
水稻OsZAT12基因响应非生物胁迫和植物激素的研究
2022-12-30 06:58:48陈言博陈宗新夏快飞张明永王亚琴
广西植物 2022年11期
陈言博, 陈宗新, 夏快飞, 张明永, 王亚琴*
(1. 华南师范大学 生命科学学院, 广州 510631; 2. 中国科学院华南植物园, 广州 510650 )
锌指蛋白是真核生物体内一类被广泛研究的转录因子家族,根据半胱氨酸(C)和组氨酸(H)残基的数目和位置可将其分为C2H2、C2HC、C2C2、C2C2C2C2等类型(Laity et al., 2001)。C2H2型锌指蛋白是锌指蛋白中最常见的一类,在许多代谢途径以及植物的生长发育和非生物胁迫反应中起着至关重要的作用(Ballerini et al., 2020; Yin et al., 2020; Rodas et al., 2021; Zhang et al., 2021)。目前,在水稻和拟南芥中已经发现分别有189个和176个C2H2型锌指蛋白,该类蛋白的锌指结构域具有CX2-4CX3FX5LX2HX3-5H(C为半胱氨酸,F为苯丙氨酸,H为组氨酸,L为亮氨酸,X为任意氨基酸)特征序列(Englbrecht et al., 2004; Agarwal et al., 2007)。根据锌指结构域的数目、序列和排列方式,拟南芥176个C2H2型锌指蛋白可分为Set A、Set B和Set C,其中包含多个且离散的锌指结构域的一类归为Set C(Pabo et al., 2001; Englbrecht et al., 2004; Ciftci-Yilmaz & Mittler, 2008)。根据锌指特征序列中2个组氨酸之间的氨基酸数目,Set C可进一步分为C1、C2和C3,C1家族可进一步细分为C1-1i、C1-2i、C1-3i、C1-4i和C1-5i(Englbrecht et al., 2004)。有关C1家族的研究主要集中在C1-1i和C1-2i亚家族(Englbrecht et al., 2004; Ciftci-Yilmaz & Mittler, 2008)。
双子叶植物拟南芥C2H2型锌指蛋白的C1-2i亚家族包括ZAT5~7、ZAT10~12、ZAT18和AZF1~3等,该亚家族含有2个锌指结构,锌指特征序列中2个组氨酸之间的氨基酸数目为3个,大部分成员具有核定位信号及EAR motif(ethylene-responsive element binding factor-associated amphiphilic repression motif),主要参与各种生物和非生物胁迫响应(Lippuner et al., 1996; Meissner et al., 1997; Englbrecht et al., 2004; Sakamoto et al., 2004; Mittler et al., 2006; Liu et al., 2013; Shi et al., 2014; Yin et al., 2017)。在拟南芥中,AtAZF2对盐和干旱胁迫处理均响应强烈,而AtAZF1和AtAZF3的表达对非生物胁迫(盐、干旱和冷)的响应较弱,只有AtAZF2能够被ABA(abscisic acid)诱导,这可能是由于AtAZF2启动子中含有ABA响应元件(Sakamoto et al., 2004)。过表达AtZAT18可以提高拟南芥的耐旱性,而突变AtZAT18则导致植物对干旱胁迫的耐受性降低(Yin et al., 2017)。组成型过表达AtZAT10的转基因拟南芥生长受到抑制,并对干旱、盐、高温和渗透胁迫的耐受性增强,同时提高了ROS稳态相关基因,如AtAPX1和AtAPX2的转录(Sakamoto et al., 2004; Mittler et al., 2006)。有趣的是,AtZAT10基因敲除植株和RNAi干涉植株也表现出对盐和渗透胁迫的耐受性增加,但调控机制目前尚不清楚(Mittler et al., 2006)。除拟南芥外,其他双子叶植物中也有关于C2H2转录因子的报道。豌豆St(Stipulesreduced)基因通过影响细胞分裂和细胞扩展调控豌豆托叶的大小(Moreau et al., 2018)。番茄基因H(hair)编码C2H2型锌指蛋白,过表达该基因后叶片毛状体数量显著增加(Chang et al., 2018)。苜蓿MtSUP(SUPERMAN)是C2H2锌指蛋白,主要在分生组织、雄蕊、心皮边缘等部位表达,在苜蓿中将该基因突变后影响花器官的数量和形态及果实发育(Rodas et al., 2021)。
禾本科植物结缕草中的ZjZFN1基因,编码C2H2型锌指蛋白,其表达受盐胁迫、冷和ABA诱导,在拟南芥中异源过表达ZjZFN1发现,该基因通过影响活性氧的积累及盐胁迫响应基因的转录,提高拟南芥对盐胁迫的抗性(Teng et al., 2018)。干旱胁迫诱导小麦TaZFP1B的表达,而过表达TaZFP1B的小麦对干旱胁迫的抗性显著增加(Cheuk et al., 2020)。水稻中关于C2H2锌指蛋白有一些文献报道,如ZFP182、ZFP36、ZFP179、ZFP245和ZFP252等。过表达ZFP182能提高植物对盐胁迫的耐受性(Zhang et al., 2012)。过表达ZFP36能够提高抗氧化酶的活性,并增强水稻对干旱胁迫和氧化胁迫的耐受性;相反,ZFP36的RNAi干涉植株中抗氧化酶活性较低,对干旱胁迫和氧化胁迫更敏感(Zhang et al., 2014)。过表达ZFP179能够提高水稻的耐盐性,并且转基因幼苗对外源ABA更加敏感(Sun et al., 2010)。过表达ZFP252的植株对盐和干旱胁迫的耐受性增加,在盐和干旱胁迫处理下,过表达ZFP252植株中OsDREB1A、OsP5CS和OsProT等非生物胁迫相关基因的表达量高于野生型和ZFP252沉默株系,说明OsDREB1A、OsP5CS和OsProT可能是ZFP252的下游基因(Xu et al., 2008)。
OsZAT12属于C2H2锌指蛋白的C1-2i亚家族,该类基因在许多代谢途径以及植物的生长发育和非生物胁迫反应中起着至关重要的作用(Ballerini et al., 2020; Yin et al., 2020; Zhang et al., 2021; Rodas et al., 2021),本实验室前期(陈宗新等,2019)克隆了OsZAT12基因,该基因在水稻根中特异表达,定位于细胞核,异源过表达OsZAT12拟南芥植株矮小,根生长受到抑制。水稻为重要的粮食作物,生物/非生物胁迫、植物激素等均影响植株发育和形态建成,进而影响产量。然而,作为在植物生长发育和非生物胁迫中可能起着重要作用的OsZAT12,在水稻生长发育及非生物胁迫中的作用尚不清楚。因此,本文分析了OsZAT12的启动子元件和转录活性,并采用qRT-PCR技术分析OsZAT12在非生物胁迫和植物激素处理下的响应模式,以期为进一步研究OsZAT12参与不同非生物胁迫应答的分子机制和参与ABA信号转导途径的调控奠定基础。
1 材料与方法
1.1 材料
粳稻“中花11”(WT),实验室自存。双元载体pCAMBIA1301来自本实验室,CRISPR编辑相关载体pYLCRISPR/Cas9Pubi-H、pYLgRNA-OsU6a/LacZ、pYLgRNA-OsU6a、pYLgRNA-OsU3/LacZ、pYLsgRNA-OsU3由华南农业大学刘耀光实验室惠赠,大肠杆菌DH5a感受态菌种和根癌农杆菌EHA105感受态菌种由本实验室保存。
1.2 OsZAT12及其同源基因的保守结构域分析
在NCBI和TAIR数据库中查找拟南芥和水稻中的C1-2i亚家族成员,并导出这些基因的氨基酸序列。利用软件ClusatlX 1.83进行多重序列比对,使用软件DNAMAN 6.0输出图片。本研究中,进行多重序列比对的基因为AtAZF1(At5g67450)、AtAZF2(At3g19580)、AtAZF3(At5g43170)、AtZAT5(At2g28200)、AtZAT6(At5g04340)、AtZAT7(At3g46090)、AtZAT10(At1g27730)、AtZAT11(At2g37430)、AtZAT12(At5g59820)、AtZAT18(At3g53600)、OsZFP252(AAO46041.1)、OsZFP245(AAQ95583)、OsZFP182(NP001051718.1)、OsZFP179(AAL76091.1)、OsZFP36(AAP51130.1)。
1.3 OsZAT12转录活性分析
双荧光素酶报告系统是萤火虫荧光素酶(firefly luciferase,LUC)检测系统和海肾荧光素酶(renilla luciferase,REN)检测系统的组合,常用来分析转录因子的转录活性。本研究使用拟南芥原生质体瞬时表达系统进行双荧光素酶实验,其中拟南芥原生质体的提取参考Wu等(2009)的方法,双荧光素酶活性的检测根据Promega公司Dual-Luciferase Reporter Assay System试剂盒(Cat. No. E1910)中的说明书操作,并通过计算LUC/REN的比值分析OsZAT12的转录激活/抑制活性。
1.4 OsZAT12启动子分析
以水稻OsZAT12基因的起始密码子(ATG)上游2 000 bp 作为研究对象,利用在线启动子分析工具PLACE(http://www.dna.affrc.go.jp/PLACE/)和PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/),对OsZAT12的启动子序列进行调控元件的预测和分析。
1.5 OsZAT12基因在非生物胁迫和植物激素处理下的表达分析
1.5.1 水稻种子消毒及幼苗水培方法 水稻种子去壳后放入50 mL 离心管中,加75%乙醇表面消毒1 min,用2.5%次氯酸钠消毒50 min,无菌水漂洗5次,将种子置于1/2 MS固体培养基中培养。将以上培养5 d 的无菌苗移至96孔PCR塑料板(剪去管底)进行水培(16 h 光照/8 h 黑暗,白天28 ℃/夜晚24 ℃),水稻营养液配方参照“国际水稻研究所水稻营养液”配制,5~6 d 换一次水培液。
1.5.2 非生物胁迫处理方法 取苗龄14 d 的水稻幼苗为材料进行非生物胁迫处理。具体如下:低温胁迫,将水稻幼苗放置于4 ℃光照培养箱(16 h 光照/8 h 黑暗)中;渗透胁迫,将水稻幼苗置于含有20% PEG(polyethylene glycol)6 000的水稻营养液中;氧化胁迫,将水稻幼苗置于含有20 μmol·L-1甲基紫精(methyl viologen,MV)的水稻营养液中;盐胁迫处理,将水稻幼苗置于含有100 mmol ·L-1NaCl的水稻营养液中。分别收取处理0、0.5、1、3、6、12、24、48 h 的整株幼苗,未处理材料同时取样作为对照组,所有样品于液氮速冻后-80 ℃保存备用。除低温处理外,其他胁迫处理均在植物培养室(16 h 光照/8 h 黑暗,白天28 ℃/夜晚24 ℃)进行。
1.5.3 植物激素处理方法 取苗龄14 d 的水稻幼苗作为材料,进行植物激素处理。在培养液中分别添加100 μmol ·L-1脱落酸(abscisic acid,ABA),1 μmol ·L-12, 4-表芸苔素内酯(2,4-epibrassinolide, 2,4-eBL),1 μmol ·L-1吲哚-3-乙酸(indole-3-acetic acid,IAA),处理0、1、24、48 h 后取材(取整株幼苗),未处理材料同时取样作为对照组,所有样品于液氮速冻后-80 ℃保存。激素处理的材料均放在植物培养室(16 h 光照/8 h 黑暗,白天28 ℃/夜晚24 ℃)。
1.5.4 水稻RNA的提取及cDNA第一条链的合成 RNA提取根据华越洋超快型植物RNA提取试剂盒(Cat. No. 0416-50)的说明书进行。参照Toyobo反转录试剂盒(Cat. No. FSQ-301)说明书进行cDNA第一条链的合成。
1.5.5OsZAT12基因的qRT-PCR(quantitative real-time PCR)检测 本研究中的qRT-PCR采用SYBR染料法(GenStar 2 × RealStar Green Fast Mixture, Cat. No. A301-10)。以水稻eEF-1a基因为内参(qPCR-eEF-1a-F: 5′-GCACGCTCTTCTTGCTTTC-3′;qPCR-eEF-1a-R: 5′-AGGGAATCTTGTCAGGGTTG-3′),采用2-△△CT法计算(Livak & Schmittgen, 2001)OsZAT12基因的相对表达量(qPCR-OsZAT12-F: 5′-GACCTGAACCATCCACCCTG-3′; qPCR-OsZAT12-R: 5′-CGGTATCCAAGAACTGGTGGAA-3′)。
1.6 OsZAT12基因过表达载体和CRISPR/Cas9载体构建
由于OsZAT12在水稻根中特异表达(陈宗新等,2019),因此以野生型水稻根的cDNA为模板,利用引物OsZAT12-F: 5′-CGGGATCCATGAAGAGGTTTGCA-3′(酶切位点BamHI),OsZAT12-R: 5′-AACTGCAGCTAGTAGCCGACGCA-3′(酶切位点PstI)扩增OsZAT12基因,利用双酶切法构建到pCAMBIA1301载体上,得到过表达载体pCAMBIA1301::35S::OsZAT12。
利用华南农业大学刘耀光实验室设计的网站CRISPR-GE(http://skl.scau.edu.cn/)设计OsZAT12敲除靶点。为提高敲除效率,采用双靶点载体的策略,以OsZAT12基因设计2个靶点,其中靶点5′-CATGAGGCGCCACCGCGCCA-3′以U6为启动子,靶点5′-TGCGACGACATGAGCATCAG-3′以U3为启动子,具体载体构建参考Ma等(2015)的方法。
1.7 OsZAT12基因遗传转化水稻及转基因植株鉴定
将1.6中构建好的重组载体(过表达载体和CRISPR/Cas9载体)转化至根癌农杆菌EHA105感受态细胞中。采用农杆菌介导的遗传转化法将重组载体转进水稻愈伤组织中,具体参考李美茹和李洪清(2003)、王亚琴等(2011)的方法。
转基因植株的鉴定采用PCR方法。剪取T1代幼苗约2 cm 长的叶片,利用TPS溶液(100 mmol · L-1Tris-HCl, pH 8.0; 10 mmol · L-1EDTA-Na2, pH 8.0; 1 mol · L-1KCl)提取DNA。过表达OsZAT12的转基因植株利用抗性筛选标记基因HptII(hygromycin phosphotransferase II) 设计引物Hyg-F: 5′-GATGTTGGCGACCTCGTATT-3′, Hyg-R: 5′-TCGTTATGTTTATCGGCACTTT-3′,以DNA为模板进行PCR扩增检测。CRISPR植株采用引物CRISPR-F: 5′-TCAGACAACAGAGAGGTTGGT-3′和CRISPR-R: 5′-TAGTAGCCGACGCAGTCAAC-3′扩增OsZAT12包括靶位点在内的片段,经测序后检测突变位点。
1.8 转基因水稻幼苗对外源ABA的敏感性分析
取野生型、过表达OsZAT12和敲除OsZAT12植株的种子,经表面消毒后播种于含不同浓度ABA(0、1、5、10 μmol ·L-1)的1/2 MS培养基上,每个株系在不同ABA浓度培养基上播种30粒种子,在植物培养室(16 h 光照/8 h 黑暗,白天28 ℃/夜晚24 ℃)培养10 d 后拍照并统计株高和根长。
1.9 数据分析
所有实验均进行3次生物学重复,每次重复中每个样品设置3个技术重复。数据统计为平均值±标准偏差。利用软件SPSS Statistics中的单因素方差分析进行数据的显著性分析(*P<0.05;**P<0.01)。
2 结果与分析
2.1 转录因子OsZAT12保守结构域的分析
OsZAT12(Os05g0114400)基因编码区全长597 bp,该基因不含内含子,编码198个氨基酸(陈宗新等,2019)。为进一步研究OsZAT12蛋白结构域的保守性及其序列特点,对拟南芥和水稻C1-2i亚家族中的部分成员进行多重序列比对,结果如图1所示,OsZAT12的结构域与拟南芥和水稻中的同源蛋白一致,均含有2个包含QALGGH保守序列的锌指结构域以及1个EAR motif(ethylene-responsive element binding factor-associated amphiphilic repression motif)。蛋白C末端的EAR motif通常被认为具有抑制活性(Meissner & Michael, 1997; Englbrecht et al., 2004; Ciftci-Yilmaz & Mittler, 2008)。以上结果说明,OsZAT12是一个典型的C2H2型锌指蛋白,属于C1-2i亚家族,可能具有转录抑制活性。

黑色部分表示相似性=100%,粉色部分表示相似性在75%~100%之间(不包括100%),蓝色部分表示相似性在50%~75%之间(不包括75%),黄色部分表示相似性在33%~50%之间(不包括50%),白色部分表示相似性低于33%。Black parts indicate similarities = 100%, pink parts indicate similarities between 75% and 100% (excluding 100%), blue parts indicate similarities between 50% and 75% (excluding 75%), yellow parts indicate similarities between 33% and 50% (excluding 50%), the white parts indicate similarities <33%.图 1 水稻和拟南芥部分C2H2型锌指蛋白氨基酸序列的多重比对Fig. 1 Multiple alignment of amino acid sequences of partial C2H2 zinc finger proteins in Oryza sativa and Arabidopsis thaliana
2.2 转录因子OsZAT12的转录活性分析
采用双荧光素酶报告系统检测OsZAT12的转录活性,将OsZAT12与包含GAL4 DNA结合结构域(GAL4 DNA binding domain,GALBD)的效应载体融合(图2:A),同时与携带荧光素酶基因的报告载体共同转化拟南芥原生质体,检测对照组和实验组的萤火虫荧光素酶活性和海肾荧光素酶活性并计算相对LUC/REN比值。实验组的相对LUC/REN比值显著低于对照组(图2:B),说明转录因子OsZAT12具有转录抑制活性。
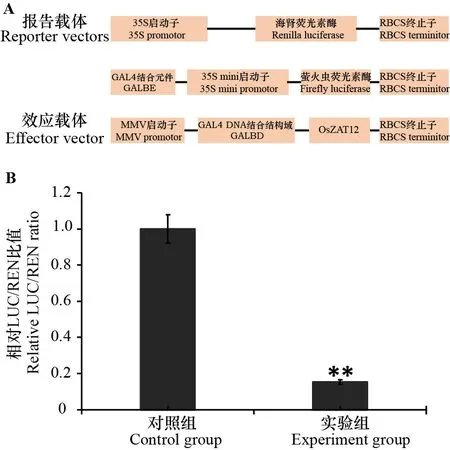
A. 双荧光素酶实验载体构建示意图,其中GALBE是GAL4结合元件,GALBD是GAL4 DNA结合结构域,35S mini启动子是只含46 bp 的35S启动子; B. 双荧光素酶报告系统检测OsZAT12的转录活性,报告载体和效应空载体作为对照组,报告载体和连有OsZAT12的效应载体作为实验组,实验结果为3次生物学重复的平均值±标准偏差。对照组的LUC/REN比值设为1,**表示与对照组相比达到显著水平(P<0.01)。A. Schematic structures of plasmids used in dual-luciferase assay, GALBE is GAL4 binding element, GALBD is GAL4 DNA binding domain, and 35S mini promoter indicates that 35S promoter only contains 46 bp; B. Dual luciferase reporter system detects the transcriptional activity of OsZAT12, the reporter and effector vectors serve as control group, the reporter vectors and the effector vector with OsZAT12 are used as the experimental group, values are ± s from three biological replicates. The LUC/REN ratio of control group is defined as 1, ** indicates significant difference compared with control group (P<0.01). 图 2 OsZAT12的转录活性分析Fig. 2 Transcriptional activity analysis of OsZAT12
2.3 OsZAT12启动子的序列分析
以水稻OsZAT12基因的起始密码子(ATG)上游2 000 bp 为研究对象,使用启动子在线分析网站对OsZAT12启动子进行调控元件预测,结果显示,OsZAT12启动子上调控元件非常丰富,除了基本的核心启动子元件(TATA box和CAAT box元件)以外,该段序列中还具有非生物胁迫和激素相关的顺式作用元件。其中,与非生物胁迫相关的有2个MYB转录因子结合元件;与激素相关的元件包括3个GA响应元件、3个ABA响应元件、2个Auxin响应元件和1个JA响应元件等(图3)。由此推测,OsZAT12基因的表达可能受非生物胁迫因子和多种激素的调节。
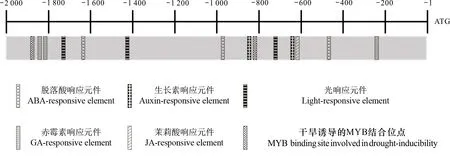
图 3 OsZAT12启动子的生物信息学分析Fig. 3 Bioinformatics analysis of OsZAT12 promoter
2.4 非生物胁迫处理下OsZAT12的表达分析
拟南芥ZAT12在活性氧以及非生物胁迫信号传导中起着重要作用(Davletova et al., 2005)。水稻OsZAT12启动子中含有非生物胁迫相关的元件(图3),推测其可能响应非生物胁迫。因此,进一步检测了水稻幼苗在低温(4 ℃)、渗透胁迫(20% PEG 6 000)、氧化胁迫(20 μmol·L-1MV)和盐胁迫(100 mmol·L-1NaCl)处理下OsZAT12的表达。如图4:A所示,低温处理48 h 内,OsZAT12的表达均呈下调趋势,其中在处理12 h时,OsZAT12的表达量降到最低,之后略微上调。在渗透胁迫处理下,OsZAT12表达量呈先下降后上升的趋势,从3 h 上升一直到48 h,OsZAT12的表达量达到未处理时的2倍左右(图4:B)。而在20 μmol·L-1MV和100 mmol·L-1NaCl的处理下,OsZAT12的表达量随时间变化未出现明显的上调或下调趋势(图4:C,D)。以上结果表明,OsZAT12基因的表达响应多种非生物胁迫,并随胁迫时间的延长其表达量出现不同的变化趋势。
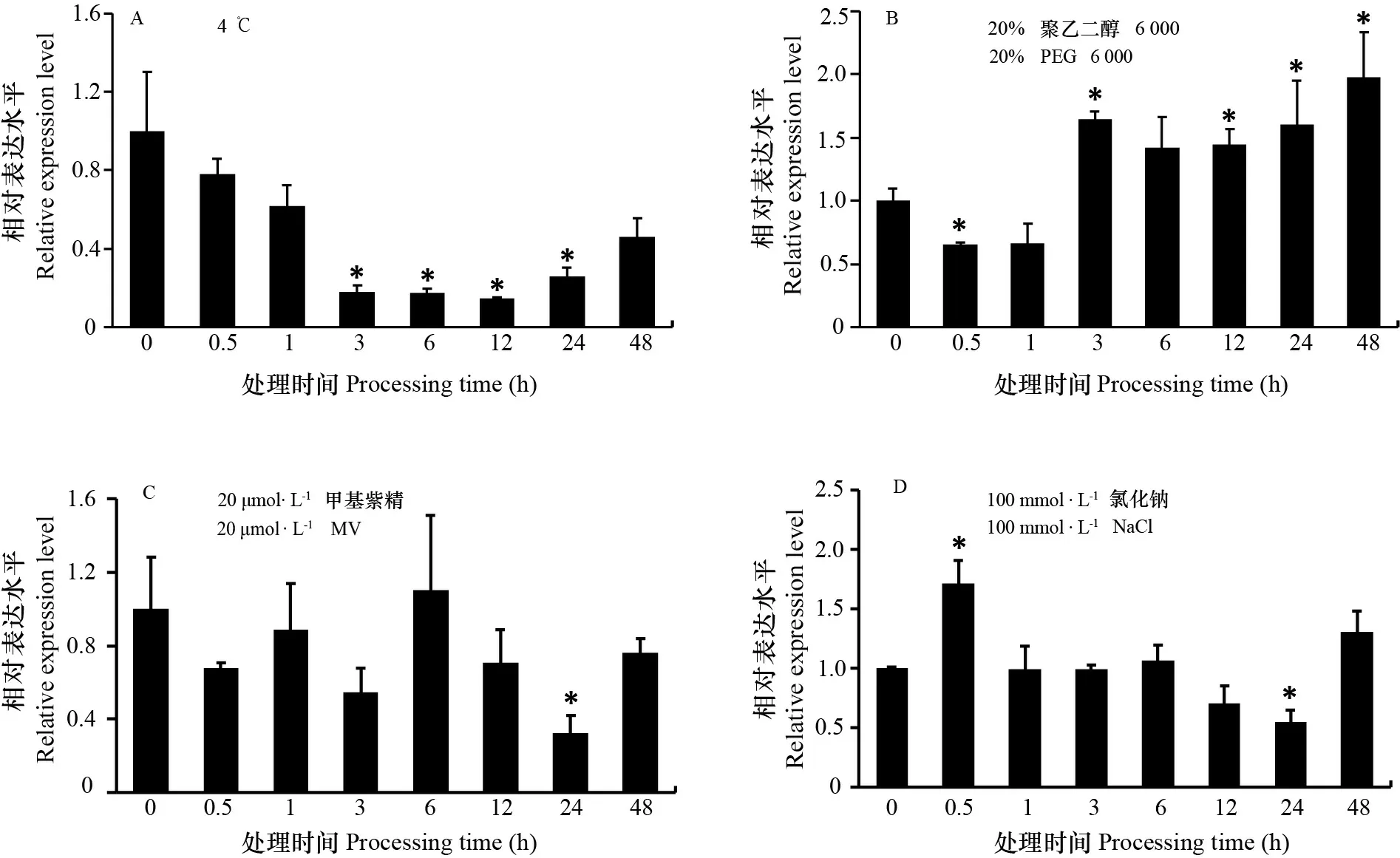
A. OsZAT12在4 ℃胁迫处理下的表达量分析; B. OsZAT12在20% PEG胁迫处理下的表达量分析; C. OsZAT12在20 μmol ·L-1 MV胁迫处理下的表达量分析; D. OsZAT12在100 mmol ·L-1 NaCl胁迫处理下的表达量分析。每组处理0 h 的相对表达量分别设为1,下同。*表示与对0 h比达到显著水平(P<0.05)。A. Expression level of OsZAT12 under 4 ℃ treatment; B. Expression level of OsZAT12 under 20% PEG treatment; C. Expression level of OsZAT12 under 20 μmol ·L-1 MV treatment; D. Expression level of OsZAT12 under 100 mmol·L-1 NaCl treatment. The relative expression level of 0 h after each treatment is defined as 1, the same below. * indicates significant differences compared with 0 h (P<0.05). 图 4 不同非生物胁迫下OsZAT12的表达水平Fig. 4 Expression levels of OsZAT12 under different abiotic stresses
2.5 不同植物激素处理下OsZAT12的表达分析
植物激素作为植物内源信号分子,在植物生长发育过程中具有重要作用。OsZAT12的启动子中含有多个激素相关元件(图3),推测其可能响应激素水平的变化。植物激素处理实验结果表明,外施ABA显著下调OsZAT12的表达。处理1 h 时OsZAT12的表达量迅速下降,在24 h 降到最低,约为对照的1/10,48 h 时略有升高(图5:A)。相反,外施1 μmol·L-1的2,4-eBL,在1 h 时OsZAT12 表达开始上调,一直持续到48 h 且达到最高,为对照的12倍(图5:B)。此外,IAA处理也能上调OsZAT12的表达,其表达量在24 h 达到最高,一直持续到48 h(图5:C)。以上结果表明,OsZAT12对不同植物激素的响应各异,可能参与不同植物激素信号对水稻生长发育的调控。
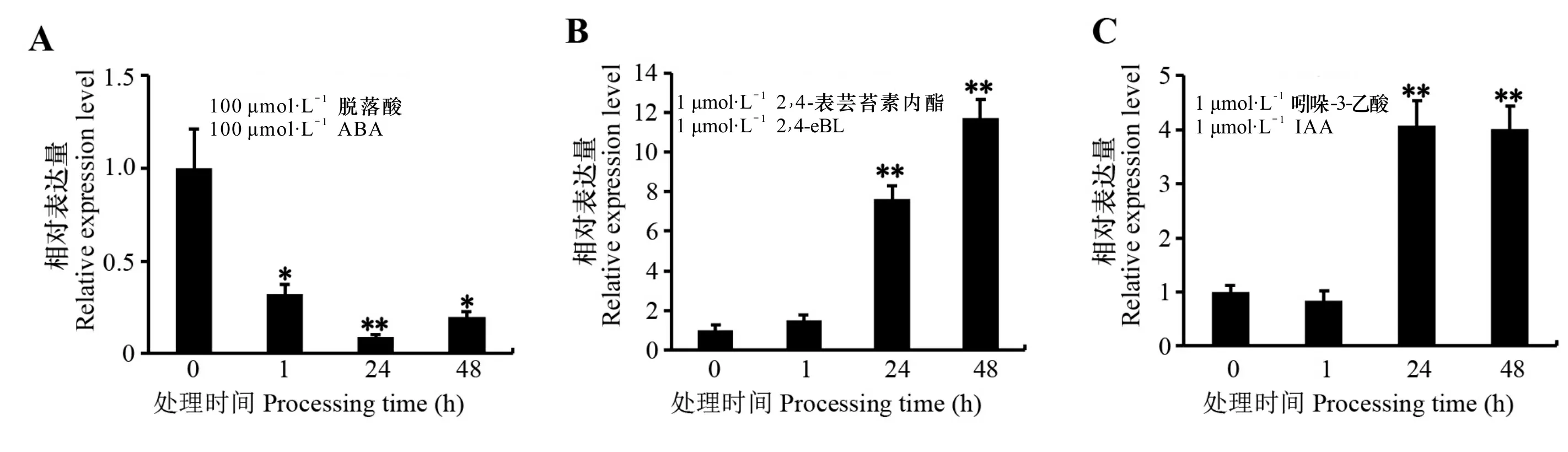
A. OsZAT12在100 μmol ·L-1 ABA处理下的表达量分析; B. OsZAT12在1 μmol ·L-1 24-eBL处理下的表达量分析; C. OsZAT12在1 μmol ·L-1 IAA处理下的表达量分析。*和**表示与0 h相比达到显著水平(*P<0.05,**P<0.01)。A. Expression level of OsZAT12 under 100 μmol ·L-1 ABA treatment; B. Expression level of OsZAT12 under 1 μmol ·L-1 24-eBL treatment; C. Expression level of OsZAT12 under 1 μmol ·L-1 IAA treatment. * and ** indicate significant differences compared with the 0 h (*P<0.05,**P<0.01).图 5 不同植物激素处理下OsZAT12的表达水平Fig. 5 Expression levels of OsZAT12 under different phytohormones treatments
2.6 OsZAT12转基因植株的获得
2.6.1OsZAT12敲除植株的筛选 使用特异检测引物(CRISPR-F和CRISPR-R)先对OsZAT12敲除植株的基因组DNA进行PCR扩增,再对得到的单一PCR片段产物进行测序,结果得到了3个敲除OsZAT12的纯合株系,其中oszat12-12-3和oszat12-8-15株系均为单碱基插入,而oszat12-10-10株系则是缺失一段序列(图6)。

图 6 不同OsZAT12敲除株系的测序结果Fig. 6 Sequencing results of different OsZAT12 knockout lines
2.6.2OsZAT12过表达植株的表达量检测 获得4个过表达OsZAT12纯合株系,并分别命名为OE8、OE3、OE9和OE5。qRT-PCR结果(图7)显示,与野生型相比,过表达株系OE8、OE3和OE5的表达量显著提高,OE9的表达量虽有所提高但与野生型无显著差异。因此,本研究选取OE8和OE3用于后续研究。
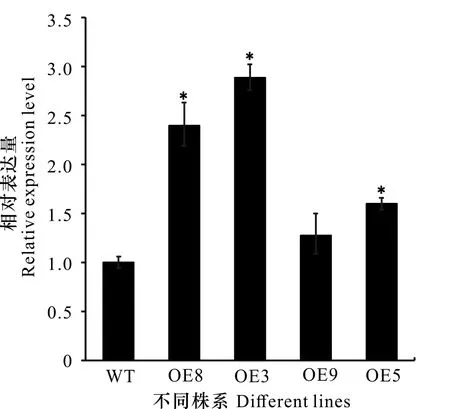
WT中OsZAT12基因的相对表达量设为1。*表示与WT相比达到显著水平(P<0.05)。Relative expression level of OsZAT12 in WT is defined as 1. * indicates significant differences compared with WT (P<0.05).图 7 qRT-PCR分析转基因植株中OsZAT12的表达量Fig. 7 qRT-PCR analysis of OsZAT12 expression in transgenic plants
2.7 OsZAT12转基因水稻对ABA的敏感性分析
ABA作为胁迫激素,是植物响应生物/非生物胁迫的重要调控因子(Chen et al., 2020; Bharath et al., 2021)。OsZAT12启动子序列中包含3个ABA响应元件(图3),并且ABA有抑制OsZAT12表达的作用(图5:A)。在获得过表达OsZAT12和敲除OsZAT12的植株后,进一步检测其对ABA的敏感性,结果表明,ABA抑制了野生型和OsZAT12超表达幼苗的生长,并随着ABA浓度的升高抑制程度越大,而OsZAT12过表达株系的株高、根长都显著高于野生型(图8)。OsZAT12敲除植株的株高在5 μmol ·L-1ABA处理下显著低于野生型;在低浓度ABA(1 μmol ·L-1)处理下,其根长显著高于野生型;但在较高浓度ABA(10 μmol ·L-1)处理下,根长与野生型无显著差异(图8)。以上结果表明,过表达OsZAT12会降低水稻对ABA的敏感性,而敲除OsZAT12则在合适的ABA浓度下才会增强水稻对ABA的敏感性。
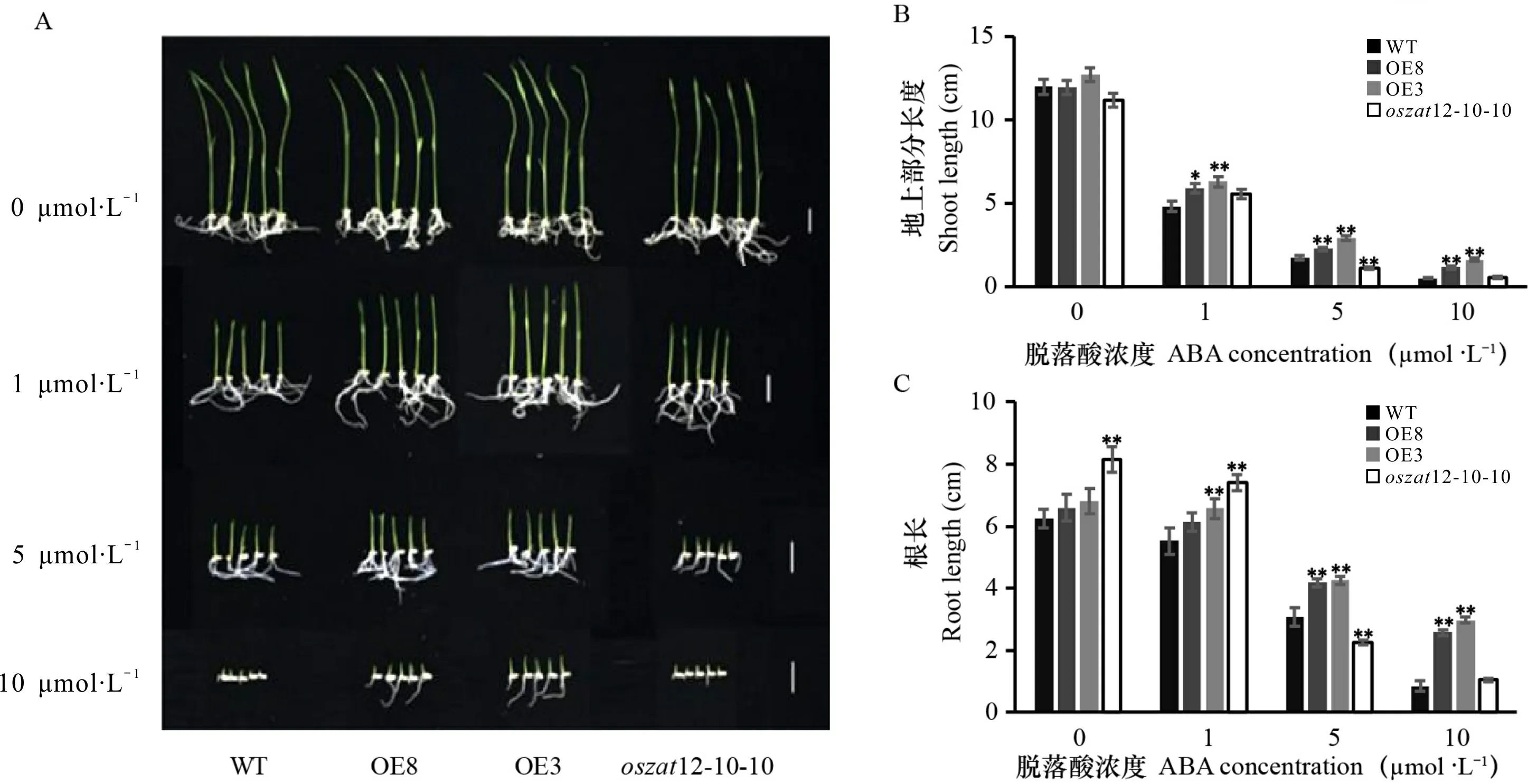
OsZAT12转基因水稻在不同浓度外源ABA处理下的表型 (A)、株高 (B)和根长 (C),标尺=2 cm。OE8、OE3为OsZAT12过表达植株;oszat12-10-10为OsZAT12敲除植株。*和**表示与WT相比达到显著水平(*P <0.05,**P<0.01), n=30。下同。Phenotype (A), shoot length (B) and root length (C) of OsZAT12 transgenic rice under different concentrations of exogenous ABA treatment, scale bar=2 cm. OE8 and OE3 are OsZAT12 overexpression plants; oszat12-12-3, oszat12-10-10 and oszat12-8-15 are OsZAT12 knockout plants. * and ** indicate significant differences compared with WT (*P <0.05,**P<0.01), n=30. The same below.图 8 OsZAT12转基因水稻幼苗对ABA的敏感性分析Fig. 8 Sensitivity analysis of OsZAT12 transgenic rice seedlings to ABA
2.8 OsZAT12转基因植株农艺性状的观察与统计
农艺性状的统计分析结果表明,在分蘖期、抽穗期、成熟期这3个时期中,OsZAT12过表达水稻的株高均显著低于野生型,而OsZAT12敲除植株则与野生型没有显著差异(图9:A,B,D,F); 根长和分蘖数这2个指标,无论是OsZAT12过表达还是敲除植株,均与野生型无显著差异(图9:C,E,G)。OsZAT12敲除植株的每株穗数和结实率均显著低于野生型, 而OsZAT12过表达株系与野生型则无显著差异(图9:H,I)。以上结果表明,OsZAT12影响水稻的株高、每株穗数和结实率。
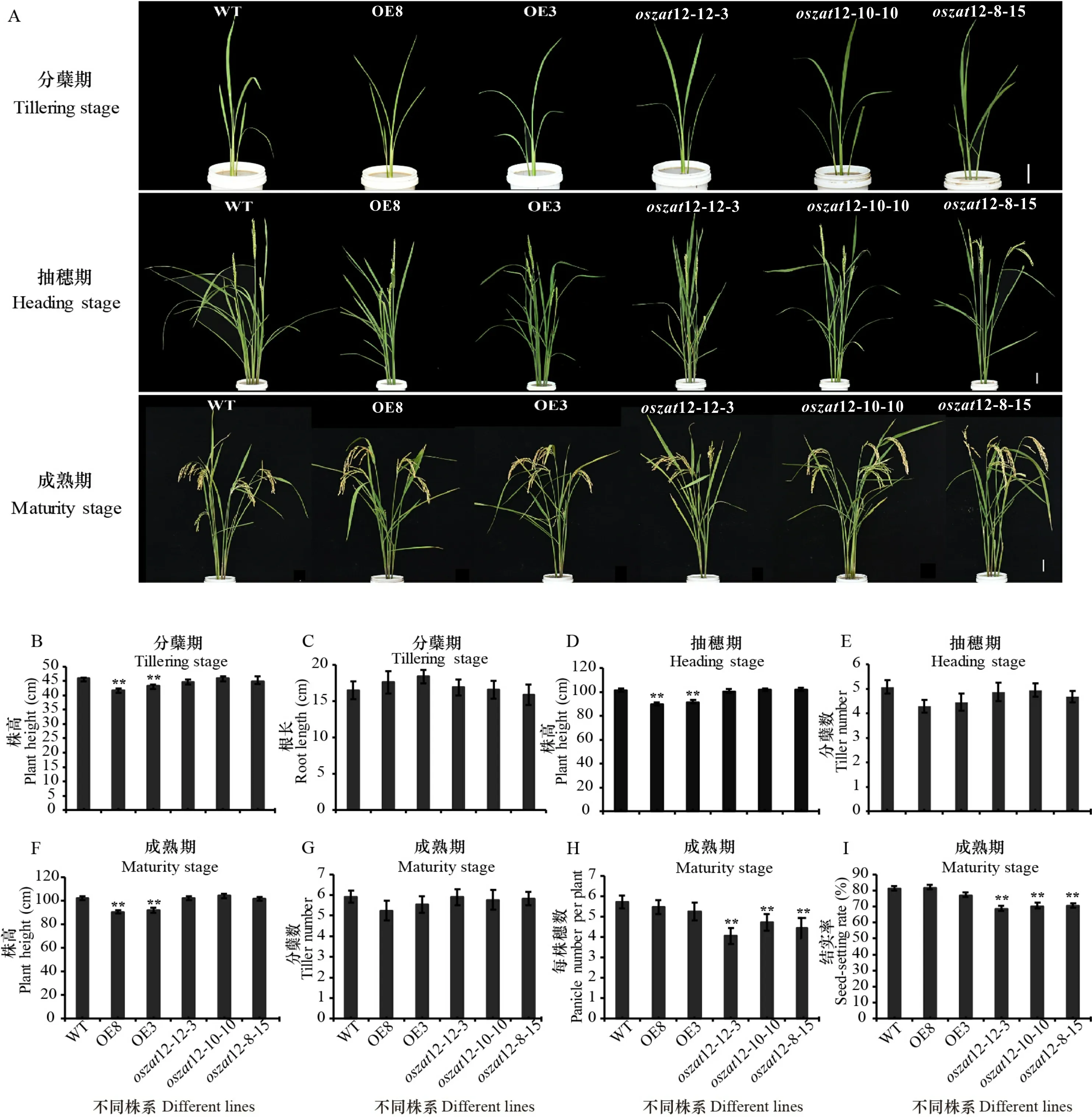
A. OsZAT12转基因水稻分蘖期、抽穗期和成熟期的表型,标尺=5 cm; B-C. OsZAT12转基因水稻分蘖期的株高和根长; D-E. OsZAT12转基因水稻抽穗期的株高和分蘖数; F-I. OsZAT12转基因水稻成熟期的株高、分蘖数、每株穗数和结实率。OE8和OE3为OsZAT12过表达植株; oszat12-12-3、oszat12-10-10和oszat12-8-15为OsZAT12敲除植株。A. Phenotype of OsZAT12 transgenic rice at tillering stage, heading stage and maturity stage, scale bar = 5 cm; B-C. Plant height and root length of OsZAT12 transgenic rice at tillering stage; D-E. Plant height and and tiller number of OsZAT12 transgenic rice at heading stage; F-I. Plant height, tiller number, panicle number per plant and seed-setting rate of OsZAT12 transgenic rice at maturity stage. OE8 and OE3 are OsZAT12 overexpression plants; oszat12-12-3, oszat12-10-10 and oszat12-8-15 are OsZAT12 knockout plants. 图 9 野生型和OsZAT12转基因水稻的农艺性状Fig. 9 Agronomic traits of wide type and OsZAT12 transgenic rice
3 讨论与结论
锌指蛋白是真核生物体内一类重要的转录调控因子家族,其中C2H2型锌指蛋白是最常见的一类(Takatsuji, 1999)。C2H2型锌指蛋白通常包含1~6个锌指结构域,并在锌指结构的α-螺旋中含有QALGGH保守序列(Sakamoto et al., 2000; Englbrecht et al., 2004; Ciftci-Yilmaz & Mittler, 2008; Wang et al., 2019)。本研究发现,水稻OsZAT12具有2个典型的C2H2锌指结构和1个EAR motif,并与拟南芥ZAT12有较高的同源性,说明OsZAT12属于水稻C2H2锌指蛋白家族成员。大多数含有EAR motif的锌指蛋白都表现出转录抑制活性(Ohta et al., 2001),如矮牵牛ZPT2-3的瞬时表达分析表明其起着阻遏物的作用(Sugano et al., 2003),而含有EAR motif like的拟南芥ZAT12,在响应冷胁迫时可能作为AtCBF转录因子的抑制子起作用(Vogel et al., 2005)。本研究结果表明,水稻OsZAT12蛋白具有转录抑制活性,是一个有功能的转录抑制因子。
植物C2H2型锌指蛋白作为一类重要的转录因子,是目前研究较为深入的一类锌指蛋白。该类转录因子在植物生长发育和响应非生物胁迫中具有重要调控作用(Sakamoto et al., 2004; Davletova et al., 2005; Mittler et al., 2006; Rossel et al., 2007; Xie et al., 2012; Shi et al., 2014; Chen et al., 2016; Yin et al., 2017; Ballerini et al., 2020; Yin et al., 2020; Zhang et al., 2021; Rodas et al., 2021)。油菜BnLATE(LATE FLOWERING)通过限制油菜角果中木质素的聚合减少角果的破裂(Tao et al., 2017)。水稻NSG1基因编码C2H2型锌指蛋白,与拟南芥SUP(SUPERMAN)、JAG(JAGGED)和NUB(NUBBIN)及水稻SL1(STAMENLESS1)的功能类似,参与调控水稻花发育(Dinneny et al., 2004; Ohno et al., 2004; Xiao et al., 2009; Zhuang et al., 2020)。组成型过表达AtZAT10的转基因拟南芥表现为生长阻滞(Sakamoto et al., 2004; Mittler et al., 2006)。本实验室前期研究(陈宗新等,2019)发现,异源过表达OsZAT12拟南芥植株矮小,根生长受到抑制。与异源转化OsZAT12的拟南芥表型相似,该研究也发现过表达OsZAT12水稻植株在分蘖期、抽穗期和成熟期的株高均显著降低。该类表型都类似于拟南芥或水稻等植株遭遇胁迫后或过表达胁迫相关转录因子如DREB1植株的表型(Kasuga et al., 1999),说明OsZAT12可能为胁迫相关基因。
植物对非生物胁迫的耐受性,主要依赖于激活植物体内与胁迫相关的分子调控网络,包括信号刺激的应答、激素信号转导途径、诱导信号通路基因的表达等(Dansana et al., 2014; Lima et al., 2015)。冷胁迫会对植物造成损伤,严重时可使植物死亡(Wang et al., 2017)。C2H2锌指蛋白可通过直接调控下游冷胁迫相关基因来增强植物的抗寒能力(Han et al., 2020)。番茄SlCZFP1基因通过诱导COR(cold-regulated)或冷响应相关基因的组成型表达,增强转基因拟南芥和水稻的耐寒性(Zhang et al., 2011)。大豆GmZF1通过结合COR6.6启动子区并上调该基因的表达,调节转基因拟南芥对冷胁迫的抗性(Yu et al., 2014)。在香蕉中,过表达MaC2H2-2和MaC2H2-3显著抑制MaICE1(inducer of CBF expression,冷信号传导途径的一个关键组成部分)的转录。因此,MaC2H2s可能通过抑制MaICE1的转录来增强香蕉的抗寒性(Han & Fu, 2019)。低温处理上调拟南芥ZAT12的表达(Davletova et al., 2005),过表达该基因则下调CBF(C-repeat/DREBindingFactor)基因的表达,表明ZAT12负调控拟南芥对冷胁迫的适应(Vogel et al., 2005)。本研究表明,4 ℃下调OsZAT12的表达,说明ZAT12在拟南芥和水稻中对低温胁迫的响应模式不同,暗示两者的功能可能不同。许多非生物胁迫(如盐、冷和干旱),可诱发植物产生渗透胁迫(Han et al., 2020)。渗透胁迫导致植物生理干旱、离子失衡、氧化损伤和生长抑制(Yamaguchi-Shinozaki & Shinozaki, 2006)。拟南芥ZAT10在渗透胁迫处理后表达显著上调,尤其是在叶片中;过表达ZAT10拟南芥和zat10突变体均表现出对渗透胁迫的耐受性增强(Mittler et al., 2006)。在拟南芥中,ZAT10被认为是渗透胁迫的正调控因子并受MAP激酶的调控(Nguyen et al., 2016)。杨树中鉴定出16个C1-2i亚家族成员,其中6个成员参与了渗透胁迫(Gourcilleau et al., 2011)。在拟南芥中异源过表达大豆GmZAT4,该基因可通过ABA途径提高拟南芥对20% PEG 6 000的耐受性(Sun et al., 2019)。在水稻中,20% PEG 6 000处理后RZF71的表达显著上调,表明该基因在渗透胁迫反应中发挥重要作用(Guo et al., 2007)。与拟南芥ZAT10和水稻RZF71的表达模式不同,本研究发现,在20% PEG 6 000的渗透胁迫处理过程中,OsZAT12的表达先下降,随后逐渐转变为上调趋势。综合以上结果表明,OsZAT12的表达受到非生物胁迫(如低温或渗透胁迫)的调控,因此推测OsZAT12参与了水稻响应非生物胁迫的过程。
本研究对水稻OsZAT12启动子分析发现其中含有激素相关元件,OsZAT12的表达在2,4-eBL、IAA处理后上调,推测OsZAT12参与了不同激素信号途径。ABA是植物非生物胁迫中一个重要的调控因子,由于其广泛在干旱、寒冷、高温、 盐胁迫和水涝等逆境中具有重要作用,因此被称为胁迫激素(Chen et al., 2020; Bharath et al., 2021)。水稻C1-2i亚家族成员的ZFP179,ABA处理3 h 后其表达上调,随后下调,至24 h 时达到最高(Sun et al., 2010),而本研究却发现,ABA显著下调OsZAT12的表达,说明两者在ABA信号传导途径中的功能可能存在不同。过表达ZFP179基因的水稻幼苗表现增加对ABA的敏感性(Sun et al., 2010),而本研究则发现过表达OsZAT12降低了水稻对ABA处理的敏感性。过表达OsZAT12水稻幼苗和过表达ZFP179水稻幼苗对外源ABA敏感性的差异可能与这两个基因对ABA的响应模式不同有关。此外,OsZAT12敲除植株的株高只有在5 μmol ·L-1ABA处理下显著低于野生型;在正常条件及低浓度ABA(1 μmol ·L-1)处理下,根长显著高于野生型;在较高浓度ABA(10 μmol ·L-1)处理下,根长与野生型无显著差异。我们推测,敲除OsZAT12后可能会降低植株内源ABA含量,只有在外施合适浓度的ABA时,敲除OsZAT12植株才会表现出对ABA的敏感性增强,这与osbglu33水稻突变体及过表达ZmWRKY114的水稻植株对外源ABA的响应方式相似(Ren et al., 2019; Bo et al., 2020)。结合OsZAT12对非生物胁迫(低温胁迫和渗透胁迫)及胁迫激素ABA的响应模式,推测OsZAT12参与调节非生物胁迫及激素信号途径,进而影响了水稻株型的发育,并在水稻穗型及结实中具有重要调控作用。本文进一步深入研究了OsZAT12参与植物不同非生物胁迫应答的分子机制和ABA信号转导途径的调控,将为利用OsZAT12进行水稻耐逆稳产分子设计育种提供实验依据。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
学与玩(2022年10期)2022-11-23 08:32:00
今日农业(2022年3期)2022-06-05 07:12:08
昆明医科大学学报(2022年1期)2022-02-28 07:46:10
实用临床医药杂志(2021年13期)2021-01-10 19:52:18
生命科学研究(2018年1期)2018-05-29 01:12:47
上海农业学报(2017年3期)2017-04-10 12:39:14
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
创新科技(2015年1期)2015-12-24 06:23:21
中华胰腺病杂志(2015年5期)2015-12-08 12:18:10
