
Cr6+对人工湿地薏苡光合特性和微量元素吸收的影响
2022-12-30 07:20:08柳心怡黄建祥李素丽李良香程夕冉王学礼李正文李志刚
广西植物 2022年11期
柳心怡, 农 宇, 黄建祥, 李素丽, 李良香, 程夕冉, 王学礼, 李正文, 李志刚
(广西大学 农学院, 南宁 530005 )
铬能够通过食物链途径进入植物、动物和人的体内,并进行积累,且极易对生态环境和人体健康造成伤害(Wenzel et al.,2003;Rajkumar et al.,2009;樊霆等,2013)。铬(Cr)在环境中的常见价态有Cr(Ⅲ)和Cr(Ⅵ),其中,Cr(Ⅲ)毒性较低,是人体必需的微量元素之一;而Cr(Ⅵ)通常以铬酸盐或重铬酸盐离子的形式与氧结合,其强氧化性、高水溶性和膜渗透性使得极低浓度的Cr(Ⅵ)也具有高毒性(王爱云等,2012;Alahmad et al.,2019)。铬广泛应用于电镀业和染料业等行业,这些行业工厂排放的废水是铬污染传播的重要途径,并会导致耕地污染。因此,加强含铬废水治理和达标排放是避免耕地铬污染和保护有限耕地资源的重要手段。
前人研究发现,运用化学固化法、化学稳定化法、化学还原法、离子交换法和微生物还原法等技术可以去除污水中铬,但这些技术存在成本较高、操作复杂和不适于低浓度Cr(Ⅵ)污染水体处理(伍清新等,2014;田小利和李倩,2017)等问题。人工湿地是植物-微生物联合修复的一种修复模式,因其成本低、耗能少和具有一定的生态效益的特点,被认为是处理重金属污染修复的有效途径(Rivastava et al.,2009; Zhuang et al.,2019;Xu et al.,2020)。人工湿地对含铬污水具有较好的净化效果,李志刚等(2010)研究发现,薏苡(Coixlacryma-jobi)人工湿地能够高效去除生活污水中低浓度Cr6+(<10 mg·L-1),且植物表现出相对较高的耐受性,而高浓度Cr6+(30 mg·L-1)胁迫则对薏苡的生长有显著的抑制作用。光合作用强弱是衡量植物生长状态的重要指标,铬含量过高会导致植物叶绿素分解,净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、蒸腾速率(transpiration rate,Tr)和胞间CO2浓度(intercelluar CO2concentration,Ci)等光合生理指标下降,最终导致植物的生长受到显著抑制(Choudhury et al.,2012;邵云等,2012)。其中,植物光合速率的下降可能导致光合电子传递过程中电子传递给O2分子,活性氧含量增加,从而引起膜脂过氧化,丙二醛(malondialdehyde, MDA)在体内过量积累,使得植物体的各类代谢活动受到严重阻碍甚至失调(杨雯一,2021)。植物可通过提高超氧化物歧化酶(superoxide dismutase, SOD)和过氧化物酶(peroxidase, POD)等抗氧化酶的活性来抵御重金属的毒害作用(钟旻依等,2019)。此外,植物体内产生的可溶性蛋白等渗透调节物质亦可缓解重金属胁迫(吕冬梅等,2021)。但是,有关铬胁迫条件下垂直流人工湿地植物光合参数、抗氧化酶活性的变化特征的研究并不多见。
植物光合能力的大小与矿质养分的吸收关系极为密切,Cu、Zn、Fe和Mn等微量元素与光合作用关系极为密切,重金属胁迫对植物的养分吸收和代谢产生重要影响,如Ali 等(2012)研究发现铬胁迫抑制大麦对Fe、Mn、Cu和Zn的吸收。肖家昶等(2021)研究发现在铝胁迫下,西瓜叶片钾、钙和镁元素积累,而钾、钙和镁元素与植物叶片光合和抗逆等功能密切相关。因为Fe、Zn、Mn和Cu都是植物正常生长和代谢所必需的微量营养元素,也是某些酶的活化剂,在植物的光合作用、呼吸作用等方面的氧化还原过程中具有非常重要的作用(徐根娣等,2015),所以随着这些元素吸收量的减少,将会直接导致植物光合能力的下降和生物量的减少等问题。迄今为止,有关Cr6+胁迫对植物吸收营养元素的影响的研究仍鲜见报道。因此,了解Cr6+胁迫下植物的养分元素变化对于深入了解人工湿地植物响应铬胁迫机制具有重要的实际意义。
本研究拟采用构建模拟垂直流薏苡人工湿地的方法,以含不同浓度Cr6+的铬废水(利用1/2 Hoagland’s营养液配制)作为人工湿地灌溉用水,拟探讨以下问题:(1)不同浓度Cr6+对人工湿地中薏苡吸收微量元素的影响;(2)不同浓度Cr6+人工湿地中薏苡生长、光合特性和人工湿地对Cr6+去除的关系。本研究结果可为实际人工湿地运作提供一定的理论依据,并为今后人工湿地修复含重金属污水提供参考价值和指导。
1 材料与方法
1.1 试验设计
本试验于2020年的5—8月在广西大学农学院科研基地网室(108°17′14″ E,22°50′17″ N)内进行。当地年平均气温为22.0 ℃,属亚热带季风气候,年均降雨量为1 300~2 000 mm。
薏苡是禾本科(Poaceae)、薏苡属(Coix)植物。薏苡为我国东南部常见栽培或野生植物,生于温暖潮湿的边地和山谷溪沟。薏苡是湿生性植物,适应性强,对土壤要求不严,可以在富营养化的水体中生长,根系发达,具有修复受污染水体能力(高冲,2008)。因此,本试验以薏苡为人工湿地植物,试验材料来自广西大学农学院试验基地。参考李志刚等(2010)的方法构建微型垂直流人工湿地系统,选取上、下口直径和高度分别为71、45、61 cm的塑料桶,于桶内从下至上依次装填高度约10 cm的鹅卵石(直径2~5 cm),铺垫40 cm厚的细沙层,在距桶底约10 cm处安装水龙头作为出水口。每个桶内种植6株株高20 cm且长势均匀的薏苡苗。在1/2 Hoagland’s营养液中添加K2Cr2O7以获得含0(对照,CK)、5、20、40 mg·L-1浓度Cr6+的污水,不同浓度每个处理重复3次。
Cr6+处理前20 d,用1/2 Hoagland’s营养液灌溉人工湿地。5月14日开始进行不同浓度Cr6+处理,参考李志刚等(2018)的方法,采取间歇式进水方式,进水后停留3 d,然后落干4 d。每7 d为一个周期,直至当年8月30日试验结束。
1.2 样品采集
分别于Cr6+处理后10、30、60、100 d进行样品采集。取出植株并将根、茎和叶洗净拭干,一部分叶片鲜样用于逆境生理指标的测定;另一部分把根、茎和叶分开,放入烘箱105 ℃杀青30 min,70 ℃烘至恒重后粉碎,最后过60目筛后装样保存待测。
1.3 测定方法
1.3.1 逆境生理指标的测定 叶片SOD活性采用苏州科铭试剂盒测定;POD活性测定参照李合生(2000),采用愈创木酚动力学扫描法;MDA含量测定参照刘大林等(2015),采用硫代巴比妥酸比色法。
1.3.2 光合生理指标的测定 分别于Cr6+处理后的第10、第30、第60 和第100 天的上午9:00—11:30使用LI-6400XTR光合作用测定仪(生产厂家:LI-COR,产地:美国),选择第一片展开叶(叶片基部出现叶环时即为展开叶),测定薏苡叶片的净光合速率、气孔导度、蒸腾速率和胞间CO2浓度指标。测定期间光照强度为(480±10 )μmol ·m-2,光照时间为(16±0.5) h,相对湿度保持在60%~70%。
1.3.3 微量元素铬、铁、铜、锰和锌含量的测定 将植株的根、茎和叶分别冲洗干净后,将根系浸泡于浓度为20 mmol·L-1的Na2-EDTA溶液中30 min,然后用超纯水冲洗干净,以除去吸附在根表面的铬。将根、茎和叶样品烘干后粉碎过100目筛。参照王爱云等(2012)的方法,用浓硝酸与高氯酸按4∶1(V/V=4∶1)的方法进行消煮,并用电感耦合等离子发射体发射光谱仪(型号:ICP-5000,生产厂家:北京聚光科技有限公司,产地:中国)测定各组分中的Cr、Fe、Cu、Mn和Zn元素含量。
1.3.4 农艺形状测定 分别于Cr6+处理后第10、第30、第60 和第100 天,测量薏苡茎粗、株高(从基部至顶部第一个可见叶环)和分蘖数。
1.3.5 水中Cr6+含量测定 参照苏长青(2016)的方法,使用二苯碳酰二肼分光光度法测定出水中Cr6+含量。其中所用比色管均用10%稀硝酸浸泡16 h以上,以防止试管内壁吸附Cr6+。
1.4 统计分析
采用软件Microsoft Excel 2010进行数据处理和计算,数据分析软件DPS 9.50进行统计分析,Duncan’s法进行显著性检验(P<0.05)。平均数据以“平均数 ± 标准差”表示。
2 结果与分析
2.1 不同浓度Cr6+对薏苡农艺性状的影响
由表1可知,各处理的薏苡株高随着Cr6+处理时间的延长呈现出提高的趋势,高浓度的Cr6+胁迫(20~40 mg·L-1)抑制薏苡植株生长。处理过程中5 mg·L-1Cr6+处理的薏苡株高与CK相比差异不显著,而20 mg·L-1和40 mg·L-1Cr6+处理均显著抑制薏苡的株高,表明Cr6+对薏苡的抑制随Cr6+浓度的增加而显著增强。
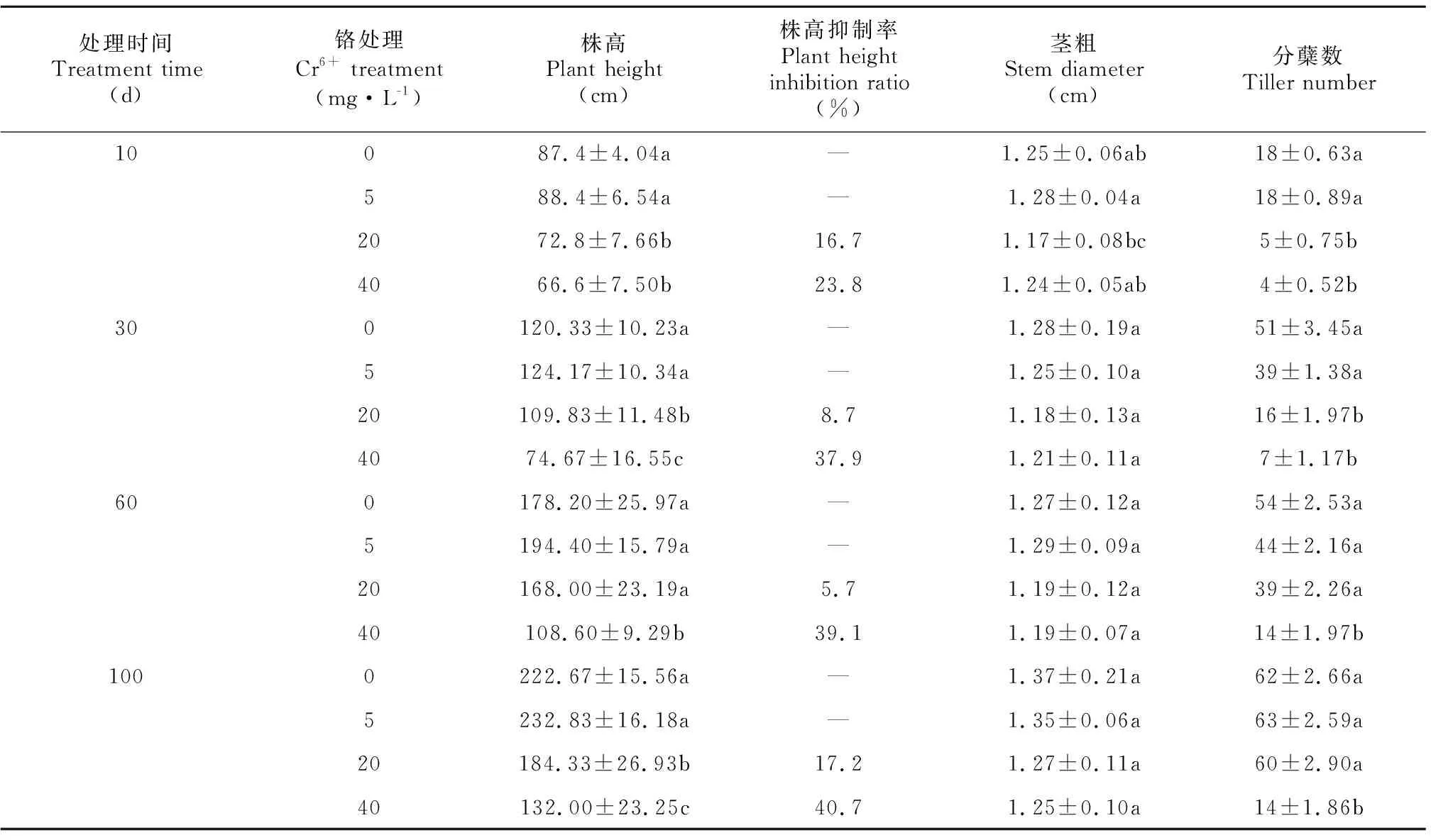
表 1 不同浓度Cr6+处理下薏苡株高、茎粗和分蘖数变化Table 1 Changes of plant height, stem diameter and tiller number of Coix lacryma-jobi treated with different concentrations of Cr6+
不同铬浓度下薏苡茎粗变化不明显,与CK差异不显著。分蘖数随Cr6+浓度的增加而减少。5 mg·L-1Cr6+处理可促进薏苡分蘖,20 mg·L-1和40 mg·L-1Cr6+处理均抑制薏苡分蘖,40 mg·L-1Cr6+处理与CK相比分蘖数差异显著。
2.2 不同处理下薏苡叶片光合作用的变化
由表2可知,薏苡叶片的Pn和Tr随Cr6+处理时间的延长而呈先升后降的趋势,Cr6+处理5 d,Pn和Tr值最高;5 mg·L-1Cr6+处理下,Pn和Tr均显著大于CK,Cr6+处理30 d,Pn和Tr达到最高,分别比CK高1.1和1.4倍,随后逐渐下降;不同Cr6+处理浓度相比较,5 mg·L-1Cr6+处理下,Pn和Tr值最高,而20、40 mg·L-1Cr6+处理下,Pn和Tr均显著小于CK,浓度越高,Pn和Tr值越低,分别比对照低6.1%~66.4%和28.2%~55.9%。
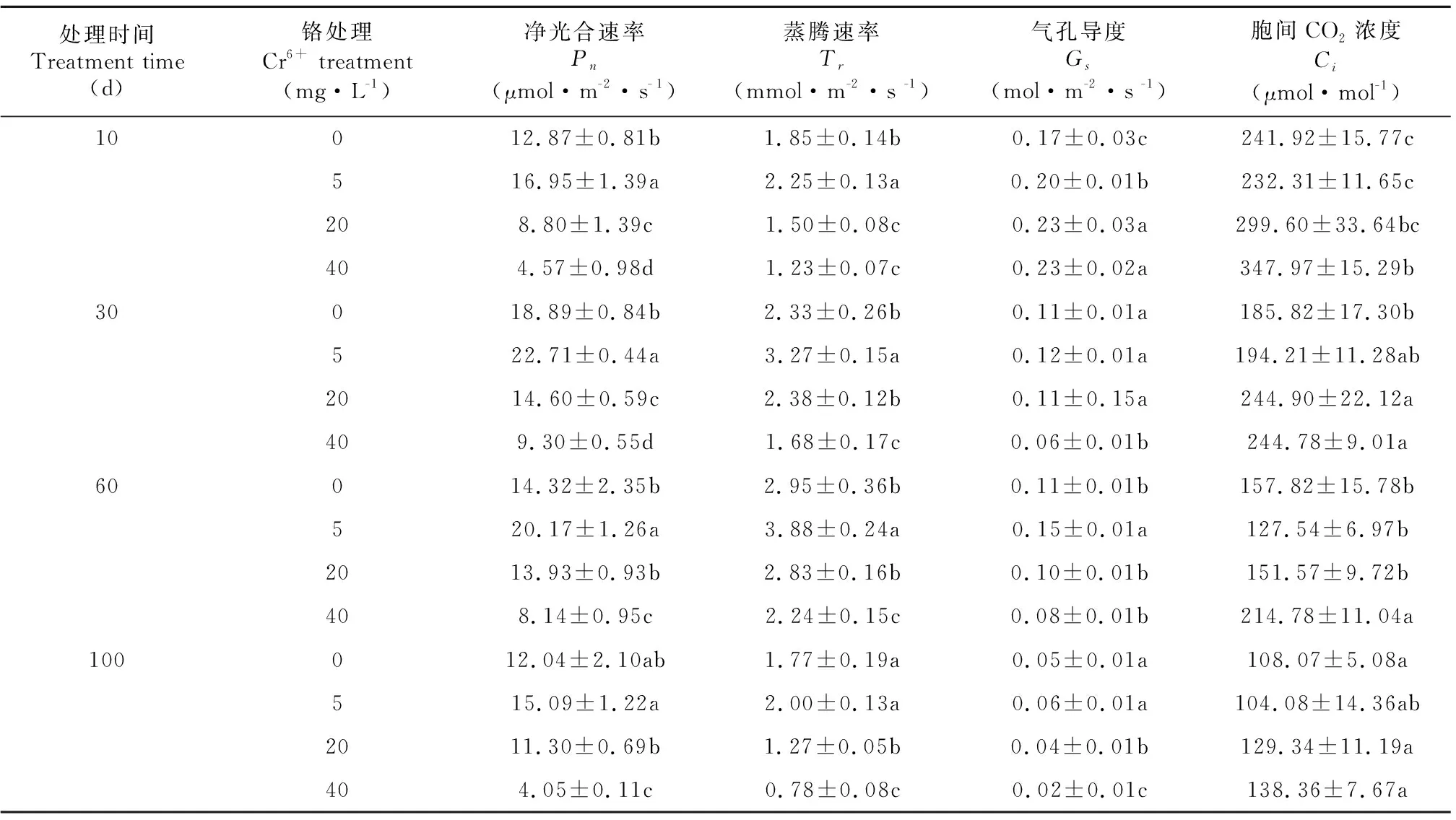
表 2 不同浓度Cr6+处理下薏苡叶片净光合速率、气孔导度、胞间CO2浓度和蒸腾速率的变化Table 2 Changes of Pn, Gs, Tr and Ci in the leaves of Coix lacryma-jobi treated with different concentrations of Cr6+
随处理时间的延长,CK的Gs呈无规律变化。5 mg·L-1Cr6+处理下,叶片Gs大于CK,提高幅度在16.7%~26.7%之间,而高浓度的Cr6+胁迫(20~40 mg·L-1)下,除Cr6+处理10 d时Gs大于对照外,20 mg·L-1Cr6+处理Gs与CK差别不大,而40 mg·L-1Cr6+处理Gs值则显著低于CK,降低幅度在27.2%~60%之间;与对照相比,5 mg·L-1Cr6+处理叶片Ci与CK差异不大,其余处理叶片Ci均显著大于CK,并随着Cr6+处理浓度的提高而提高。
2.3 不同处理下薏苡叶片逆境生理指标变化
2.3.1 不同处理下薏苡叶片SOD活性的变化 由图1可知,薏苡叶片的SOD活性在87.9~155.4 U·g-1之间。随Cr6+处理时间的延长,薏苡叶片SOD活性呈上升的趋势。5 mg·L-1Cr6+处理叶片SOD大于CK;20 mg·L-1Cr6+处理薏苡叶片SOD活性与CK差异不显著;而40 mg·L-1Cr6+随着处理时间的延长,表现出先升后降的趋势。

不同字母表示相同天数不同处理间差异显著性(P<0.05)。下同。Different letters indicate significant differences between different treatments for the same number of days (P<0.05). The same below.图 1 不同浓度Cr6+处理下薏苡叶片超氧化物歧化酶(SOD)活性变化Fig. 1 Changes of SOD activities in the leaves of Coix lacryma-jobi treated with different concentrations of Cr6+
2.3.2 不同处理下薏苡叶片POD活性的变化 由图2可知,薏苡叶片的POD活性在1 194.8~3 562.4 U·g-1·min-1之间,随Cr6+处理时间的延长和浓度的提高而提高。在5、20 mg·L-1Cr6+处理前期30 d内,POD活性与CK差异不显著,但随Cr6+处理时间的延长,Cr6+对叶片POD活性有显著的促进作用,Cr6+处理60~100 d,与CK相比,5、20 mg·L-1Cr6+处理叶片POD活性分别提高了27.8%~36.3%和36.9%~39.4%。40 mg·L-1Cr6+处理下,在Cr6+处理初期(10 d),叶片POD活性与CK差异不显著,但Cr6+处理30 d后,叶片POD活性显著大于其他浓度处理,提高了29.5%~40.6%和26.6%~31.3%。
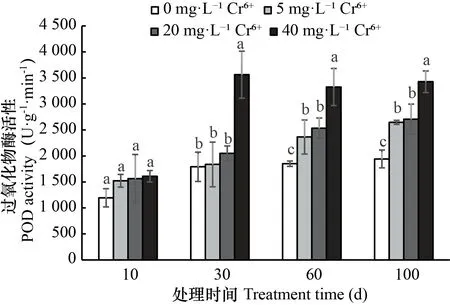
图 2 不同浓度Cr6+处理下薏苡叶片过氧化物酶(POD)活性变化Fig. 2 Changes of POD activities in the leaves of Coix lacryma-jobi treated with different concentrations of Cr6+
2.3.3 不同浓度Cr6+对薏苡叶片MDA含量的影响 由图3可知,薏苡叶片MDA含量在9.12~48.83 μmol·g-1之间。随Cr6+处理时间的延长,叶片MDA含量呈先升后降的趋势,Cr6+处理30 d时,各处理MDA含量达到最高值,30 d后呈下降趋势。5 mg·L-1Cr6+处理下,叶片MDA含量与CK相比差异不显著,而20、40 mg· L-1Cr6+处理叶片MDA的含量则显著大于CK及5 mg ·L-1Cr6+处理的。叶片MDA含量随Cr6+浓度增大呈上升。随处理时间的延长,不同浓度Cr6+处理叶片MDA含量差异逐渐缩小。
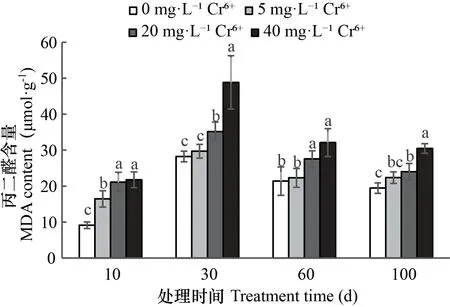
图 3 不同浓度Cr6+处理下薏苡叶片丙二醛(MDA)含量变化Fig. 3 Changes of MDA contents in the leaves of Coix lacryma-jobi treated with different concentrations of Cr6+
2.4 不同浓度Cr6+对薏苡吸收微量元素及铬的影响
由表3可知,薏苡植株根、茎、叶的Fe含量分别在450.36~1 996.30 mg·kg-1、39.37~172.44 mg·kg-1和198.33~382.81 mg·kg-1之间,不同部位Fe含量大小依次为根>叶>茎。处理30 d时,5 mg·L-1Cr6+处理根、茎、叶Fe含量均比处理10 d时有不同程度的增加,增加幅度为7.8%~29.2%,其中茎中Fe含量增加幅度最大,根次之,最低为叶,而20、40 mg·L-1Cr6+处理下,根、茎、叶中Fe含量均呈下降趋势,下降幅度最大的为20 mg·L-1Cr6+处理的根,下降幅度为24.6%;不同Cr6+处理浓度相比较,除5 mg·L-1Cr6+处理与CK差异不显著外,其他浓度Cr6+处理均显著降低了根系对Fe的吸收,与CK相比,降幅最大的为40 mg·L-1Cr6+处理,降幅为47.5%。
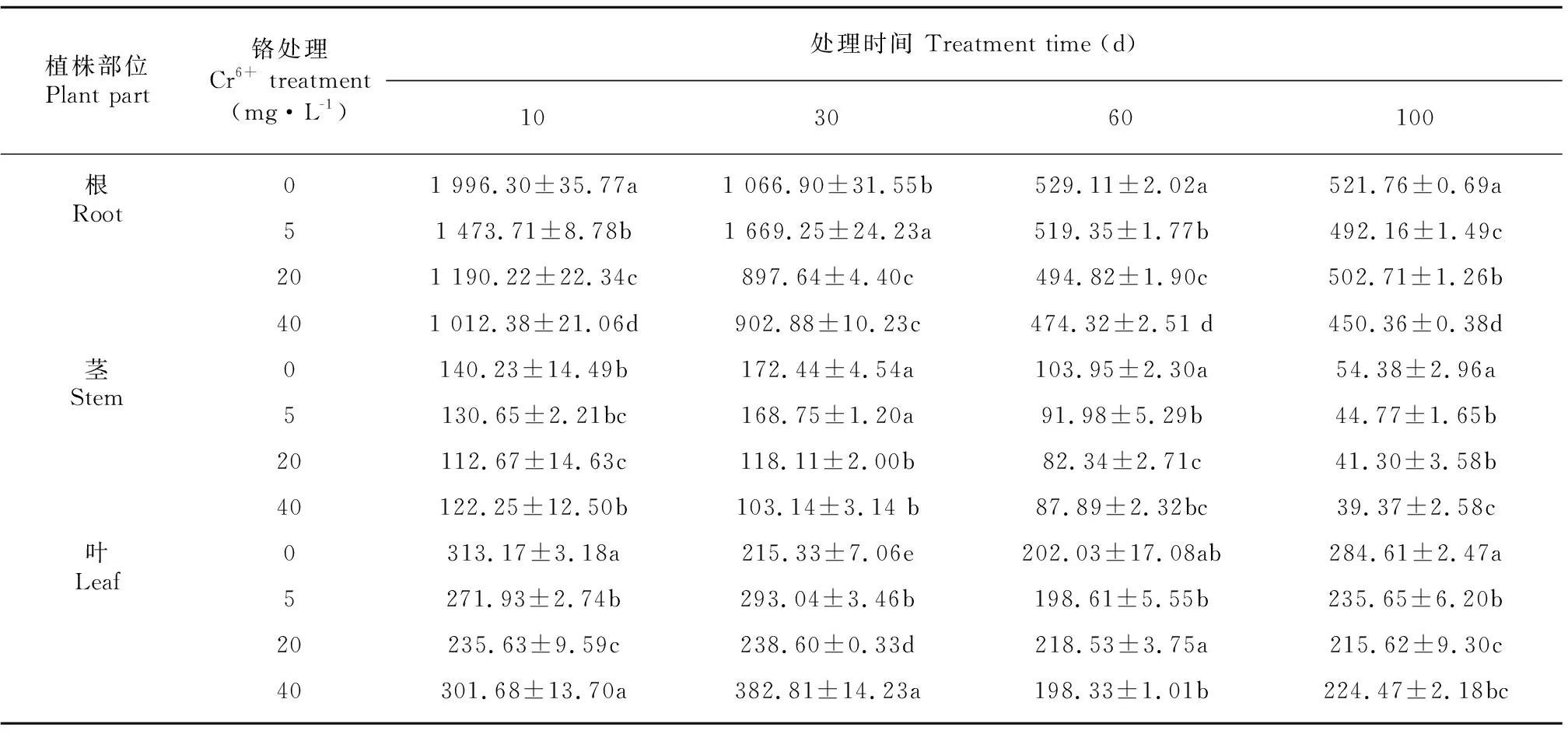
表 3 不同浓度Cr6+处理对薏苡根、茎、叶铁含量的影响(单位: mg·kg-1)Table 3 Effects of different concentrations of Cr6+ treatments on Fe contents in root, stem and leaf of Coix lacryma-jobi (Unit: mg·kg-1)
薏苡根系的Mn含量最高,其次是茎,最低的是叶(表4)。除CK和5 mg·L-1Cr6+处理30 d的茎Mn含量大于10 d处理外,随着处理时间的延长,其他各Cr6+处理下,薏苡根、茎、叶对Mn含量的吸收皆呈显著下降趋势。与CK相比,在20、40 mg·L-1Cr6+处理下,薏苡根系对于Mn的含量显著下降,下降幅度分别在4.4%~10.6%和20.0%~42.6%之间。整个处理过程中,Cr6+处理10、40、60 d薏苡叶片的Mn含量随Cr6+浓度升高而显著降低。
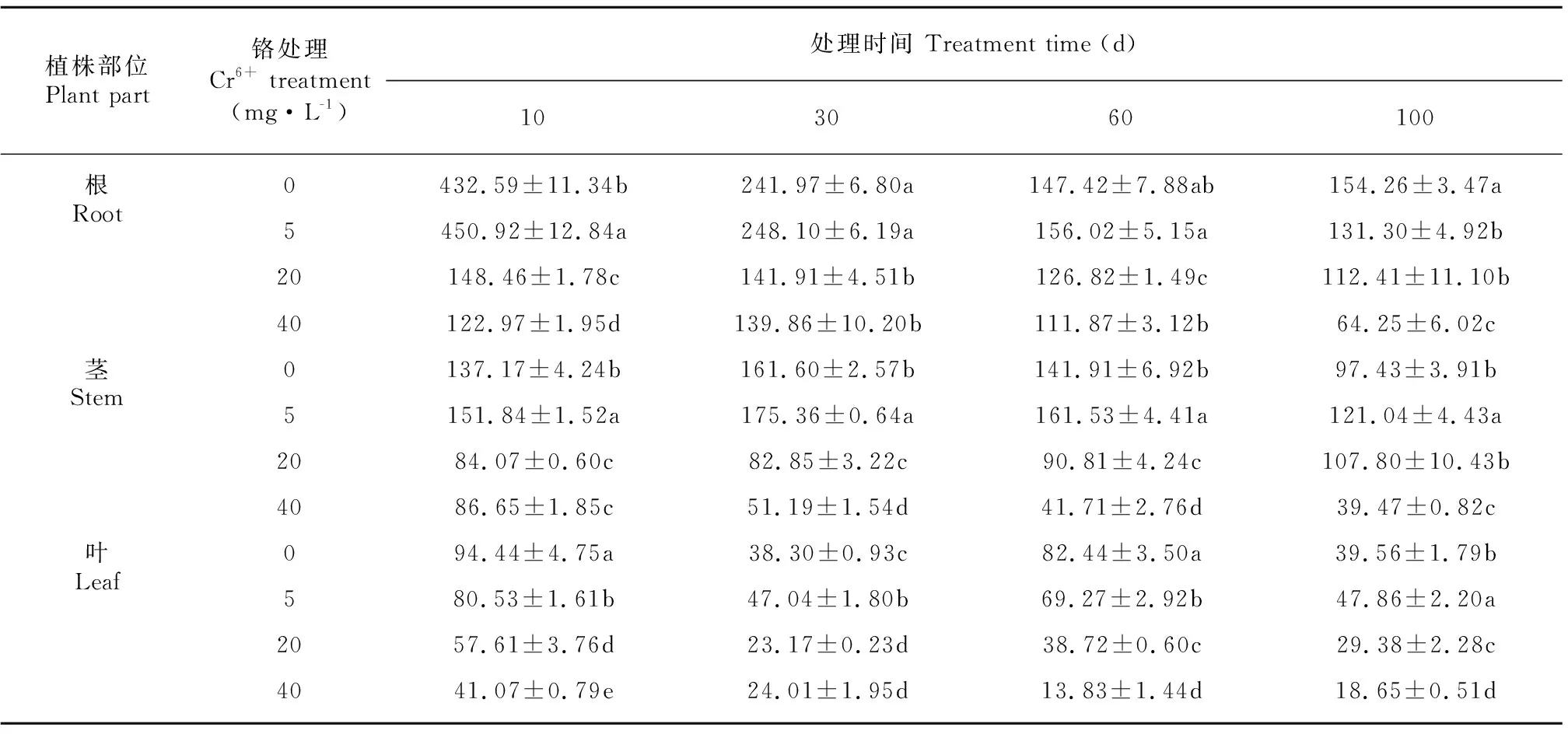
表 4 不同浓度Cr6+处理对薏苡根、茎、叶锰含量的影响(单位: mg·kg-1)Table 4 Effects of different concentrations of Cr6+ treatments on Mn contents in root, stem and leaf of Coix lacryma-jobi (Unit: mg·kg-1)
由表5可知,薏苡植株体内积累的Zn含量均随处理时间的延长而呈下降的趋势,不同部位Zn含量大小依次为根>叶>茎。茎Zn含量随铬处理浓度的提高而显著下降外,根和叶片Zn含量的变化没有一致的规律。
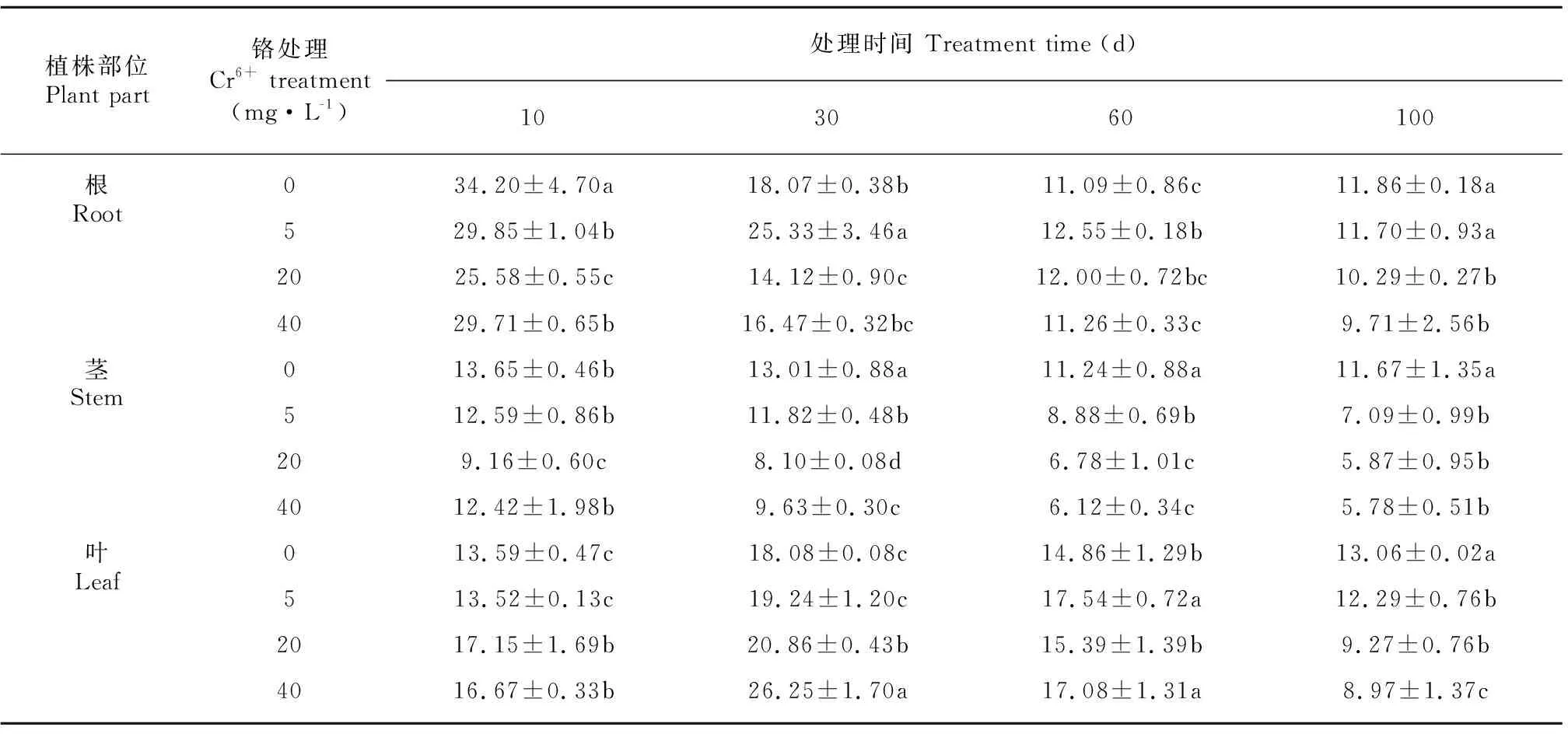
表 5 不同浓度 Cr6+处理对薏苡根、茎、叶锌含量的影响(单位: mg·kg-1)Table 5 Effects of different concentrations of Cr6+ treatments on Zn contents in root, stem and leaf of Coix lacryma-jobi (Unit: mg·kg-1)
薏苡植株根、茎和叶的Cu含量分别在4.69~15.83 mg·kg-1、 2.12~5.23 mg·kg-1、 2.89~5.29mg·kg-1之间,薏苡植株体内的Cu含量最高是根,其次是叶,最低是茎。薏苡植株体内积累的Cu含量均随处理时间的延长而呈下降趋势(表6)。
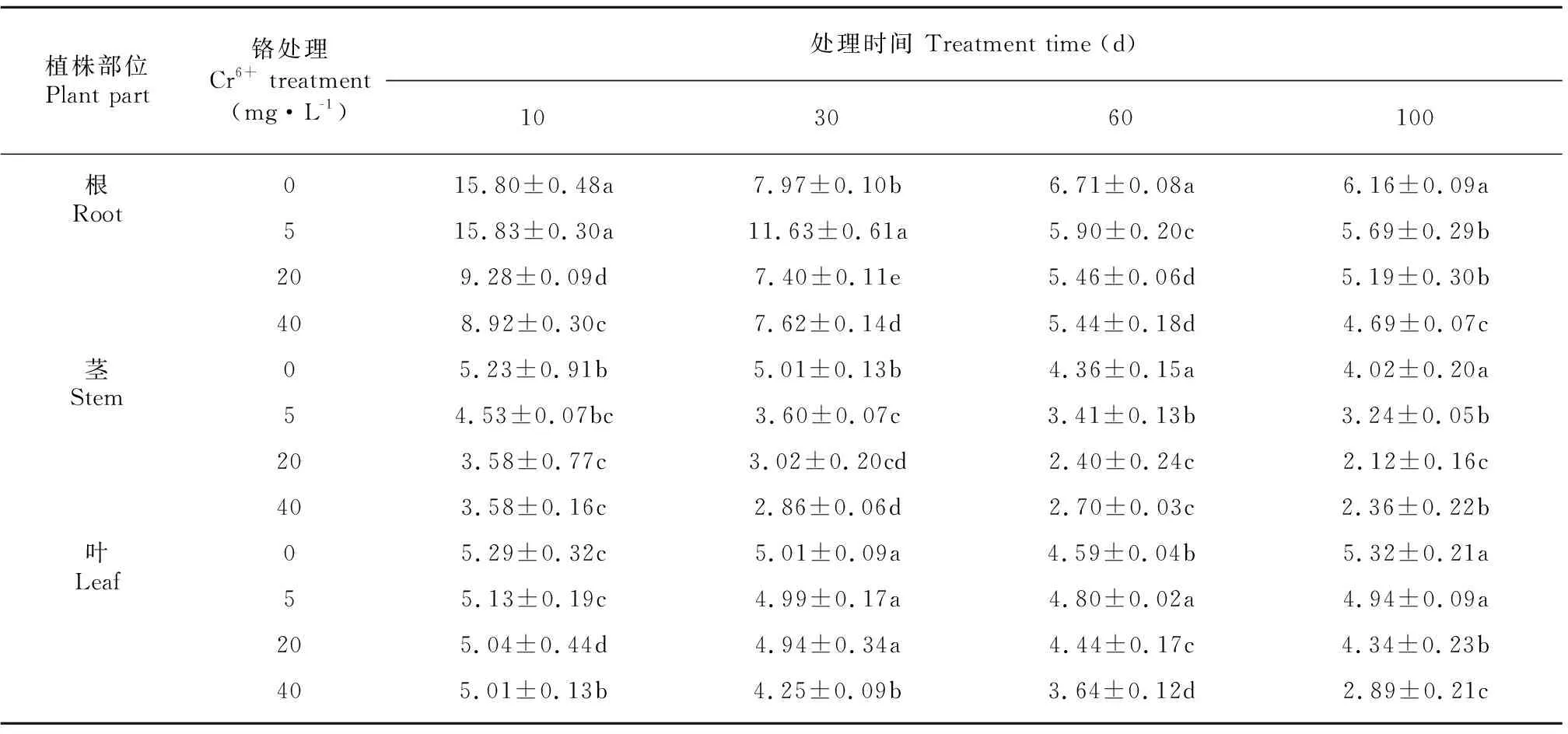
表 6 不同浓度Cr6+处理对薏苡根、茎、叶铜含量的影响(单位: mg·kg-1)Table 6 Effects of different concentrations of Cr6+ treatments on Cu contents in root, stem and leaf of Coix lacryma-jobi (Unit: mg·kg-1)
由表7可知,薏苡植株内积累的铬含量大小依次为根>叶>茎。薏苡根系铬的含量随Cr6+处理浓度的提高而增加,处理10 d和100 d,高浓度的Cr6+胁迫(20~40 mg·L-1)对薏苡根系的铬含量影响差异不显著,其他时期表现均为显著。在整个Cr6+处理过程中,40 mg·L-1Cr6+处理薏苡茎对于铬的吸收呈现出先升后降的趋势。处理30 d前,40 mg·L-1Cr6+薏苡叶片对铬含量的吸收呈上升趋势,与10 d相比增加了35.1%;而30 d促进作用却不明显,表现为下降的趋势,处理100 d与30 d相比薏苡叶片对铬含量的吸收下降了26.6%。
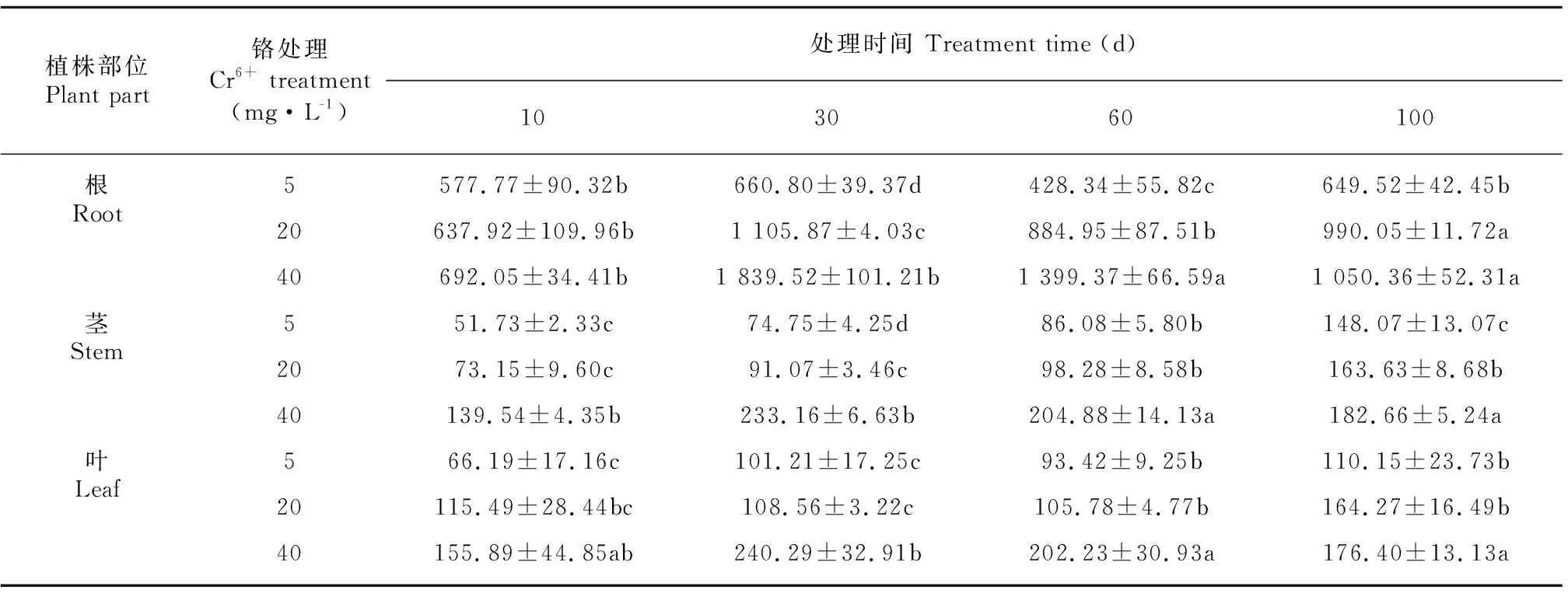
表 7 不同浓度Cr6+处理对薏苡根、茎、叶总铬含量的影响 (单位: mg·kg-1)Table 7 Effects of different concentrations of Cr6+ treatments on Cr contents in root, stem and leaf of Coix lacryma-jobi (Unit: mg·kg-1)
2.5 不同处理下出水Cr6+含量变化
由图4可知,5、20 mg·L-1Cr6+处理下湿地出水Cr6+含量极低,去除率最高可达99%,随铬处理时间的延长,两种人工湿地对Cr6+的去除率下降,但在处理后期去除率最低也高达89%。而40 mg·L-1Cr6+处理对Cr6+的去除率与5、20 mg·L-1Cr6+处理相比差异显著且去除效果不明显,其在30 d时达到最高,为86%,30 d后表现为下降趋势。
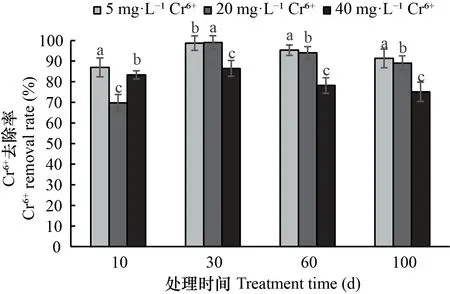
图 4 不同浓度Cr6+处理下出水Cr6+变化Fig. 4 Changes of Cr6+ in effluent treated with different concentrations of Cr6+
3 讨论
前人研究发现,低浓度铬胁迫可促进西兰花生长,而高浓度铬胁迫则会产生抑制作用(徐芬芬,2014)。本研究亦发现,低浓度5 mg·L-1Cr6+处理提高了薏苡株高;20 mg·L-1以上浓度 Cr6+处理则对薏苡株高有抑制作用,随Cr6+浓度提高,抑制效果明显,但Cr6+处理对薏苡的茎粗影响不大。薏苡总分蘖数随Cr6+处理浓度增加而减少,40 mg·L-1Cr6+抑制作用显著。
光合作用是绿色植物最基本和最重要的生命活动过程,光合速率、气孔导度、胞间CO2浓度等是衡量光合作用强度的重要指标。有研究表明,适量的铬可提高植物叶片叶绿素含量,提高植物光合能力,促进植物生长(Bonet et al.,1991; 欧阳峥嵘等,2010);而过量的铬胁迫会则造成植物藻细胞线粒体损伤,叶绿体破坏,使得植物的光合效率和呼吸作用降低,从而抑制植物生长,严重时甚至还会导致植物死亡(Lu et al.,2013;杨国远等,2014;王碧霞等,2016)。光合作用的限制酶主要受到CO2浓度的调控,而气孔导度受到CO2的影响(姚佳等,2015)。在本研究中,薏苡叶片Gs随铬处理时间的延长而下降,表明铬胁迫会引起叶片含水量降低,从而促使气孔关闭,导致Ci下降,进而引起光合速率降低。本研究发现,低浓度的5 mg·L-1Cr6+处理促进薏苡叶片Pn和Tr的提高,而高浓度40 mg·L-1Cr6+处理则表现出抑制作用,植物生长的表现与光合参数的变化一致,表明高浓度Cr6+胁迫可能是通过抑制薏苡的光合作用,引导光合机构受损、光化学活性降低,从而产生明显的光抑制,最终导致植物生长受到抑制。
重金属对植物的毒害机理之一是干扰植物对养分的吸收,破坏植物体内的养分平衡(董钻和谢甫练,1996)。Fe、Mn、Zn和Cu是植物体内参与代谢作用的酶类辅基或激化酶活性的活化剂,参加代谢氧化还原过程,影响着植物呼吸作用、光合作用的过程。同时,还能提高作物对逆境和病害的抗性(陈永林,2016)。本研究发现,高浓度的Cr6+胁迫(20~40 mg·L-1)显著抑制薏苡根、茎和叶对Fe、Mn和Zn的吸收,而且抑制效果随着处理时间的延长而增加。铬胁迫干扰了植物对营养元素的吸收与运输,其原因可能是大量的活性离子在根尖细胞壁与质膜表面的阴离子之间相互作用,改变了其质膜结构以及膜电位,从而使得离子转运体的活性发生了变化,而在土壤中溶出的不同形态活性铬也降低了多价阳离子在根皮层细胞质外体的负载量,从而影响吸收;此外,也有可能是铬离子与金属阳离子竞争阳离子结合位点,使得其他营养元素的结合位点减少,从而抑制Fe、Mn、Zn等营养元素的吸收。高浓度铬胁迫使植物清除氧自由基和光合能力下降,造成植物氧化损伤,从而导致运输至根系光合产物量下降,根系因得不到足量的物质和能量供应,使其根系吸收元素能力下降,进而导致植物的代谢紊乱(徐根娣等,2015)。
Fe、Mn和Zn是植物正常生命活动所必需的微量元素,在植物体内参与调控叶绿素合成,调节植物体内CO2的供应和基质中的pH,从而影响到植物的光合作用和氧化还原反应等重要生理过程(杜新民和张永清,2008)。过量的Cr6+导致Fe、Mn和Zn吸收量的下降,可能导致薏苡光合速率和抗氧化系统对活性氧的调节能力下降,最终使得薏苡的生长受到抑制。
在重金属胁迫下,植物细胞内氧化还原平衡被破坏,从而导致植株中活性氧含量明显增加,影响了整个活性氧清除系统对活性氧的清除能力,导致整个生理生化过程紊乱, 从而进一步加重氧化损伤,影响植物生长(周希琴和莫灿坤,2003)。在本研究中的Cr6+处理下,薏苡叶片MDA含量随铬浓度的增大和时间的延长而呈现先升后降的趋势,叶片MDA含量提高,说明Cr6+胁迫下细胞膜脂过氧化水平高,膜结构受损伤程度加深,植物的抗逆性减弱(陈晶等,2017)。SOD和POD是植物体内重要的抗氧化酶,协助植物清除氧自由基,抵抗氧化胁迫(曾凡荣,2010)。本研究发现,在5 mg·L-1Cr6+处理下,薏苡叶片SOD和POD活性显著增强,有利清除氧自由基,减少叶片中MDA的积累,从而提高薏苡抵抗铬胁迫能力;而40 mg·L-1Cr6+高浓度铬胁迫对薏苡造成了严重的氧化胁迫,脂质过氧化程度增强,已超出了薏苡SOD和POD清除氧自由基抵抗氧化胁迫的能力,尽管此时SOD仍具有较高活性,但可能由于活性氧生成量过多,SOD来不及清除,从而使MDA生成量提高,因此40 mg·L-1Cr6+胁迫下薏苡的抗逆性降低,使其生长受到抑制。这一结果与汤茜等(2018)和朱秀红等(2017)的研究结果一致。说明Cr6+胁迫影响了植株的抗氧化系统,并且植株对外源抗氧化胁迫迅速做出了反应。
人工湿地对污水中Cr(Ⅵ)的去除与基质的吸附、植物吸收和微生物的活动密切相关,而植物的活动直接影响到人工湿地氧化还原条件、有机质分配和微生物活动,从而影响到人工湿地对Cr(Ⅵ)去除,人工湿地植物的生长状况与其对重金属的去除能力有密切的关系(Vymazal & Brezinova,2016)。本研究中,低浓度Cr6+胁迫下,人工湿地出水所含Cr6+含量较低,对Cr6+去除能力较强,可能是低浓度Cr6+胁迫下薏苡人工湿地能够维持相对较好的长势,对Cr6+去除率较高的重要原因。
4 结论
综上所述,本研究发现高浓度Cr6+铬胁迫下,薏苡植株体内的Fe、Mn、Zn和Cu等与光合和抗氧化等生理功能密切相关的微量元素吸收量大幅度下降,导致薏苡光合速率下降,抗氧化系统受损,从而使薏苡的生长受到抑制,最终导致人工湿地处理含Cr6+废水的能力下降。但是,低浓度Cr6+胁迫下的薏苡人工湿地能够维持相对较好的长势,且人工湿地对Cr6+去除效率较高。
猜你喜欢
广东第二课堂·小学(2023年5期)2023-05-29 02:16:13
现代农业科技(2022年14期)2022-12-14 14:16:21
天天爱科学(2022年12期)2022-11-10 08:33:28
军事文摘(2022年8期)2022-11-03 14:22:01
小学科学(学生版)(2021年3期)2021-04-13 08:26:18
哈哈画报(2021年11期)2021-02-28 07:28:45
装备制造技术(2020年1期)2020-12-25 05:19:10
农技服务(2020年2期)2020-05-20 09:55:12
种子(2019年2期)2019-03-21 07:28:04
中华老年口腔医学杂志(2016年1期)2017-01-15 14:24:42
