
矮化香蕉及其野生型GA3ox基因的结构特点和表达分析
2022-12-30 06:58林佳琦李燕培肖世祥禤维言
广西植物 2022年11期
林佳琦, 李燕培, 肖世祥, 冯 斗, 禤维言
(广西大学 农学院, 南宁 530005 )
香蕉是芭蕉科的高大草本植物,主要分布于热带和亚热带地区。香蕉产业是种植地区和国家的重要经济来源之一(Singh et al., 2016),在我国华南地区香蕉产业是仅次于柑橘的第二大重要水果产业。在生产过程中,尤其是在沿海地区,由于香蕉植株高大、冠幅重、抗风性能差,遭遇台风或热带风暴危害时容易出现严重的倒伏现象,使香蕉产业遭受重大损失(舒海燕等,2016)。株高是影响作物抗倒伏和丰产性能的重要农艺性状,作物越高、冠幅越大其抗倒伏性能越差(张瑞茂等,2019)。因此,筛选和创制优良的矮化新品种和新种质是当前育种的重要目标。然而,目前由于香蕉的栽培品种和野生资源多数是三倍体或四倍体,高度不育,很难通过杂交技术改良矮化其株高性状。但是,通过转基因或基因编辑技术可以改良或创制矮化的香蕉新品系或新种质(崔霞和张率斌,2017;王福军和赵开军,2018;李树磊等,2020);而采用转基因或基因编辑技术途径进行香蕉株高矮化方面的分子育种,首先必须研究清楚香蕉株高生长调控的机制和挖掘与香蕉矮化相关的关键调控基因。
植物的株高生长主要受内源激素影响,其中赤霉素(gibberellin,GA)是植物生长发育过程中对株高伸长生长影响最大的激素(杨益善等,2015;范业赓等,2019)。目前,已鉴定的赤霉素有136种,其中具有生物活性的赤霉素有GA1、GA3、GA4、GA7(Hedden & Thomas, 2012)。这些具有生物活性的赤霉素在植物生长发育各个阶段都发挥着重要作用,如叶片伸展、茎的伸长、果实发育等(Hu et al., 2018)。许多植物的株高受赤霉素(GAs)生物合成和代谢过程中相关酶基因的调控,如古巴焦磷酸合成酶基因(copay diphospate synthase,CPS)、内根-贝壳杉烯合成酶基因(kaurene synthase,KS)、内根-贝壳杉烯氧化酶基因(kaurene oxidase,KO)、GA3氧化酶基因(GA3 oxidation enzyme,GA3ox)和GA2氧化酶基因(GA2 oxidation enzyme,GA2ox)等(Hedden & Pnillips, 2000; Yamaguchi, 2008)。目前,大多数赤霉素合成途径的关键酶基因在多种植物中已被研究和鉴定,如在拟南芥(Helliwel et al., 1998)、玉米(Chen et al., 2014)、水稻(Ashikari et al., 2002; Sasaki et al., 2002)和豌豆(Ait-Ali et al., 1997; Davidson et al., 2004)等植物中都发现了许多由于赤霉素合成途径中代谢酶基因的变异导致矮化表型的相关植株。因此,赤霉素生物合成途径关键酶基因的研究在植物矮化机制中十分重要。
GA3-氧化酶(GA3ox)是活性赤霉素代谢途径中最后步骤的关键酶,是由多基因家族编码的双加氧酶,其功能是介导一个3β-羟基基团到GA9和GA20上,将无生物活性的GA9和GA20催化形成具有生物活性的GA1、GA4和GAs(Yamaguchi, 2008;陈晶晶等,2014a)。若GA3ox基因发生突变,使植物不能合成具有活性的赤霉素,植物株高生长受到抑制,从而导致矮化表型出现。此外,在许多矮化突变体中发现植物株高性状与GA3ox基因的表达水平密切相关,当GA3ox基因的表达水平受抑制时,植株会出现矮化性状(Roumeliotis et al., 2013)。因此,研究GA3ox基因结构变化及其表达特点与香蕉茎杆矮化的关系是揭示香蕉矮化变异分子机制的重要内容,对于挖掘调控香蕉株高关键基因及其应用具有重要的研究意义。在矮化变异的香蕉突变体苗期,其假茎GA1和GA3含量显著低于野生型亲本,并且外源GA3和吲哚-3-乙酸(indole-3-acetic,IAA)能使其株高恢复到野生型高度(陈晶晶等,2014b)。但是,香蕉的矮化变异是否与GA3ox基因的结构和表达水平改变有关,目前尚未见有研究报道。
本研究以威廉斯B6矮化突变体及其野生型亲本为材料,通过RT-PCR技术克隆得到矮化香蕉及其野生型亲本GA3ox基因的全长cDNA序列,并利用qRT-PCR 技术对GA3ox基因在不同组织中的表达差异进行分析。拟探讨以下问题:(1)矮化香蕉和野生型香蕉GA3ox蛋白同源性、理化性质分析;(2)矮化香蕉和野生型香蕉GA3ox氨基酸序列的结构差异分析;(3)GA3ox基因在矮化香蕉及野生型香蕉不同组织中的表达水平差异分析。本研究结果旨在为揭示香蕉矮化突变的分子机制与筛选优良矮化香蕉株系奠定基础。
1 材料与方法
1.1 材料
以威廉斯B6矮化突变体及其野生型亲本为试验材料,均采自于广西大学农学院农科基地温网室。选取长势一致、无病虫害的矮化突变体及其亲本香蕉树各2株,分别采集香蕉生长发育前期嫩叶、假茎样品用于基因克隆及表达分析。所有材料采集后立即放入液氮中冻存,-80 ℃冰箱保存备用。
1.2 方法
1.2.1GA3ox基因克隆与测序 采用TIANGEN多糖多酚植物总RNA提取试剂盒,提取香蕉幼嫩叶片中的总RNA。按照M-MuLV第一链cDNA合成试剂盒说明书将完整度较好、纯度较高的RNA反转成cDNA第一链,以cDNA为模板,根据NCBI上发表的Musaacuminatasubsp.malaccensis的GA3ox编码基因的ORF(XM_009398371.2)设计1对特异引物,此引物序列为GA3ox-F:5′-ATGAATCCCAATCCAACGAC-3′,GA3ox-R:5′-TTAACAACAAATCCCTTCG-3′。采用高保真taq酶进行RT-PCR扩增,PCR反应程序:95 ℃ 3 min;95 ℃ 15 s,55 ℃ 15 s,72 ℃ 5 min,35个循环;72 ℃延伸10 min,4 ℃保存。扩增产物经1%琼脂糖凝胶电泳检测正确后,使用SanPrep柱式DNA胶回收试剂盒将目的片段回收纯化,将回收产物与 PUCI-Blunt克隆载体相连接后,送至上海生工生物公司测序。
1.2.2GA3ox基因表达分析 利用qRT-PCR分析GA3ox基因在矮化突变体及野生型香蕉不同组织中的表达模式。分别采集矮化香蕉和野生型香蕉第10片、第15片、第20片、第25片叶龄期嫩叶及对应时期的假茎,液氮速冻后提取RNA;利用PrimeScriptTMRT reagent Kit with gDNA Eraser将RNA转化为cDNA,并根据克隆得到的矮化香蕉和野生型香蕉的GA3ox全长cDNA序列设计特异性引物,此引物序列为GA3ox-qF:5′-CTGGATCACGCCCTCAAGCTC-3′和GA3ox-qR:5′-TCAACTGCAACACGGCGGACA-3′,扩增片段长度为203 bp;以香蕉Actin作为内参基因,在GenBank登录号为AB022041,引物序列为Actin-F:5′-GCCATACAGTGCCAATCTACGAGG-3′和Actin-R:5′-ATGTCACGAACAATTTCCCGCTCA-3′,扩增片段长度为157 bp,进行基因的表达定量分析。采用2XUniversal SYBR GreenFast qPCR Mix染料说明书进行操作,qRT-PCR流程第一步为95 ℃ 3 min;第二步为95 ℃ 5 s,60 ℃ 34 s,40个循环;第三步熔解曲线的绘制为95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s。每个反应重复3次,结果采用2-△△CT方法计算基因的相对表达量。
1.2.3 序列分析 利用NCBI数据库的Blastp进行GA3ox氨基酸序列相似性分析;通过ExPASy在线软件(https://web.expasy.org/protparam)对GA3ox蛋白的分子质量、等电点、蛋白质稳定性等理化性质进行分析和预测;利用 SignalP(http://www.cbs.dtu.dk/services/SignalP)和TMHMM Server V.2.0(http://www.cbs.dtu.dk/services/TMHMM)分析GA3ox氨基酸序列的跨膜结构和信号肽。
2 结果与分析
2.1 香蕉矮化突变体与野生型GA3ox基因的序列比对分析
以矮化香蕉和野生型香蕉嫩叶的cDNA为模板,利用primer 5.0软件设计GA3ox基因的特异性引物进行PCR扩增。扩增产物通过1%琼脂糖凝胶电泳进行检测,得到两条1 000 bp左右的特异性条带(图1),测序后这2个靶序列的全长均为1 096 bp,将获得矮化突变体GA3ox基因命名为GA3ox-A、野生型香蕉GA3ox基因命名为GA3ox-G。通过测序得到GA3ox-A和GA3ox-G的全长序列均为1 096 bp,GA3ox-A和GA3ox-G的ORF均为864 bp,5′-UTR均为232 bp,均编码287个氨基酸。

M. DL5000 Maker; A. GA3ox-A; B. GA3ox-G。图 1 矮化香蕉与野生型香蕉 GA3ox基因PCR扩增产物Fig. 1 PCR amplification products of the GA3ox gene from dwarf banana and wild type banana
对GA3ox-A和GA3ox-G的氨基酸序列比对分析结果显示,两条氨基酸序列之间的同一性高达98.26%,存在5个位点的差异,分别位于第29位、第38位、第84位、第196位和第259位(图2)。
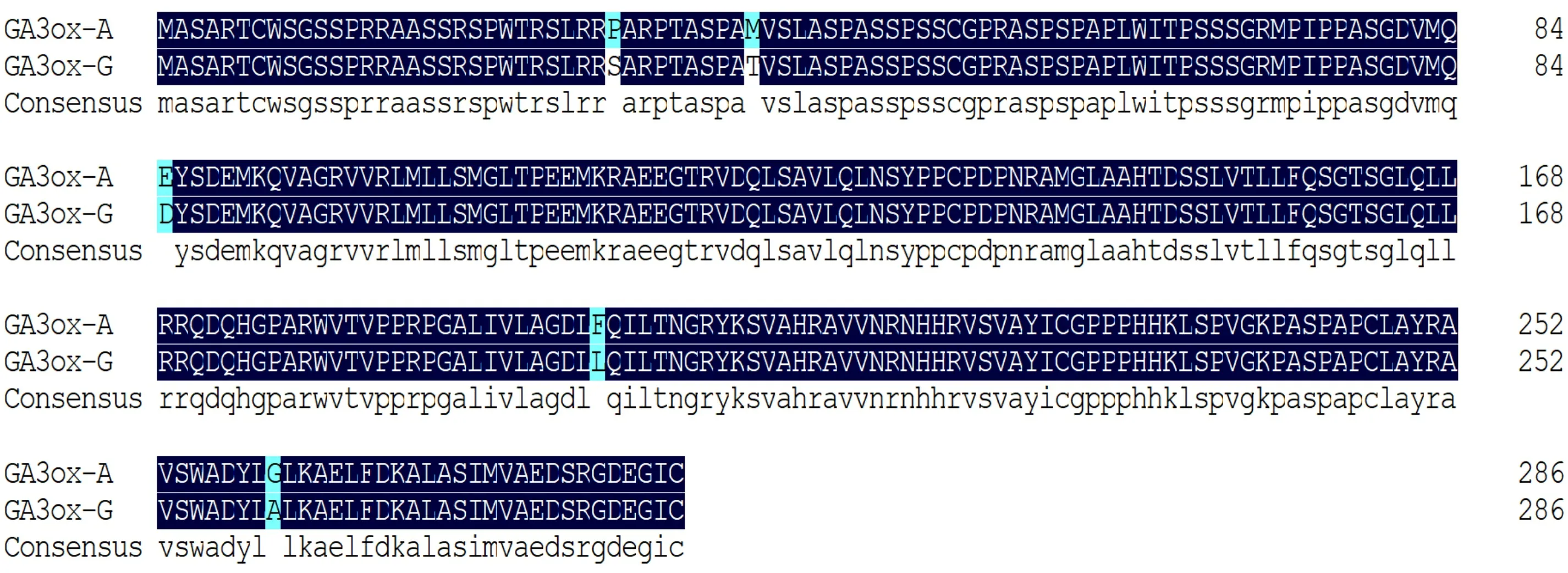
图 2 矮化香蕉与野生型香蕉GA3ox氨基酸序列比对分析Fig. 2 Comparison and analysis of GA3ox amino acid sequence from dwarf banana and wild type banana
2.2 香蕉矮化突变体与野生型GA3ox蛋白质理化性质及保守结构域分析
矮化突变体与野生型香蕉GA3ox氨基酸序列的理化性质分析结果显示,GA3ox-A和GA3ox-G分子式分别为C1338H2152N402O395S17和C1332H2150N402O397S16,蛋白分子量分别为30 735.28 Da和30 661.14 Da,理论等电点均为9.78。其中,GA3ox-A和GA3ox-G蛋白负电荷的残基总数(Asp+Glu)均为20个,正电荷的残基总数(Arg+Lys)均为32个;GA3ox-A和GA3ox-G蛋白的不稳定指数分别为 67.46和68.79,均属于不稳定蛋白;脂肪指数分别为76.90和78.61;亲水平均系数分别为-0.240和-0.236。根据 NCBI 在线软件对编码蛋白的结构域分析发现,GA3ox-G和GA3ox-A具有2-酮戊二酸依赖性的双加氧酶与Fe2+结合的保守结构域,与其他植物的GA3ox蛋白相同。
2.3 香蕉矮化突变体与野生型GA3ox蛋白质磷酸位点及其高级结构预测分析
蛋白磷酸位点分析结果显示,GA3ox-A和GA3ox-G蛋白均含有丝氨酸、苏氨酸和酪氨酸3种氨基酸磷酸化位点,GA3ox-A和GA3ox-G分别含有丝氨酸22个和24个,苏氨酸7个和8个,酪氨酸2个和4个。跨膜结构分析显示GA3ox-A和GA3ox-G氨基酸序列的期望值分别为0.037 29和0.028 98,均无跨膜区域。
利用SOPMA在线软件对 GA3ox-A和GA3ox-G蛋白的二级结构的预测分析结果显示,GA3ox-G蛋白含有4种构象,包括28.57%的α-螺旋、16.38%的延伸链、3.48%的β-转角和51.57%的无规则卷曲(图3);GA3ox-A蛋白含有4种构象,分别为26.13%的α-螺旋、17.42%的延伸链、3.48%的β-转角和52.96%的无规则卷曲(图4)。
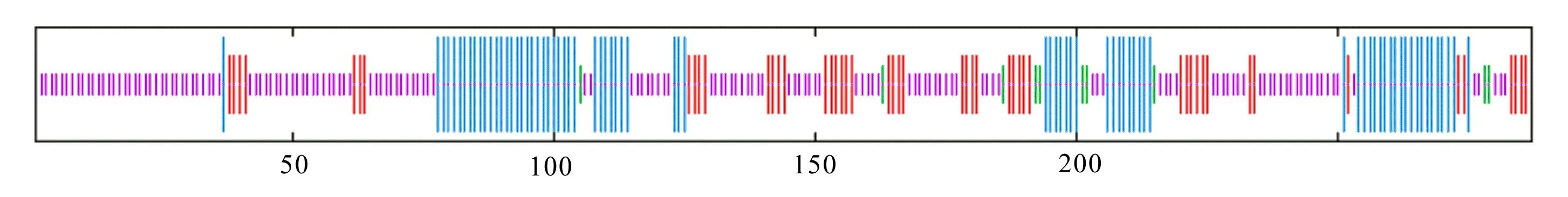
红色. 延伸链; 蓝色. α-螺旋; 紫色. 无规则卷曲; 绿色. β-转角。下同。Red. Extended chain; Blue. α-helix; Purple. Random coil; Green. β-turn. The same below.图 3 GA3ox-G蛋白二级结构预测Fig. 3 Secondary structure prediction of GA3ox-G protein
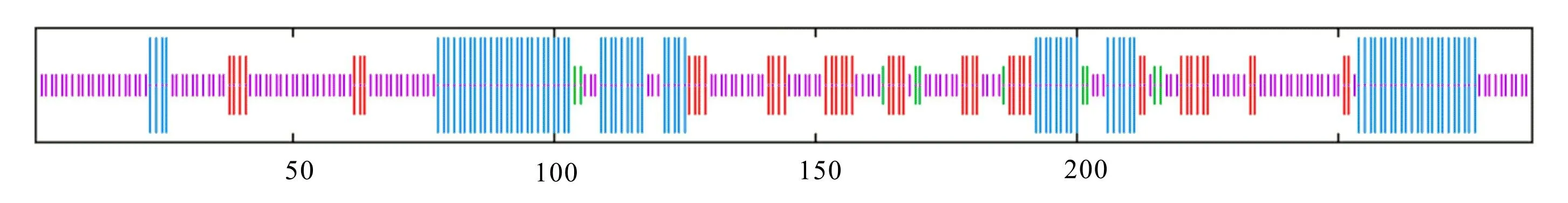
图 4 GA3ox-A蛋白二级结构预测Fig. 4 Secondary structure prediction of GA3ox-A protein
2.4 香蕉矮化突变体与野生型GA3ox氨基酸序列同源性比对分析
利用NCBI中的Blastp分析矮化突变体与野生型香蕉GA3ox蛋白的氨基酸序列,同时与其他物种的氨基酸序列同源性比对结果发现,矮化香蕉GA3ox的氨基酸序列与油棕(XP_010915137.1)、海枣(XP_008811603.3)、椰子(ARI45601.1)的同源性最高,序列同源性分别为62.5%、62.5%和61.5%。经序列比对发现,香蕉矮化突变体及其野生型亲本GA3ox的氨基酸序列在N末端比油棕、海枣和椰子3种植物的GA3ox少了63个氨基酸,C末端少了7个氨基酸(图5)。

黑色阴影和其他阴影框分别表示相同和相似氨基酸。Black shaded and other shaded boxes show identical and similar amino acids.图 5 矮化香蕉和野生型香蕉GA3ox与其他植物同源蛋白的序列比对分析Fig. 5 Sequence comparison and analysis of dwarf banana and wild type banana GA3ox with other plants homologous proteins
2.5 香蕉矮化突变体与野生型GA3ox基因的表达模式分析
利用qRT-PCR技术研究GA3ox基因在矮化突变体及野生型香蕉茎秆生长的不同叶龄期的表达情况。由图6可知,GA3ox基因在矮化突变体及野生型香蕉不同叶龄期的叶片中表达水平不同。在第10叶和第15叶龄期GA3ox在野生型香蕉叶片中的相对表达量均高于矮化香蕉,但在第20叶和第25叶叶龄期时野生型香蕉叶片中的相对表达量均显著低于矮化香蕉。在野生型香蕉叶片中,GA3ox在第15叶叶龄期表达水平最高,其次是第10叶叶龄期,而在第20叶和第25叶叶龄期时表达水平较低,第10叶和第15叶叶龄期的表达水平极显著高于第20叶和第25叶叶龄期,而在第20叶和第25叶叶龄期中的表达差异不显著; 在矮化突变体中,GA3ox在第15叶和第25叶叶龄期时表达水平显著或极显著高于第10叶和第20叶叶龄期时的表达水平,而在第10叶和第20叶叶龄期之间的表达差异不显著。
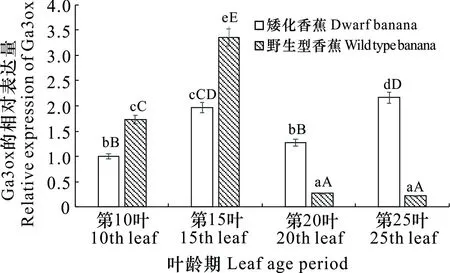
小写和大写字母分别表示0.01和0.05水平的显著性差异。下同。Lowercase and uppercase letters indicate significant differences at the 0.01 and 0.05 levels. The same below.图 6 GA3ox在矮化香蕉和野生型香蕉叶片中的表达水平Fig. 6 Expression levels of GA3ox in dwarf banana and wild type banana leaves
在香蕉假茎中,GA3ox基因在矮化突变体及野生型香蕉不同叶龄期的表达水平不同。在野生型香蕉茎秆中,GA3ox在第20叶叶龄期时表达水平最高,第25叶叶龄期是其次,在第10叶和第15叶叶龄期中的相对表达量次之,并且在第20叶叶龄期的表达水平显著或极显著高于其他叶龄期;在矮化突变体中,GA3ox在第25叶叶龄期表达水平最高,第20叶叶龄期是其次,在第20叶和第25叶叶龄期时表达水平显著或极显著高于第10叶和第15叶叶龄期时的表达水平,而在第10叶和第15叶叶龄期之间的表达差异不显著。同时,GA3ox在矮化植株茎秆中的表达量均显著或极显著低于野生型,其中在第10叶叶龄期时野生型茎秆中GA3ox的表达量是矮化型的32倍,在第15叶叶龄期野生型茎秆中GA3ox的表达水平是矮化型的7倍,在第20叶叶龄期时野生型植株茎杆中GA3ox的表达水平达最高,其表达水平是矮化型的2.2倍(图7)。
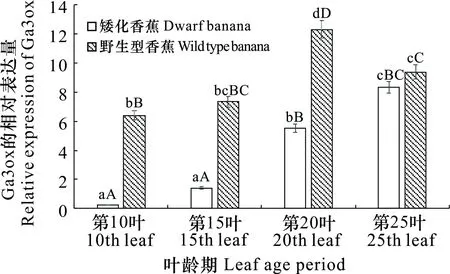
图 7 GA3ox在矮化香蕉和野生型香蕉假茎中的表达水平Fig. 7 Expression levels of GA3ox in dwarf banana and wild type banana pseudo stems
3 讨论与结论
赤霉素是影响植物生长发育的重要植物激素之一,植物体内活性赤霉素含量的减少会导致植物矮化,GA3-氧化酶(GA3ox)是赤霉素生物合成途径中的关键酶,GA3ox的重要功能是将无生物活性的GA9和GA20催化形成具有生物活性的GA1和GA4(Yamaguchi,2008)。目前,已在拟南芥(Helliwell et al., 1998)、水稻(Sasaki et al., 2002)、豌豆(Reinecke et al., 2013)等植株中鉴定得到GA3-氧化酶对植物茎秆的矮化具有重要的调控作用。为进一步探究GA3ox对香蕉矮化的分子调控机制,本研究克隆得到矮化香蕉及野生型香蕉的GA3ox的全长cDNA序列并对其氨基酸序列进行生物信息学分析。结果表明,GA3ox-A和GA3ox-G蛋白均属于不稳定蛋白,具有亲水性,并且GA3ox-A和GA3ox-G蛋白的二级结构均具有4种构象,蛋白磷酸位点分析显示GA3ox-A和GA3ox-G蛋白均含有丝氨酸、苏氨酸和酪氨酸 3 种氨基酸磷酸化位点,本研究结果与水稻(殷小林等,2019)、甘蔗(闫海锋等,2020)的GA3ox蛋白分析结果一致。氨基酸同源比对分析发现,矮化香蕉及其野生型GA3ox的氨基酸序列与油棕、海枣、椰子的同源性最高。
在水稻(李金华等,2007)、马铃薯(Roumeliotis et al., 2013)、紫花苜蓿(Dalmadi et al., 2008)和西瓜(Sun et al., 2020)等作物中研究发现,当GA3ox基因发生突变时会导致其编码产物的功能缺失,植物体内GA1和GA4含量降低,导致植株表现出矮化表型。在水稻矮化突变体d18中由于OsGA3ox2基因第2个外显子中的一个鸟嘌呤(G)的缺失改变了阅读框,导致OsGA3ox2功能的缺失,使得植株表现出矮化特性(李金华等,2007);在马铃薯中由于StGA3ox2基因的突变,造成植株节间变短(Roumeliotis et al., 2013),从而产生矮化表型;紫花苜蓿矮化突变体由于MsGA3ox氨基酸序列发生突变,损害MsGA3ox的功能并导致植株矮化(Dalmadi et al., 2008);在西瓜中由于GA3ox功能的缺失阻断了GA4的合成,导致植物赤霉素含量降低,植株表现出矮化性状(Sun et al., 2020)。本研究对威廉斯B6矮化突变体及其野生型亲本GA3ox的cDNA序列的比对分析发现,矮化突变体的GA3ox和野生型GA3ox的cds长度相同,但其编码产物的氨基酸序列中存在5个位点的差异。因此,推测GA3ox结构差异可能是引起香蕉茎秆矮化变异重要的因素。
GA3ox基因的表达异常对植物株高发育有较大的影响,目前已在马铃薯(Roumeliotis et al., 2013)、山核桃(魏广利等,2021)和豌豆(Reinecke et al., 2013)等作物中发现,GA3ox表达量的变化影响了植物株高性状。在马铃薯中,由于StGA3ox2的表达下调,因此突变体植株表现出矮小、节间较短等表型(Roumeliotis et al., 2013);在山核桃中,过量表达CcGA3ox基因使得植株变高(魏广利等,2021);豌豆PsGA3ox1的过量表达导致GA1含量增加,从而促进了豌豆节间的伸长(Reinecke et al., 2013)。本研究发现,GA3ox在香蕉矮化突变体茎秆中的表达水平显著低于野生型,这与马铃薯、豌豆、山核桃等植物的表达模式相似,说明GA3ox基因的表达量会影响植物株高,当GA3ox表达量下调时植株表现出节间缩短和矮化等表型。
综上所述,推测矮化香蕉表型的变异原因可能是GA3ox的序列发生突变导致其编码产物GA3ox的功能发生改变;或是其表达水平降低导致GA3ox的酶活性功能降低,使香蕉的内源GA1/GA4含量下降,从而影响了茎秆和其他器官的伸长生长。这表明GA3ox基因的突变和表达水平的变化对于香蕉茎秆的矮化变异可能具有重要的调控作用。但是,GA3ox的突变和表达水平降低是否引起香蕉茎秆矮化变异,还需要进一步验证GA3ox基因的功能来确定。
猜你喜欢
浙江农业科学(2022年1期)2022-12-28
亚热带农业研究(2022年1期)2022-08-08
农业工程技术(2021年12期)2021-12-23
江西农业学报(2021年2期)2021-12-08
河北果树(2021年4期)2021-12-02
农业科技通讯(2021年1期)2021-03-06
中国农业科技导报(2020年3期)2020-03-15
山西农业科学(2020年2期)2020-02-29
河北果树(2020年1期)2020-02-09
河北果树(2020年1期)2020-02-09
