
STAT3抑制剂及烟酰胺联合用药对肝癌HepG2细胞增殖的抑制作用及机制研究
2022-07-26 02:45詹雯静梁铭杰刘媛黄远凤王玮璇
天津医药 2022年7期
詹雯静,梁铭杰,刘媛,黄远凤,王玮璇
肝癌是我国最严重的恶性肿瘤之一,其发病率和病死率均位居前列[1]。作为机体内调控细胞增殖、凋亡、上皮-间充质转化(epithelial-mesenchymal transition,EMT)和糖代谢等多种生物过程的重要转录因子,信号传导和转录激活因子3(signal transducer and activator of transcription 3,STAT3)在包括肝癌在内的多种恶性肿瘤中被过度激活[2]。因此,对STAT3 的活性进行抑制是一种有效的潜在抗癌策略。Stattic和S3I-201是2种常用的STAT3抑制剂,通过干扰STAT3 的二聚化,使其不能进入细胞核,从而抑制STAT3的转录调控活性[3]。然而,在实际应用中需要高浓度的药物才能较好地抑制STAT3活性,但这会对正常细胞产生损伤。目前尚无获准应用于临床的直接靶向STAT3 的药物[4]。烟酰胺(nicotinamide,Nam)是烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NAD)的前体。作为细胞生命活动的重要分子,NAD 参与了包括糖酵解、三羧酸循环和β-氧化等在内的重要生物过程[5]。补充NAD 前体Nam 具有包括防治癌症在内的多种有益作用[6]且可以下调肺癌细胞中STAT3 Y705 位的磷酸化水平[7]。但Nam 及其与STAT3 抑制剂联合用药对肝癌的影响及机制仍不明确。本研究以肝癌细胞HepG2为模型,从细胞凋亡、EMT及糖酵解角度探讨STAT3 抑制剂、Nam 以及两者联合用药对肝癌细胞增殖的影响及其机制,以期为肝癌临床治疗提供新方案。
1 材料与方法
1.1 材料 人肝癌细胞株HepG2购自中国科学院上海生科院细胞资源中心。STAT3 抑制剂S3I-201 和Stattic 购自Selleck 公司;Nam购自Sigma-Aldrich 公司;青/链霉素购自维森特生物技术(南京)有限公司;胎牛血清、DMEM 高糖培养基、磷酸盐缓冲液(PBS)购自Thermo Fisher Scientific 公司;PVDF 膜购自Millipore 公司;辣根过氧化物酶(HRP)标记的羊抗兔和羊抗鼠IgG以及p-STAT3兔单克隆抗体、β-actin鼠单克隆抗体购自Cell Signaling Technology 公司;STAT3 鼠单克隆抗体购自ProteinTech 公司;ECL 发光液购自Bio-Rad 公司;RNAiso-Plus 试剂、逆转录试剂盒和荧光定量PCR(qPCR)试剂盒购自Takara公司;BCA蛋白定量试剂盒、RIPA细胞裂解液、二甲基亚砜(DMSO)购自北京索莱宝科技有限公司;蛋白酶抑制剂cocktail 和磷酸酶抑制剂购自Selleck 公司。Centrifuge 5810R 离心机购自Eppendorf 公司;NanoDrop 2000 核酸浓度测定仪购自Thermo Fisher Scientific 公司;Eon微孔板分光光度计购自BioTek 公司;LightCycler 480 荧光定量PCR 仪购自Roche 公司;电泳仪、转膜仪及凝胶成像系统购自Bio-Rad公司。
1.2 Western blot检测STAT3和p-STAT3蛋白的表达 HepG2用含有1%青霉素/链霉素和10%胎牛血清的高糖DMEM 培养基,于37 ℃、含5% CO2的培养箱中培养。将对数生长期的HepG2细胞接种于6 孔板中,过夜培养,待细胞贴壁后,根据文献中使用的药物浓度,设置对照组(Ctrl)、单用药组(100µmol/L S3I-201[8]、1.5µmol/L Stattic[9]和5 mmol/L Nam[7])与联合用药组(100 µmol/L S3I-201+5 mmol/L Nam 组和1.5 µmol/L Stattic+5 mmol/L Nam组)。药物处理24 h后,吸去细胞培养基,用预冷的PBS洗涤细胞2次,收集细胞后加入适量的RIPA裂解液(添加1%蛋白酶抑制剂cocktail和1%磷酸酶抑制剂),冰上静置30 min裂解细胞。在4 ℃,12 000 r/min离心20 min 后收集上清液。用BCA 蛋白检测试剂盒测定浓度后,用12%SDS-PAGE 凝胶分离蛋白,然后将蛋白转移到PVDF 膜上。使用5%的脱脂牛奶进行封闭,一抗STAT3(稀释比例1∶1 000)、p-STAT3(稀释比例1∶1 000)和β-actin(稀释比例1∶3 000)4 ℃孵育过夜,HRP 标记的羊抗兔或羊抗鼠IgG二抗(稀释比例1∶2 000)室温孵育1.5 h后,将ECL化学发光显影液均匀地滴在PVDF 膜上,在全自动凝胶成像仪中进行显影,结果使用Image Lab软件进行分析。
1.3 RNA的提取和qPCR 将对数生长期的HepG2细胞接种于6孔板中,过夜培养,待细胞贴壁后,按照1.2所述设置对照组(Ctrl)、单用药组与联合用药组,处理24 h后收集细胞提取RNA并进行逆转录。RNA的提取以及qPCR过程根据Takara公司的说明书进行。具体反应体系如下:TB Green Premix Ex TaqⅡ(2×)5 µL,cDNA(1 g/L)2 µL,上、下游引物(10µmol/L)各0.6µL,ddH2O 1.8µL。反应条件如下:95 ℃30 s;95 ℃5 s,60 ℃30 s,40 个循环。采用2-ΔΔCt法对基因相对表达水平进行定量分析。引物由北京睿博兴科生物技术公司合成,见表1。
1.4 细胞增殖实验 将对数生长期的HepG2细胞接种于6孔培养板中,过夜培养,待细胞贴壁后,按照1.2 所述设置对照组(Ctrl)、单用药组与联合用药组,处理36 h 后,细胞长至80%~90%时,收集细胞,用0.4%的台盼蓝试剂染色,用细胞计数板进行细胞计数。
1.5 统计学方法 采用GraphPad Prism 8软件进行数据分析与做图,数据采用均数±标准差(±s)表示。多组间比较采用单因素方差分析(One-way ANOVA),组间多重比较采用Tukey法,P<0.05为差异有统计学意义。
2 结果
2.1 STAT3 抑制剂及Nam 对STAT3 Y705 磷酸化水平的影响 S3I-201、Stattic 及Nam 均可显著降低HepG2细胞STAT3 Y705 的磷酸化水平,且联合用药(Nam+S3I-201 和Nam+Stattic)比单用药对STAT3 Y705磷酸化抑制效果更强,见图1。
2.2 STAT3 抑制剂及Nam 对STAT3 下游靶基因的影响 相比Ctrl组,S3I-201、Stattic及Nam单用药与联合用药组STAT3 下游靶基因SNAIL1、VEGFA和ZEB1的mRNA 表达水平显著降低;与S3I-201 单药相 比,Nam 和S3I-201 联 合 用 药 组 中SNAIL1和VEGFAmRNA 表达水平显著降低;与Stattic 单药相比,Nam 和Stattic 联 合 用 药 组 中VEGFA和ZEB1mRNA表达水平显著降低(P<0.05),见图2。

Tab.1 Primer sequences for qPCR表1 qPCR引物序列

Fig.2 Effects of STAT3 inhibitor and Nam on the mRNA expression levels of STAT3 downstream target genes图2 STAT3抑制剂及Nam对STAT3下游靶基因mRNA表达水平的影响
2.3 STAT3 抑制剂及Nam 对细胞增殖的影响 相比于Ctrl 组,Nam 单独用药与联合用药可以显著抑制细胞增殖,并且Nam 与Stattic 联合用药对于细胞增殖的抑制效果比Stattic单独用药更明显,见图3。
2.4 STAT3 抑制剂及Nam 对细胞增殖相关基因表达的影响 相比Ctrl 组,单用药组和联合用药组MCM7和MYCmRNA 表达水平显著降低,且联合用药组中MCM7的表达水平均低于单用药组;相比Ctrl组,除S3I-201 及Stattic 处理组,其余各组MKI67mRNA表达水平显著降低。见图4。
2.5 STAT3 抑制剂及Nam 对细胞凋亡相关基因表达的影响 相比Ctrl 组,单用药组和联合用药组中促凋亡基因BAXmRNA 表达量显著升高,抗凋亡基因BCL-2和MCL-1mRNA 表达量显著降低。且Nam 和Stattic 联合用药组中BAXmRNA 表达水平显著高于单用药组,Nam 和S3I-201 联合用药组中BCL-2mRNA表达水平显著低于单用药组,见图5。
2.6 STAT3 抑制剂及Nam 对EMT 相关基因表达的影响 相比Ctrl 组,除Stattic 外的用药组中TJP1mRNA 表达水平显著升高,各用药组SMAD3mRNA表达水平显著降低。Nam 和Stattic 联合用药组中TJP1mRNA 表达水平显著高于单独用药组,SMAD3mRNA表达水平显著低于单独用药组,见图6。
2.7 STAT3 抑制剂及Nam 对糖代谢相关基因的影响 相比Ctrl 组,单用药组和联合用药组中糖酵解相关基因HK2、PFKL和PKMmRNA表达水平显著降低;与S3I-201 单独用药组相比,Nam 和S3I-201 联合用药组中HK2、PFKL和PKMmRNA表达水平显著降低;与单独用药组相比,Nam和Stattic 联合用药组中PKMmRNA 表达水平显著降低。见图7。相比Ctrl 组,单用药组和联合用药组中葡萄糖转运蛋白GLUT1mRNA表达水平显著降低,除S3I-201外的用药组中乳酸脱氢酶LDHAmRNA 表达水平显著降低。且Nam 和S3I-201 联合用药组中LDHAmRNA表达水平显著低于S3I-201单用药组,Nam 和Stattic联合用药组中LDHAmRNA 表达水平显著低于Stattic单用药组。见图8。

Fig.3 Effects of STAT3 inhibitor and Nam on cell proliferation图3 STAT3抑制剂及Nam对细胞增殖的影响
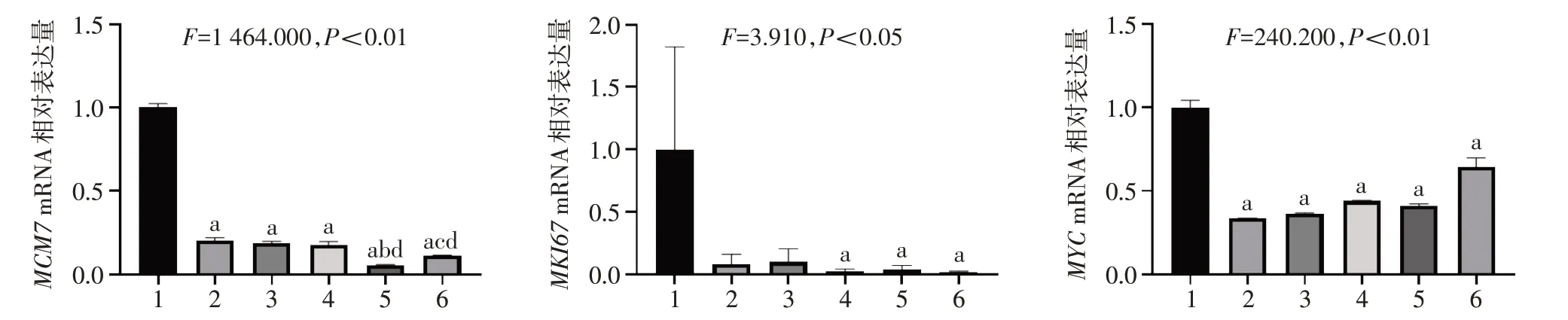
Fig.4 Effects of STAT3 inhibitor and Nam on the mRNA expression levels of cell proliferation-related genes图4 STAT3抑制剂及Nam对细胞增殖相关基因mRNA表达水平的影响
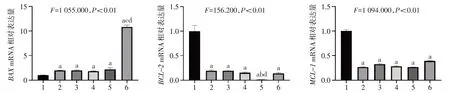
Fig.5 Effects of STAT3 inhibitor and Nam on the mRNA expression levels of apoptosis-related genes图5 STAT3抑制剂及Nam对细胞凋亡相关基因mRNA表达水平的影响
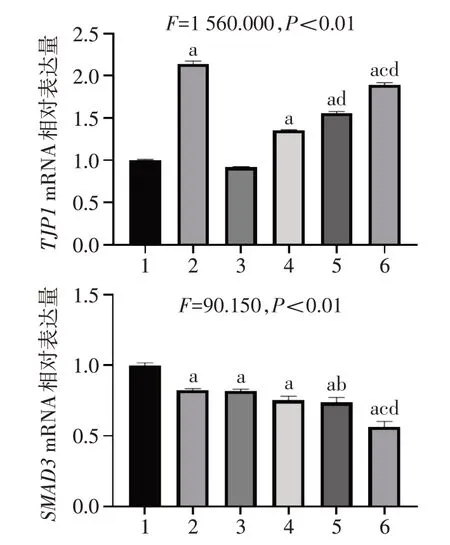
Fig.6 Effects of STAT3 inhibitor and Nam on the mRNA expression levels of EMT-related genes图6 STAT3抑制剂及Nam对EMT相关基因mRNA表达水平的影响
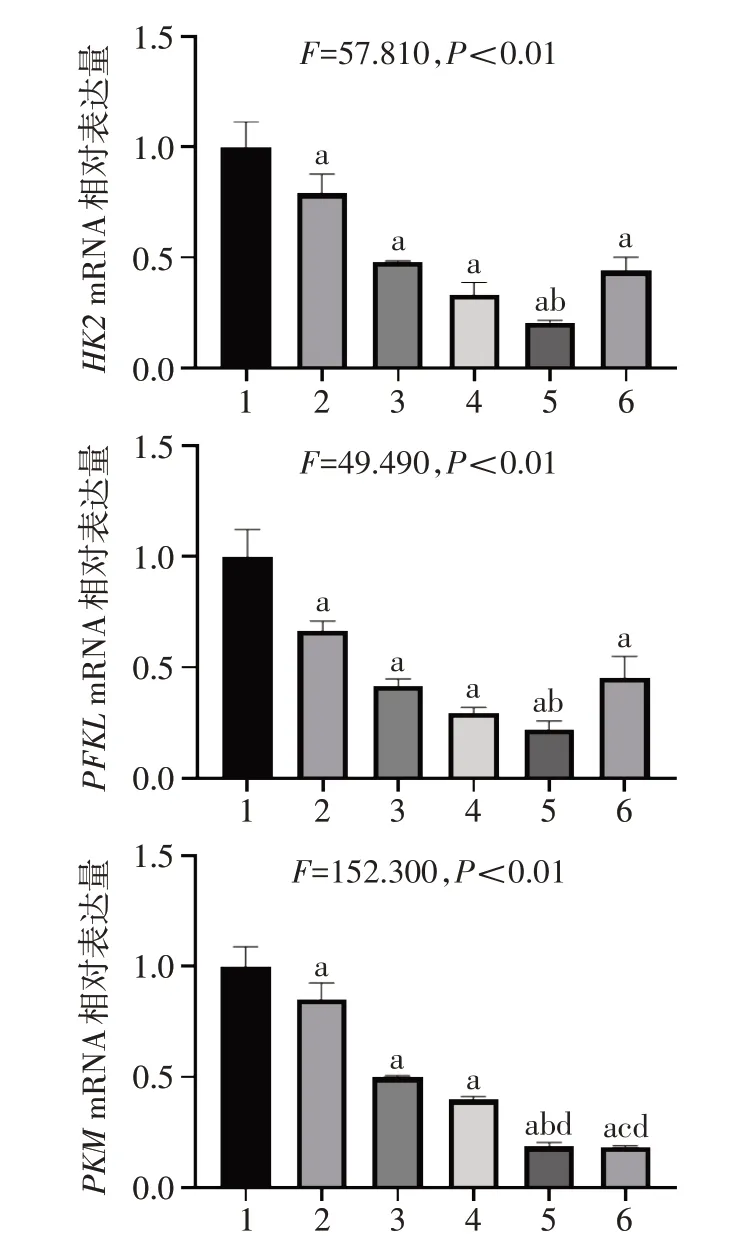
Fig.7 Effects of STAT3 inhibitor and Nam on the mRNA expression levels of glycolysis-related genes图7 STAT3抑制剂及Nam对糖酵解相关基因mRNA表达水平的影响
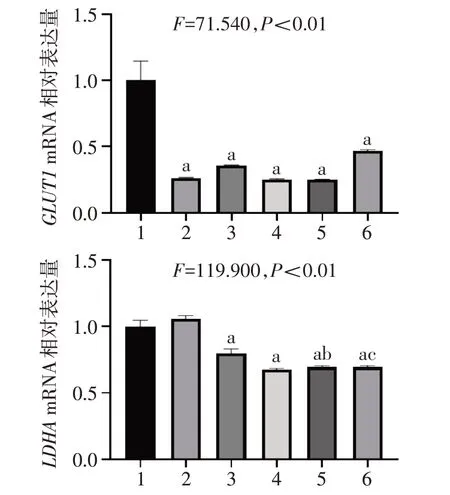
Fig.8 Effects of STAT3 inhibitor and Nam on the mRNA expression levels of GLUT1 and LDHA图8 STAT3抑制剂及Nam对GLUT1和LDHA mRNA表达水平的影响
3 讨论
相关研究表明,Stattic 和S3I-201 可以降低肺癌[7]、前列腺癌[10]和鼻咽癌[11]细胞中STAT3 Y705位的磷酸化水平,同时Nam可以降低肺癌细胞中Y705位的磷酸化水平[7]。本研究发现,单独使用STAT3抑制剂或Nam均可以降低肝细胞癌HepG2中STAT3 Y705 位的磷酸化水平,抑制细胞增殖,且联合用药效果更显著。
肝癌的发生与发展受多种因素的影响,细胞增殖和凋亡的失控是其中重要的环节。癌细胞的一个重要标志是具有无限的增殖能力。MCM7、MKI67和MYC是3 个与细胞增殖密切相关的基因,MCM7 是ATP依赖的DNA解旋酶,在DNA复制起始发挥重要作用,是细胞增殖活性的标志物[12]。MCM7 过表达可以促进肝细胞癌的增殖,其高表达与肝细胞癌不良的预后相关[13]。MKI67 是一个核蛋白,其在肝细胞癌中表达量升高[14-15],敲低MKI67 可以抑制肝细胞癌的增殖[16]。MYC 在包括肝细胞癌在内的多种类型肿瘤中表达量上调[17],在肝癌发生和肝细胞增殖中起着重要作用[18]。本研究发现STAT3抑制剂或者Nam 处理可以降低MCM7、MKI67和MYCmRNA表达水平,且联合用药效果更显著。细胞凋亡是有机体为了维持自身内环境稳定而启动的细胞程序性死亡的过程,这一过程受到BCL-2蛋白家族的调控,包括抗凋亡蛋白(如BCL-2、BCL-XL和MCL-1)以及促凋亡蛋白(如BAX和BAK)[19]。研究表明,肝癌组织中BCL-2 和MCL-1 高表达,BAX 低表达,肿瘤细胞凋亡受到抑制[20-21]。本研究发现,STAT3 抑制剂或者Nam 处理后,促凋亡基因BAXmRNA 表达量升高,抗凋亡基因BCL-2和MCL-1mRNA 表达量降低,且联合用药效果更显著,提示STAT3抑制剂以及Nam可以通过促进细胞凋亡来抑制肝癌细胞增殖。
EMT是指上皮细胞向具有迁移和侵袭能力的间充质细胞转化的过程,与肿瘤细胞增殖密切相关[22]。本研究发现,STAT3抑制剂或者Nam处理后,与EMT相关基因SNAIL1、VEGFA、ZEB1和SMAD3mRNA 表达量降低,TJP1mRNA 表达量升高,且联合用药效果更显著。这些基因不仅在EMT 过程中发挥重要作用,还影响肿瘤细胞的增殖。TJP1是将跨膜的紧密连接蛋白和细胞质蛋白、肌动蛋白细胞骨架相连的支架蛋白[23],肝癌组织中TJP1 表达下调,而过表达TJP1 可以降低肝癌细胞增殖,抑制细胞迁移[24]。SMAD3是EMT过程的关键调控因子,可以被转化生长因子(TGF)-β 激活,从而启动下游一系列与促进肿瘤细胞生长和侵袭相关基因的转录[25]。Snail1是EMT和细胞增殖的重要转录因子,可通过下调E-钙黏蛋白,上调间充质细胞相关基因的表达来促进EMT 过程[26]。研究表明,过表达Snail1 可以通过上调细胞周期蛋白(Cyclin)D1 和增殖细胞核抗原(PCNA)的表达水平来促进肿瘤细胞增殖,反之亦然[27]。VEGFA是一种促进血管生成的生长因子,可以促进细胞迁移。研究表明,VEGFA在肝癌组织与肝癌细胞系中高表达[28]。在结直肠癌细胞中,VEGFA过表达可以促进细胞增殖[29]。在小鼠体内,VEGFA 过表达可促进肝癌生长,而抑制VEGFA 活性可以抑制肝癌生长[30]。ZEB1 是EMT 过程的关键转录因子,可以促进EMT 过程。研究表明,ZEB1 在肝癌细胞中高表达[31],敲低ZEB1可以抑制肝癌细胞增殖[32]。因此,笔者认为STAT3抑制剂及Nam 可能通过调控EMT 相关基因的表达,抑制肝癌细胞增殖。
正常细胞主要依赖线粒体的氧化磷酸化来产生细胞所需的能量,而大部分的肿瘤细胞依赖于有氧糖酵解,即无论是否存在氧气,肿瘤细胞都倾向于将大部分的葡萄糖转化为乳酸,这一现象被称为Warburg 效应[33]。Warburg 效应不仅可以为快速增殖的肿瘤细胞提供合成核酸、脂质以及蛋白的原料,还可以调节活性氧水平和肿瘤微环境,从而赋予肿瘤细胞生长上的优势、耐药性和转移性[34]。本研究发现,STAT3 抑制剂及Nam 可以抑制糖酵解过程关键基因的表达水平,并抑制葡萄糖转运蛋白GLUT1以及乳酸脱氢酶LDHA的表达,且联合用药效果更显著。这提示STAT3 抑制剂及Nam 处理会导致糖酵解过程被抑制,减少乳酸生成,抑制Warburg 效应,从而影响肝癌细胞的能量代谢,抑制肝癌细胞增殖。
综上,STAT3抑制剂及NAD 前体Nam 可以抑制肝癌细胞增殖,并且联合用药效果更显著,进一步的机制研究表明,其对肝癌细胞增殖的抑制作用可能是通过对细胞增殖、凋亡、EMT及糖代谢相关基因表达水平的调控实现的,提示STAT3 抑制剂和Nam 联合用药可作为潜在的肝癌治疗方案。
猜你喜欢
波谱学杂志(2022年1期)2022-03-15
昆明医科大学学报(2022年1期)2022-02-28
华夏医学(2021年1期)2021-12-05
天津医科大学学报(2021年4期)2021-08-21
现代临床医学(2021年2期)2021-03-29
读与写·上旬刊(2018年1期)2018-05-06
分析化学(2017年12期)2017-12-25
教育教学论坛(2016年51期)2017-03-22
教育教学论坛(2016年49期)2017-02-27
安徽医科大学学报(2015年9期)2015-12-16
