开始萌发,随着吸胀时间的增加,种子的萌发率也增加;当吸胀240 d时,种子的萌发率分别为55%和44% (图3:A)。而在15、20、25 ℃的条件下,到吸胀的240 d,种子的萌发率分别为2%、0和0。种子在4 ℃和10 ℃的条件下能够部分萌发,在4 ℃下的萌发速率和萌发率明显高于10 ℃ (图3:A)。

A. 采集的新鲜干燥种子; B. 10 ℃层积60 d后转移到4℃层积30 d的种子。A. Freshly-collected dry seeds; B. Seeds stratified at 10 ℃ for 60 d and transfered to 4 ℃ for 30 d.图 3 温度对箭叶淫羊藿种子萌发的影响Fig. 3 Effects of temperature on germination of Epimedium sagittatum seeds
为进一步找到种子的适宜萌发温度,将在10 ℃层积60 d后转移到4 ℃中层积30 d的种子置于不同温度和黑暗条件下萌发。结果发现,在4、10、15、20、25 ℃下,种子的萌发率分别为88%、85%、3%、2%和0,适宜的萌发温度为4 ℃和10 ℃。在萌发初期,种子在10 ℃中的萌发速率稍微高于4 ℃,但随着萌发时间的增加,在4 ℃下的萌发速率和萌发率都高于10 ℃ (图3:B)。
2.4 层积处理对胚生长和种子萌发的影响
2.4.1 层积对胚生长的促进作用 箭叶淫羊藿种子从果荚脱落时,种子中胚的体积较小,呈球形胚,E/S比值仅为0.07 (图4:A,B)。 随着在4 ℃和10 ℃中层积时间的增加,胚逐渐从球形胚发育到心形胚、鱼雷形胚和成熟胚,胚的体积也不断增加 (图4)。

采集的新鲜干燥种子用不同的温度组合层积表明的时间后,用体式显微镜观察和测定胚的大小,拍照和计算E/S比值。A,B. 在4 ℃和10 ℃层积过程中胚的形态变化; C-E. 在不同温度组合层积过程中E/S比值的变化。图中箭头表示温度变化的时间点。所有的数据为10粒种子3次重复的平均值±标准误。The freshly-collected dry seeds were stratified in different temperature combinations for an indicated time, and the embryo size was then observed, measured, and photographed by stereomicroscope, and the E/S ratio was calculated. A,B. Morphological change of embryo during stratification at 4 ℃ and 10 ℃, respectively; C-E. Change of E/S ratio during stratification at different temperature combinations. The arrow in the figure indicates the time point of temperature change. All values are of three replicates of 10 seeds each.图 4 不同层积处理对箭叶淫羊藿胚生长 (胚/种子, E/S) 的影响Fig. 4 Effects of different stratification treatments on embryo growth (embryo/seed, E/S) of Epimedium sagittatum seeds
当种子在4 ℃中层积时,胚在层积0~60 d时生长缓慢 (E/S比值较小),到层积60 d时,E/S比值为0.09;在层积的61~90 d里胚逐渐生长,到层积90 d时E/S比值为0.14;在层积90 d后胚迅速生长,到层积165 d时E/S比值为0.88 (图4:C)。
当种子在10 ℃中层积时,胚的生长逐渐增大;到层积165 d时,其E/S比值为0.8 (图4:D)。我们观察到,在10 ℃中层积的种子,其E/S比值显著大于在4 ℃中层积的种子,如种子在10 ℃中层积60 d的E/S比值 (0.24) 比在4 ℃中层积相同时间的比值(0.09) 增加了167% (图4:D)。
当种子在4 ℃中层积30 d后转移到10 ℃中层积时,胚的生长速率比一直在4 ℃中层积生长更快。例如,从4 ℃中层积30 d后转到10 ℃中继续层积60 d的种子,其E/S比值 (0.25) 比在4 ℃中层积90 d的种子 (0.144) 增加了约74%;在4 ℃中层积60 d后转移到10 ℃中层积的种子其E/S比值在层积初期与一直在4 ℃中层积的种子类似,随着在10 ℃中层积时间的延长,其E/S比值有所增加 (图4:C)。种子在10 ℃中层积30 d或60 d后转移到4 ℃中层积时,其E/S比值均显著大于一直在10 ℃中层积种子的E/S比值。例如,种子一直在10 ℃中层积90 d时,其E/S比值为0.4;在10 ℃中层积30 d后转移到4 ℃中层积60 d,其E/S比值为0.83;在10 ℃中层积60 d后转移到4 ℃中层积30 d, 其E/S比值为0.67 (图4:D)。
种子在4 ℃层积30 d和10 ℃层积90 d后转移到4 ℃中层积,或在4 ℃层积60 d和10 ℃层积60 d后转移到4 ℃中层积,胚的生长 (E/S) 都表现出相同的趋势;而在层积75~135 d时,前者的生长比后者稍微快些,在136~165 d时,二者的生长没有明显差异;到层积165 d时,二者的E/S比值均大于0.8 (图4:E)。
2.4.2 层积对种子萌发的促进作用 种子在4 ℃中分别层积30 d或60 d后,置于10 ℃和黑暗中萌发,随着层积时间的延长,种子的萌发速率和萌发率显著增加 (图5:A)。种子在10 ℃中层积30 d或60 d后,置于4 ℃和黑暗中萌发,种子的萌发速率和萌发率也显著增加,在10 ℃中层积30 d和60 d的作用效果类似 (图5:B)。

A. 种子在4 ℃中分别层积30 d和60 d, 之后在10 ℃和黑暗条件下萌发; B. 种子在10 ℃中分别层积30 d和60 d,之后在4 ℃和黑暗条件下萌发; C. 种子在4 ℃中层积30 d后转移到10 ℃中继续层积90 d,或在4 ℃中层积60 d后转移到10 ℃中继续层积60 d,之后在4 ℃和黑暗条件下萌发。图中箭头表示种子开始萌发的时间点。胚根伸出种皮2 mm作为种子完成萌发的标准。所有的数据为50粒种子4次重复的平均值±标准误。下同。A. Seeds were stratified at 4 ℃ for 30 and 60 d, respectively, and then germinated at 10 ℃ and darkness; B. Seeds were stratified at 10 ℃ for 30 and 60 d, respectively, and then germinated at 4 ℃ and darkness; C. Seeds are stratified at 4 ℃ for 30 d and transfered 10 ℃ for 90 d or at 4 ℃ for 60 d and at 10 ℃ for 60 d, and then germinated at 4 ℃ and darkness. The arrow in the figure indicates the time point at which the seed begins to germinate. A radicle length of 2 mm was used as the criterion for completion of germination. All values are of four replicates of 50 seeds each. The same below.图 5 不同层积处理对箭叶淫羊藿种子萌发的影响Fig. 5 Effects of different stratification treatments on germination of Epimedium sagittatum seeds
种子在10 ℃中层积30 d或60 d后在4 ℃中的萌发速率和萌发率远比在4 ℃中层积30 d或60 d后在10 ℃ 中萌发的种子高得多 (图5:A,B)。例如,4 ℃中层积30 d和60 d的种子在10 ℃中的最初萌发时间分别为125 d和90 d,在萌发150 d时的萌发率分别为22.5%和67%;在10 ℃中层积30 d和60 d的种子在4 ℃中的最初萌发时间均为45 d,在萌发150 d时的萌发率分别为93%和96 %。在4 ℃中层积60 d的种子在10 ℃中萌发率达到50%的时间为135 d,而在10 ℃中层积30 d的种子在4 ℃中萌发率达到50%的时间仅为60 d(图5:A,B)。
两种变温层积 (4 ℃层积30 d + 10 ℃层积90 d或4 ℃层积60 d + 10 ℃层积60 d) 都能显著增加种子在4 ℃中的萌发速率和萌发率,萌发率达到50%的时间均为70 d (图5:C);与在10 ℃中层积30 d 或60 d后在4 ℃中萌发的种子比较,种子的萌发速率和萌发率稍微降低,但比在4 ℃中层积30 d或60 d后在10 ℃中萌发的种子高得多 (图5)。
2.5 赤霉素和氟啶酮对种子萌发的促进作用
为了解箭叶淫羊藿种子萌发对植物激素GA3和氟啶酮的响应,种子先在10 ℃和黑暗中层积120 d,使胚发育成熟,再在不同浓度的GA3、氟啶酮和GA3+ 氟啶酮中萌发。结果表明,1、10 μmol·L-1GA3对种子的萌发没有作用,100~10 000 μmol·L-1GA3明显地促进种子萌发,10 000 μmol·L-1GA3的促进作用明显低于1 000 μmol·L-1GA3,以及种子的萌发率仍然低于70% (图6:A)。5~500 μmol·L-1氟啶酮显著地促进种子萌发,其萌发率都高于90%,均比对照增加46%及以上 (图6:B)。在添加10、100 μmol·L-1GA3的同时,分别添加10 μmol·L-1或100 μmol·L-1的氟啶酮,种子的萌发速率 (未表明数据) 和萌发率都比单独添加相同浓度的GA3高得多,萌发率约为95% (图6:C)。
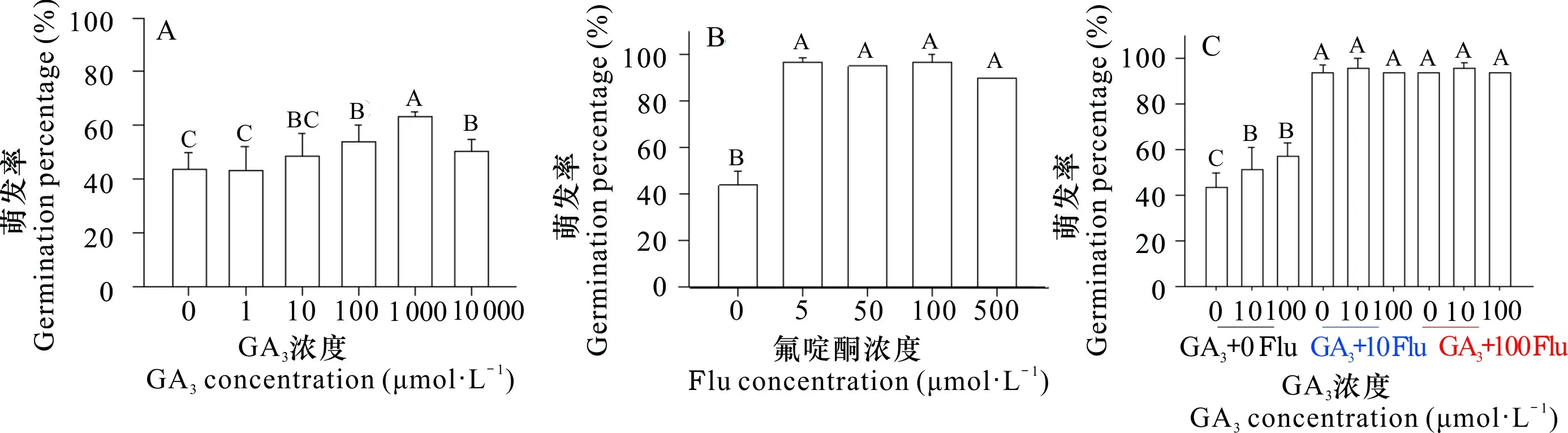
种子在10 ℃和黑暗中层积120 d,之后在不同浓度的GA3 (A)、Flu (B) 和GA3 + Flu 组合 (C) 以及4 ℃和黑暗中萌发60 d。Seeds were stratified at 10 ℃ and darkness for 120 d, and then germinated in different concentrations of GA3 (A), Flu (B) and GA3 + Flu combinations (C) as well as at 4 ℃ and darkness for 60 d.图 6 植物激素赤霉素 (GA3) 和氟啶酮 (Flu) 对淫羊藿种子萌发的影响Fig. 6 Effects of phytohormone gibberellin (GA3) and fluridone (Flu) on germination of Epimedium sagittatum seeds
3 讨论
3.1 箭叶淫羊藿种子对水分的吸收
种子萌发随着水分吸收开始,当胚根突破种皮萌发结束 (Bewley et al., 2013)。箭叶淫羊藿种子的水分吸收具有快速吸水 (阶段Ⅰ) 和缓慢吸水 (阶段Ⅱ) 两个阶段,缺乏吸水的第Ⅲ阶段。在阶段Ⅰ,种子的含水量迅速增加到52%,表明箭叶淫羊藿种子的种皮是透水的,不具有物理休眠。阶段Ⅰ的水分吸收是由衬质势 (matrix potential) 驱动的,因为干种子的水势非常低 (Obroucheva & Antipova, 1997)。阶段Ⅱ的缓慢吸水直到72 h (甚至更长,未表明数据) 是水分吸收的滞后期。以上结果与巫山淫羊藿种子的水分吸收类似 (樊家乙等, 2010)。阶段Ⅲ的水分吸收是由种子萌发引起的;在吸胀的72 h内,箭叶淫羊藿种子的萌发率为零,没能观察到水分吸收的第Ⅲ阶段。在25 ℃下,玉米和水稻种子的水分吸收具有阶段Ⅰ的快速吸水,阶段Ⅱ的短暂缓慢吸水以及阶段Ⅲ的迅速吸水。玉米和水稻种子的萌发率随着吸水逐渐增加,分别在吸胀后的34 h和42 h达到50%萌发率。
3.2 脱水对种子萌发的影响
成熟脱水是正常性种子 (orthodox seed) 发育的末端事件,在植物种子 (质) 资源保存中起关键作用 (宋松泉等, 2022)。箭叶淫羊藿种子的含水量从9.7%下降至4.6%,种子经4 ℃层积60 d后的萌发率仍然为56.3%,表明种子具有耐脱水性。值得注意的是,种子萌发率低于60%,不是由于种子被脱水伤害,而是在4℃层积60 d未能完全解除种子的休眠特性。本研究结果为淫羊藿属植物种子的干燥及其种质资源的长期保存提供了耐脱水的依据。
3.3 温度对种子萌发的影响
温度既是影响种子萌发最重要的环境因素之一,也决定种子萌发的能力和速率,并打破种子的初生和/或次生休眠 (Brándel 2004; Baskin & Baskin, 2014)。箭叶淫羊藿种子分别在4、10、15、20、25 ℃和黑暗条件下吸胀30 d,其萌发率为零,表明种子具有休眠特性。然而,在4 ℃和10 ℃的条件下,种子分别在吸胀115 d和145 d时开始萌发;到吸胀240 d时,种子的萌发率分别为55%和44%;在15、20、25 ℃的条件下,到吸胀240 d时,种子的萌发率分别为2%、0和0。这表明箭叶淫羊藿种子具有深休眠特性,温度显著影响种子的萌发,以及4~10 ℃可能是箭叶淫羊藿种子萌发的适宜温度。苏贺等 (2016) 的研究结果表明,巫山淫羊藿种子只能在低温 (4 ℃) 中萌发,不能在变温 (10 ℃/20 ℃) 中萌发。
在10 ℃层积60 d后转移到4 ℃中层积30 d的种子,在4、10、15、20、25 ℃下的萌发率分别为88%、85%、3%、2%和0,表明种子的适宜萌发温度为4~10 ℃。
3.4 层积处理对胚生长和种子萌发的影响
Graeber等 (2012) 提出,休眠是一种数量性状,其深度随着发育进程而变化。初生休眠是种子在成熟过程中形成的,在生理成熟时达到最高水平;在随后的干藏 (后熟) 或层积过程中,种子的休眠深度逐渐降低 (Finch-Savage & Leubner-Metzger, 2006; Holdsworth et al., 2008)。箭叶淫羊藿种子脱落时,种子中胚的体积较小 (E/S比值为0.07);随着种子在4、10 ℃中层积时间的增加,胚逐渐从球形胚发育到心形胚、鱼雷形胚和成熟胚 (E/S比值大于0.8);这表明箭叶淫羊藿种子具有形态休眠,在4、10 ℃层积过程中,胚能够逐渐生长发育。田向荣等 (2015) 观察到类似的结果,研究发现箭叶淫羊藿种子的胚未分化,停留在球形胚阶段,在后熟过程中,胚的结构逐步发育,形成子叶胚。朝鲜淫羊藿 (王晶等, 2013) 和巫山淫羊藿 (樊家乙等, 2010) 种子也具有类似的特性,需要经过一定时间的层积才能促进胚的生长。这些结果与朝鲜淫羊藿在田间条件下胚的生长、萌发和出苗的物候一致。朝鲜淫羊藿种子的胚未发育,在田间条件下在6—9月初,胚很少生长,但从9月下旬至11月胚迅速生长,到12月初胚已完成生长,胚的长度比初始长度增加了16倍;种子在次年3月萌发,萌发率达84.4% (Rhie & Lee, 2020)。
当种子在4 ℃中层积30 d后转移到10 ℃中层积时,胚的生长速率比一直在4 ℃中层积生长更快;在10 ℃中层积的种子,其E/S比值显著大于在4 ℃中层积的种子。这表明10 ℃层积有利于胚的生长发育。此外,当种子在10 ℃中层积30 d或60 d后转移到4 ℃中层积时,其E/S比值均显著大于一直在10 ℃中层积种子的E/S比值,表明胚和/或胚乳中可能存在某些生长抑制物质,在4 ℃中层积比在10℃中层积更有利于这些生长抑制物质的释放和胚的生长。本研究结果表明,变温层积有利于箭叶淫羊藿种胚的生长发育。Rhie和Lee (2020) 发现,当朝鲜淫羊藿种子在20 ℃或15 ℃的恒温下培养时,胚几乎不生长。
在10 ℃萌发条件下,箭叶淫羊藿种子的萌发速率和萌发率随着在4 ℃中层积时间的延长而显著增加。同样,在4 ℃萌发条件下,箭叶淫羊藿种子的萌发速率和萌发率也随着在10 ℃中层积时间的延长而显著增加;而在10 ℃中层积30 d或60 d后在4 ℃中的种子萌发速率和萌发率却远远高于在4 ℃中层积30 d或60 d后在10 ℃ 中萌发的种子 。这些萌发结果与层积温度对E/S比值的影响类似。付长珍等 (2012) 报道,拟巫山淫羊藿种子在15 ℃中处理1~7个月后转移到4 ℃中,胚的生长和种子萌发率显著增加,但随着15 ℃处理时间的延长,种子的萌发率增加,而种子的萌发速率下降。
4 ℃层积30 d + 10 ℃层积90 d或4 ℃层积60 d + 10 ℃层积60 d的变温层积都能显著增加种子在4 ℃中的萌发速率和萌发率,这与变温层积对E/S比值的影响类似。
3.5 赤霉素和氟啶酮对种子萌发的促进作用
ABA是休眠诱导与维持的正调控因子,是种子萌发的负调控因子;而GA3能释放休眠,促进萌发和拮抗ABA的效应,它们的水平和信号转导在种子休眠释放和萌发中起重要作用 (Hauvermale et al., 2012; 宋松泉等, 2020a, b; Iwasaki et al., 2022)。对于在10 ℃和黑暗中层积120 d 的 (胚成熟) 箭叶淫羊藿种子,1、10 μmol·L-1GA3对种子的萌发没有作用,100~10 000 μmol L-1GA3明显地促进种子萌发,但种子的萌发率仍然低于70%。这表明GA3能部分促进种子的萌发。5~500 μmol·L-1氟啶酮显著促进种子的萌发,其萌发率均大于90%;对于种子的萌发,10 μmol·L-1或100 μmol·L-1的氟啶酮对GA3有加成作用。苏贺等 (2016) 利用变温 (10 ℃/20 ℃) 层积90 d 的巫山淫羊藿种子作为材料,研究了氟啶酮和赤霉素对巫山淫羊藿种子休眠解除的影响,发现氟啶酮的作用较强,赤霉素的作用较弱,并且氟啶酮可以促进在变温中不能萌发的种子萌发。Rhie和Lee (2020) 观察到,GA3处理能增加朝鲜淫羊藿种子胚的生长,但对种子萌发的促进作用较小,萌发率低于10%。这些结果都表明,ABA在抑制箭叶淫羊藿种子萌发中可能具有重要作用。
4 结论
箭叶淫羊藿种子的种皮透水,不具有物理休眠;种子具有耐脱水性,能在低含水量和低温的条件下贮藏;在4~25 ℃和黑暗条件下,种子的萌发率为零;E/S比值非常小,4、10 ℃和变温层积能显著促进胚的发育以及增加种子的萌发速率和萌发率;GA3和氟啶酮能显著增加种子的萌发速率和萌发率。根据Baskin和Baskin (2004, 2014)、Finch-Savage和Leubner-Metzger (2006) 的观点,本研究认为,箭叶淫羊藿种子的休眠类型为MPD,释放休眠和促进萌发的最适方法是将种子先在10 ℃中层积30 d,再置于4 ℃的环境中让其萌发。
猜你喜欢
安徽农业科学(2022年18期)2022-10-13
江西中医药(2022年8期)2022-08-22
空间科学学报(2021年2期)2021-07-21
环球中医药(2021年3期)2021-04-07
花卉(2020年24期)2021-01-04
东北农业大学学报(2020年11期)2020-12-17
湖南林业科技(2020年5期)2020-12-07
特产研究(2020年4期)2020-08-13
空间科学学报(2020年2期)2020-04-01
西南农业学报(2019年1期)2019-03-01
