
妊娠对雌性大鼠冷防御性肩胛间区棕色脂肪组织产热的影响及其机制
2022-06-18 06:19侯晓钰谢江燕宋宜安胥建辉
吉林大学学报(医学版) 2022年3期
侯晓钰,李 娅,谢江燕,宋宜安,张 洁,胥建辉
(1.成都医学院第一附属医院产科,四川 成都 610500;2.成都医学院 体温与炎症四川省高校重点实验室,四川 成都 610500)
冷防御反应是恒温动物暴露于冷环境中时启动的前馈体温调节活动,其可在环境冷刺激引起体核温度变化前促使机体产热增加,散热减少,以维持体核温度相对稳定[1-3]。因此冷防御反应是恒温动物面临环境温度挑战时的关键体温调节活动,具有重要的生理意义[1-3]。妊娠期母体的多个系统和器官会发生一系列生理变化,其中也包括体温调节系统[4-5]。已有研究[6-7]表明:多种哺乳动物的基础体温和热中性温度区在妊娠晚期时明显下移;此外,多种刺激源(如应激和感染等)引起的体温调节活动在大鼠妊娠晚期时也受到了明显的抑制[8]。然而,在大鼠妊娠期冷防御反应是否具有适应性的改变尚不清楚。
肩胛间区棕色脂肪组织(interscapular brown adipose tissue,iBAT)是参与冷防御反应的重要产热效应器[9-10]。机体皮肤受到冷刺激时,iBAT的产热情况(即冷防御性iBAT产热)直接反映了冷防御响应的 强 度[1,9]。研究[11]显示:长时间 处 于冷环境中的妊娠晚期大鼠体温下降幅度明显高于非妊娠期雌性大鼠,提示妊娠晚期大鼠对冷的耐受能力下降。本文作者推测妊娠期大鼠的冷防御反应可能受到抑制,但目前尚无相关报道。本研究采用腹部皮肤冷刺激的方法建立冷防御反应模型,通过监测非妊娠期、妊娠中期和妊娠晚期雌性大鼠皮肤受到冷刺激时iBAT的产热情况及支配iBAT的交感神经放电活动(sympathetic nerve activity,SNA),观察妊娠对雌性大鼠冷防御反应的影响,并探讨其作用机制。
1 材料与方法
1.1 动物、主要试剂和仪器42只成年SD雌性大鼠和28只雄性大鼠,体质量220~250 g,购自成都达硕实验动物有限公司,动物生产许可证号:SCXK(川)2020-030。氨基甲酸乙酯、α-氯醛糖、液体石蜡和体积分数为75%酒精(四川省成都市科龙化工试剂厂)。多通道温度信号测量系统(JK808)和K型热电偶(常州金艾联电子科技有限公司),生物信号采集系统(BF420S)和鼠恒温试验台(JR30)(成都泰盟软件有限公司),红外线加热器(CQ-61P)(重庆航天火箭电子技术有限公司),冷光源(QAXK-TY)(千奥星科南京生物科技有限公司)。
1.2 动物实验分组和处理所有大鼠均自由饮食和进水,饲养温度(25±1)℃,明暗时间各12 h(昼光期为06:00~18:00)。适应性饲养1周后,将雌性大鼠随机分为非妊娠期、妊娠中期和妊娠晚期组,每组各14只。非妊娠期组雌性大鼠不与雄性大鼠合笼。采用阴道涂片法检测妊娠中期和妊娠晚期组雌性大鼠动情周期,在动情期前一天将雌鼠和雄鼠合笼交配,次日上午检查,以发现阴道栓视为交配成功,交配当天为妊娠第0天,此后分别单独饲养交配成功雌性大鼠至妊娠第15和20天。
1.3 各组大鼠皮肤、直肠和iBAT温度检测实验当日上午9时,腹腔注射17%氨基甲酸乙酯和1%α-氯醛糖混合液(5 mL·kg-1)麻醉各组大鼠,室温设置为25℃,采用恒温热板和红外加热灯将大鼠体核温度维持在约37℃,剔除大鼠腹部鼠毛,将水袋置于腹下,以备冰水注入。此后,剔除大鼠背部肩胛间区鼠毛,75%酒精消毒后沿肩胛间区正中线切开1个1.5~2.0 cm切口,暴露iBAT(图1A),将多通道温度信号采集系统的温度探头插入右侧iBAT中(iBAT沿大鼠背部肩胛间区中线对称分布,分为左右两叶),采用缝合线将温度探头固定于背部肌肉上。其余温度探头直接插入直肠中或采用纸胶布固定于大鼠腹部,以同步测量iBAT、直肠和腹部皮肤温度。
将记录的温度信号导入JK80X数据分析软件,通过该软件分析各组大鼠皮肤、iBAT和直肠温度。计算各组大鼠皮肤冷刺激前30 s温度的平均值,作为基础值;计算各组大鼠皮肤冷刺激时持续30 s的最高(或最低)温度平均值,作为冷刺激时的响应值。接受冷刺激后,各组大鼠iBAT、直肠和皮肤温度的变化幅度值(上升幅度或降低幅度)=接受冷刺激时温度平均值-接受冷刺激前温度平均值。
1.4 各组大鼠皮肤、直肠温度和iBAT SNA情况检测实验当日上午9时,腹腔注射17%氨基甲酸乙酯和1%α-氯醛糖混合液(5 mL·kg-1)麻醉大鼠,室温设置为25℃,采用恒温热板和红外加热灯将大鼠体核温度维持在约37℃,剔除大鼠腹部鼠毛,将水袋置于腹部下,以备冰水注入。此后,剪开气管行气管插管术,75%酒精消毒后沿肩胛间区正中线切开1个1.5~2.0 cm切口。止血钳钝性分离iBAT,暴露位于肩胛骨下的交感神经节后纤维,玻璃分针分离支配右侧iBAT的交感神经(图1B),剪断神经外周端,挂于引导电极上,滴加液体石蜡保持神经湿润,将引导电极与生物信号采集系统相连,记录iBAT SNA(单位为μV)。同时采用多通道温度信号采集系统同步监测各组大鼠直肠和腹部皮肤温度。
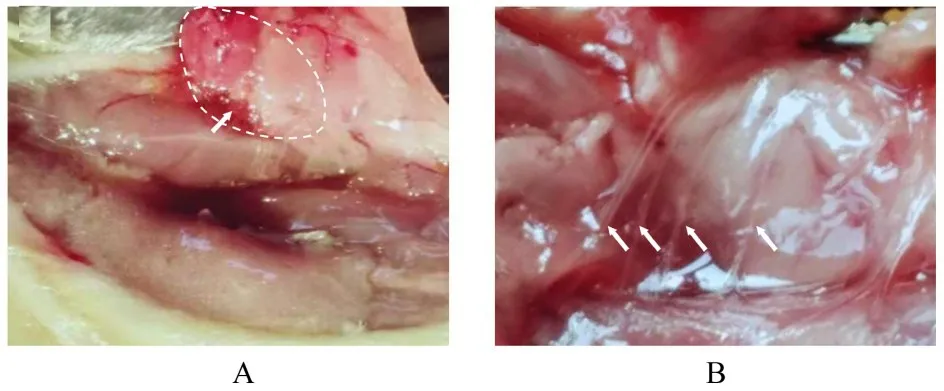
图1 雌性大鼠的iBAT和支配iBAT的交感神经纤维形态表现Fig.1 Morphology of iBAT and sympathetic nerve innervating iBAT of female rats
将记录的交感神经放电信号导入Spike2数据分析软件,通过该软件分析交感神经放电幅度值。计算皮肤冷刺激前30 s的放电幅度平均值,作为交感神经放电基础值;计算皮肤冷刺激时持续30 s的最大放电幅度平均值,作为冷刺激时交感神经放电响应值。接受冷刺激后,iBAT SNA变化幅度值(上升幅度或降低幅度)=接受冷刺激后iBAT SNA平均值-接受冷刺激前iBAT SNA平均值。
1.5 大鼠腹部皮肤冷刺激方案大鼠直肠温度稳定后,向水袋中注入4℃冰水(冰箱中预冷),150 s后抽出冰水,降低大鼠腹部皮肤温度约10℃,同时采用多通道体温和生物信号采集系统监测大鼠体温和交感神经放电变化情况。
1.6 统计学分析采用SPSS 26.0统计软件进行统计学分析。各组大鼠皮肤温度、直肠温度、iBAT温度和iBAT SNA以±s表示,多组间样本均数比较采用单因素方差分析(One-way AVOVA),组间样本均数两两比较采用SNKq检验。以P<0.05为差异有统计学意义。
2 结 果
2.1 各组大鼠皮肤、直肠和iBAT温度与冷刺激前比较,接受冷刺激后,各组大鼠腹部皮肤温度均降低(P<0.05);与冷刺激前比较,接受冷刺激后,各组大鼠的直肠温度差异无统计学(P>0.05)。与冷刺激前比较,接受冷刺激后,非妊娠期和妊娠中期组雌性大鼠iBAT温度明显升高(P<0.05),妊娠晚期组大鼠iBAT温度差异无统计学意义(P>0.05)。此外,接受冷刺激后,各组大鼠皮肤温度的降低幅度组间比较差异无统计学意义(P>0.05);各组大鼠直肠温度的变化幅度组间比较差异无统计学意义(P>0.05)。接受冷刺激后,非妊娠期组大鼠和妊娠中期组大鼠iBAT温度的上升幅度组间比较差异无统计学意义(P>0.05)。接受冷刺激后,与非妊娠期组或妊娠中期组大鼠比较,妊娠晚期组大鼠iBAT温度的上升幅度明显减小(P<0.05)。见图2和表1。

表1 皮肤冷刺激前后各组大鼠皮肤、直肠和iBAT温度Tab.1 Temperature of skin and rectum,and iBAT of rats in various groups before and after skin cooling(n=7,x±s,θ/℃)

图2 各组大鼠皮肤受到冷刺激后皮肤、直肠和iBAT温度曲线Fig.2 Temperature curves of skin,rectum and iBAT of rats in various groups after skin cooling
2.2 各组大鼠皮肤和直肠温度及iBAT SNA与冷刺激前比较,接受冷刺激后,各组大鼠腹部皮肤温度均明显降低(P<0.05),直肠温度差异无统计学意义(P>0.05)。与冷刺激前比较,接受冷刺激后,非妊娠期组和妊娠中期组雌性大鼠iBAT SNA明显增加(P<0.05);妊娠晚期组大鼠的iBAT SNA差异无统计学意义(P>0.05)。此外,接受冷刺激后,各组大鼠皮肤温度降低幅度组间比较差异无统计学意义(P>0.05);各组大鼠直肠温度改变幅度组间比较差异无统计学意义(P>0.05)。接受冷刺激后,非妊娠期组和妊娠中期组雌性大鼠iBAT SNA上升幅度比较差异无统计学意义(P>0.05)。接受冷刺激后,与非妊娠期组或妊娠中期组比较,妊娠晚期组大鼠iBAT SNA上升幅度明显减小(P<0.05)。见表2和图3。
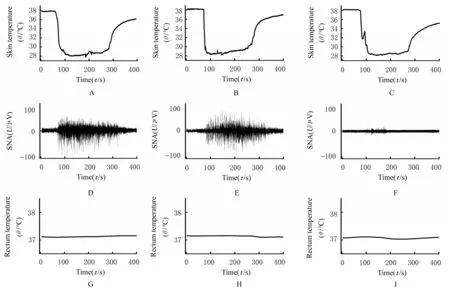
图3 各组大鼠皮肤受到冷刺激后皮肤和直肠温度及iBAT SNAFig.3 Skin and rectum temperature and iBAT SNA of rats in various groups after skin cooling
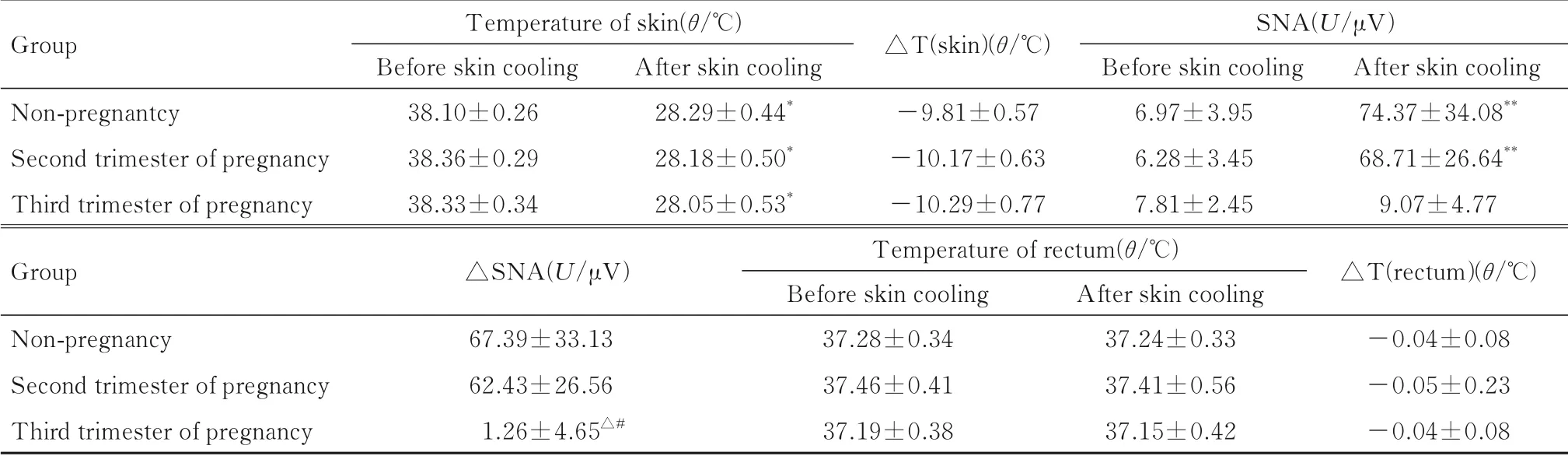
表2 皮肤冷刺激前后各组大鼠皮肤温度、iBAT SNA和直肠温度Tab.2 Temperature of skin,iBAT SNA,and temperature of rectum of rats in various groups before and after skin cooling(n=7,x±s)
3 讨 论
冷防御反应主要包括环境冷刺激引起的棕色脂肪和骨骼肌颤栗产热及血管收缩保热[1]。iBAT产热是冷防御反应的重要热量来源之一,学者们[1-12]常通过监测皮肤受到冷刺激时机体iBAT的温度变化来评估冷防御反应的情况。本研究采用冰水刺激大鼠腹部皮肤的同时监测腹部皮肤温度变化的结果显示:各组大鼠皮肤温度下降幅度比较差异无统计学意义,说明给予了各组大鼠相同程度的冷刺激。本研究结果显示:与接受冷刺激前比较,受到冷刺激时,非妊娠期和妊娠中期大鼠的直肠温度无明显改变,iBAT温度明显升高;且这2组大鼠iBAT的升温幅度相似。上述结果与国外学者[1]在雄性大鼠上的发现是相似的,说明皮肤冷刺激而非体核温度改变导致了非妊娠期和妊娠中期雌性大鼠的iBAT产热增加;提示妊娠中期和非妊娠期雌性大鼠与雄性大鼠一样具有正常的冷防御反应。本文作者发现:与接受冷刺激前比较,受到冷刺激时,妊娠晚期大鼠的直肠温度无明显改变,iBAT温度无明显变化;与非妊娠期和妊娠中期雌性大鼠比较,妊娠晚期大鼠受到冷刺激时的iBAT温度上升幅度明显减小,说明皮肤冷刺激无法引起妊娠晚期大鼠iBAT产热增加,同时还提示妊娠晚期大鼠的冷防御反应受到明显抑制。与本研究结果一致,国外学者[11]发现长 时 间(30~60 min)暴 露 于0℃或10℃冷环境中的妊娠晚期大鼠体核温度明显低于处于同样环境中的非妊娠期雌性大鼠。
近年冷防御反应的机制逐渐被揭露:皮肤温度感受器接收到的冷信号经脊髓逐级上传至体温调节中枢,引起产热神经元兴奋,参与产热的下行神经通路被激活,进而导致交感节前神经元兴奋,支配iBAT的交感神经放电增强,iBAT产热增加[3,12-14]。为了进一步探讨妊娠期大鼠冷防御反应改变是否与来自中枢的体温调节信号受到影响有关,本文作者对比观察了非妊娠期、妊娠中期和妊娠晚期大鼠的冷防御性交感神经放电活动。本研究结果显示:与接受冷刺激前比较,受到冷刺激时非妊娠期和妊娠中期雌性大鼠的直肠温度无明显改变,iBAT SNA增强,说明皮肤冷刺激而非体核温度改变导致了非妊娠期和妊娠中期雌性大鼠的iBAT SNA增强。此外,受到冷刺激时,非妊娠期和妊娠中期雌性大鼠iBAT SNA的增幅相似,且这2种大鼠iBAT SNA的变化和iBAT温度的变化一致;提示妊娠中期和非妊娠期雌性大鼠具有正常的冷防御反应,与2种大鼠冷防御性iBAT SNA未受到明显影响有关。本研究结果显示:与接受冷刺激前比较,接受冷刺激后妊娠晚期大鼠直肠温度无明显改变,iBAT SNA无明显变化;此外,与非妊娠期组和妊娠中期组比较,妊娠晚期组大鼠接受冷刺激后iBAT SNA上升幅度明显减小;且妊娠晚期组大鼠接受冷刺激后iBAT SNA的变化与iBAT温度的变化一致,提示妊娠晚期大鼠冷防御反应减弱与该期大鼠的冷防御性iBAT SNA受到明显抑制有关。
既往研究[15]表明:致热介质前列腺素E2引起的交感神经放电和棕色脂肪产热在大鼠妊娠晚期时同样明显减弱,提示来自中枢的促产热体温调节信号均受到抑制。国外学者[8]发现这种现象可能与妊娠晚期大鼠下丘脑一氧化氮合成酶的表达增加有关。因为一氧化氮可通过抑制体温调节中枢(下丘脑视前区)神经元的放电活动降低体温或抑制发热;且脑室内注射一氧化氮合成酶抑制剂可以恢复妊娠晚期大鼠的发热反应[16-17]。上述研究提示妊娠晚期大鼠冷防御反应减弱可能与一氧化氮释放引起促产热的神经通路抑制有关。该假设还有待进一步的实验进行验证。
iBAT产热除了接受体温调节中枢的调控,还受到其自身产热能力的影响[9]。近年已有研究[18-19]显示:与非妊娠期雌性大鼠比较,妊娠中期和晚期大鼠(或小鼠)iBAT内解耦连蛋白1均表现出相似程度的降低;此外,这两种大鼠iBAT的棕色脂肪细胞向白色或米色脂肪细胞转化,表明妊娠中晚期大鼠(或小鼠)iBAT的产热能力降低。据此推测:如果妊娠中晚期大鼠iBAT产热能力的改变在这些大鼠的冷防御反应中发挥了重要的作用,妊娠中期和晚期大鼠受到冷刺激时的iBAT产热均明显降低。但与此推论不一致的是,本研究结果显示:妊娠中期大鼠受到冷刺激时的iBAT产热并无明显降低。上述研究结果提示:妊娠中晚期大鼠的产热能力改变可能并非妊娠晚期大鼠受到冷刺激时产热能力减弱的主要原因,其在妊娠期大鼠冷防御反应的适应性改变中可能未发挥不可或缺的作用。
为了满足胎儿生长发育的需求,妊娠期母体会发生一系列适应性改变,以储存能量和减少消耗[19-20]。与此相反,正常的冷防御反应却会引起交感神经活动增强,棕色脂肪产热增加,机体的能量消耗上升[9]。研究[21-22]显示:妊娠期母体的棕色脂肪组织产热对胎盘营养物质转运、胎儿生长和子代的葡萄糖耐受性具有负面的影响。国外学者[9,23]发现:冷防御反应在处于极度的低温环境和食物缺乏等特殊情况下也会适应性的减弱。因此,妊娠晚期大鼠冷防御反应减弱可能是一种适应性改变,其有助于母体保存能量,维持胎儿正常生长发育。
综上所述,非妊娠期和妊娠中期大鼠具有正常的冷防御反应;妊娠晚期大鼠的冷防御反应减弱。这种现象与妊娠晚期时冷防御性交感神经放电活动受到抑制有关。本研究结果有助于更深入地认识妊娠期母体体温调节活动的适应性改变及其机制。
猜你喜欢
现代临床医学(2022年1期)2022-02-12
家庭医药·快乐养生(2021年8期)2021-08-30
科学中国人·上半月(2021年3期)2021-07-11
森林工程(2020年6期)2020-12-14
小学阅读指南·低年级版(2020年9期)2020-10-12
江苏农业科学(2020年8期)2020-06-01
文萃报·周五版(2019年18期)2019-09-10
小天使·六年级语数英综合(2019年2期)2019-01-12
儿童故事画报·自然探秘(2017年7期)2018-03-14
为了孩子(孕0~3岁)(2001年3期)2001-06-13
