
PB1-F2 长度改变对H3N2 犬流感病毒影响的研究
2022-05-26 02:57原耀贤李康健马春全李守军
中国预防兽医学报 2022年3期
原耀贤,李 舜,何 威,李康健,马春全,李守军,马 骏*
(1.佛山科学技术学院生命科学与工程学院,广东 佛山 528225;2.华南农业大学兽医学院,广东 广州 510642;3.中国农业科学院上海兽医研究所,上海 201100;4.广州市微生物研究所有限公司,广东 广州 510700)
犬 流 感(Canine influenza,CI)是 由 正 黏 病 毒科,A 型流感病毒属的CI 病毒(CIV)引起犬的呼吸道传染病,主要临床症状为发热、食欲减退、精神萎靡、咳嗽、眼鼻液分泌物增加等[1]。CIV 引起犬死亡的案例较少,多见其与其他病原共感染或继发感染,从而导致患病犬病情加重,严重时会致其死亡[2]。
2007 年,韩国学者首次发现禽源H3N2 CIV 在犬中的流行[3]。2010 年,李守军等在华南地区的犬中分离并报道了一株禽源H3N2 CIV[4]。随后,北京、上海、南京、西安等地均发现禽源H3N2 CIV,且其在当地的犬中普遍存在[5]。目前,我国犬中主要流行的仍是禽源H3N2 CIV,其PB1-F2 蛋白由PB1 基因序列的第二个开放阅读框(ORF)编码的一种非结构性蛋白,是甲型流感病毒的重要毒力因子[6]。目前已证实,PB1-F2 蛋白具有诱导细胞死亡、调节宿主的先天免疫反应和增强病毒聚合酶活性等功能[7]。并且不同宿主来源的流感病毒表达的PB1-F2 蛋白长度不同,且不同长度的PB1-F2 蛋白致流感病毒的毒力也不同[8]。因此,PB1-F2 蛋白可能在跨种间传播的过程中发挥重要作用。
犬是人类的伴侣动物,与人接触密切,它既可以感染哺乳动物流感,也可以感染禽流感病毒(AIV),从而具有成为流感病毒混合器的潜力[9]。但在CI 的研究中,少有研究者涉及对CIV PB1-F2蛋白的研究。基于此,本研究利用反向遗传技术拯救了3 株分别表达不同长度PB1-F2 蛋白的重组病毒,通过比较重组病毒的复制效率、聚合酶活性及其对小鼠的致病性,初步探究了PB1-F2 蛋白对H3N2 CIV 的影响,为该病毒后续致病机制、基因功能的研究奠定基础。
1 材料与方法
1.1 主要实验材料 H3N2 CIV A/canine/Guang-dong/02/2011(H3N2)株;犬肾上皮细胞(MDCK)、人胚肾细胞293T(293T)、荧光素酶报告质粒pPolI-Luci-NP 和海肾荧光素酶表达质粒等由华南农业大学兽医学院外产科教研室提供;E. coliDH5α 感受态细胞购自TaKaRa 公司;9 日龄SPF 鸡胚购自北京梅里亚维通实验动物技术公司;SPF 级6 周龄雌性BALB/c 小鼠,购自广东省医学实验动物中心;所有动物实验均在中华人民共和国科学技术部《爱护和使用实验动物指南》指导下进行。
无内毒素质粒大量提取试剂盒购自OMEGA 公司;无缝克隆试剂盒购自北京全式金生物技术有限公司;定点突变试剂盒购自TaKaRa 公司;TRIzol 试剂购自Invitrogen 公司;胶回收试剂盒购自Axygen 公司;转染试剂LipofectamineTM3000 购自Invitrogen 公司;本实验涉及的细胞培养试剂均购自Gibco 公司;双荧光素酶检测试剂盒购自碧云天公司。
1.2 CIV 各基因节段重组质粒的构建及鉴定 利用
Primer Premier 5.0软件设计了反转录引物CU;pHW2000线性载体的扩增引物(pHW-F/R);带有pHW2000同源臂序列的8个基因节段扩增引物;针对PB1-F2序列的突变引物(表1),均由上海生工生物工程技术服务有限公司合成。提取H3N2 CIV 的RNA,并反转录为cDNA 作为模板,经PCR 扩增出两端带有pHW2000 同源臂序列的8个基因节段。PCR产物均经1%琼脂糖凝胶电泳检测,并利用胶回收试剂盒回收。采用无缝克隆试剂盒,将各目的节段插入至pHW2000载体中,并分别命名为pHW-X,X 为节段名。各重组质粒均经PCR鉴定后再由上海生工生物工程技术服务有限公司测序鉴定,将测序结果与GenBank 中登录的H3N2 CIV各节段进行比对。将测序正确的克隆扩大培养并提取质粒,用于后续试验。

表1 扩增引物及PB1节段突变引物Table 1 Primers for amplification and mutation PB1 fragments
1.3 重组病毒rH3N2、rH3N2-M1R及rH3N2-S11Stop的拯救 参照文献[3]的反向遗传操作技术,将上述构建的8 个重组质粒各400 ng 共转染至293T 细胞,37 ℃5%CO2培养48 h后收获细胞培养液,并将其接种至9日龄SPF鸡胚,37 ℃培养48 h后收获尿囊液,用1%鸡红细胞经血凝试验(HA),测定该病毒的HA 效价。利用TRIzol 提取HA 效价≥23的尿囊液RNA,反转录为cDNA后作为模板,PCR扩增出病毒的8个基因节段并测序,测序结果与原亲本病毒的全基因组比对,确认结果无误后,将拯救的重组亲本病毒命名为rH3N2。
按照定点突变试剂盒的操作流程,以上述cDNA为模板,利用T120C 突变引物PB1-F2-M1R-F/R,PCR扩增出含点突变T120C 的PB1 序列,并通过测序鉴定该序列第二个ORF aa1(起始密码子编码)是否由M突变为R,该突变能使重组病毒不表达PB1-F2 蛋白。将测序正确的序列通过真核表达方法构建突变质粒pHW-PB1-F2-M1R。将7 个基因节段的重组质粒与突变质粒pHW-PB1-F2-M1R 各400 ng 共转染至293T 细胞,按照上述流程拯救出重组病毒,并扩增该病毒的8 个基因节段后测序鉴定,结果无误后将该拯救病毒命名为rH3N2-M1R。
以上述cDNA 为模板,利用C153A 突变引物PB1-F2-S11Stop-F/R,PCR 扩增出含点突变C153A 的PB1 序列,并测序鉴定该序列第二个ORF的aa11是否由S突变为终止氨基酸序列,该突变能使重组病毒表达长11个氨基酸的PB1-F2 蛋白。将测序正确的序列通过真核表达方法构建突变质粒pHW-PB1-F2-S11Stop。将7个基因节段的重组质粒与突变质粒pHW-PB1-F2-S11Stop各400 ng共转染至293T细胞,按照上述流程拯救出重组病毒,并扩增该病毒的8 个基因节段后测序鉴定,结果无误后将该拯救病毒命名为rH3N2-S11Stop。
1.4 重组病毒的效价及遗传稳定性的测定 将拯救的3 株重组病毒分别接种MDCK 细胞,37 ℃5% CO2培养48 h 后收获细胞培养液,并用PBS 10 倍倍比稀释,以10-12~10-1稀释度接种到96 孔板中的MDCK 细胞中,每孔接种100 μL,并根据Reed-Muench 法计算各病毒的TCID50。随后,3 株重组病毒均以MOI 0.1 的剂量感染MDCK 细胞并连续传15 代。分别收集第1代、第5代、第10代和第15代的细胞培养液,提取病毒RNA,反转录为cDNA 并作为模板,采用PCR 扩增3株重组病毒的8个基因节段后测序鉴定。
1.5 病毒RNP 聚合酶活性的测定 通过测定病毒RNP 聚合酶的活性来反映3 株重组病毒转录效率的变化。分别将质粒pHW-PB1、pHW-PB1-F2-M1R、pHW-PB1-F2-S11Stop,与pHW-PB2、pHW-PA 各250 ng,荧光素酶报告质粒pPolI-Luci-NP 和作为内参的海肾荧光素酶表达质粒各25 ng 共转染至12 孔板中的293T 细胞中,每组3 个重复,同时设不转染PB1-F2 质粒的细胞作为阴性对照。37 ℃5% CO2孵育48 h 后,利用双荧光素酶检测试剂盒检测荧光素酶活性,按照公式:RLU1/RLU2 计算双荧光素酶的比值,通过该比值反应重组病毒的聚合酶活性,其中RLU1 为NP 蛋白荧光的反应强度,RLU2 为海肾荧光素酶的荧光反应强度。
1.6 重组病毒在MDCK 细胞中生长曲线的测定 以MOI 0.1 的剂量将3 株重组病毒分别接种到12 孔板中的MDCK细胞中,每组做3个重复,置于37℃5%CO2条件下培养。分别在接种后12 h、24 h、36 h、48 h收集细胞培养液。按照1.4的方法测定病毒的TCID50,绘制重组病毒在MDCK 细胞中的生长曲线。
1.7 感染重组病毒后小鼠体质量的测定及肺脏病理变化的观察 将6 周龄的BALB/c 小鼠随机分成4组,每组8 只。使用无菌DMEM 将3 株重组病毒分别稀释至106TCID50/50 μL。小鼠经CO2麻醉后,通过鼻腔分别接种3 种重组病毒各50 μL,对照组接种同体积的无菌DMEM。每天定时称量小鼠体质量,并观察各组小鼠的临床症状。
在感染后第5 d,每组各迫杀3 只小鼠,采集肺脏,观察其剖检病变,随后用4%多聚甲醛固定24 h,将处理的样品做好标记并由武汉塞维尔生物科技有限公司制作肺组织病理切片。每组余下5 只小鼠称量体质量至第14 d。在试验过程中,若小鼠体质量下降≥20%,则判定为死亡。
1.8 数据分析 利用Graphpad Prism 8.0 统计分析实验数据。采用T-tests 方法比较各重组病毒RNP 的活性;采用Two-way ANOVA 方法比较各组小鼠的生长曲线。当P<0.05 时,则认为差异具有统计学意义[7]。
2 结 果
2.1 CIV 各基因节段重组质粒的构建及鉴定结果
对构建的CIV 各基因节段的8 个重组质粒均经RTPCR 鉴定,结果显示,各节段大小均与预期相符(图1)。对构建的8 个质粒的测序结果显示,测序结果与H3N2 CIV 相应基因节段序列一致。表明各重组质粒均正确构建。
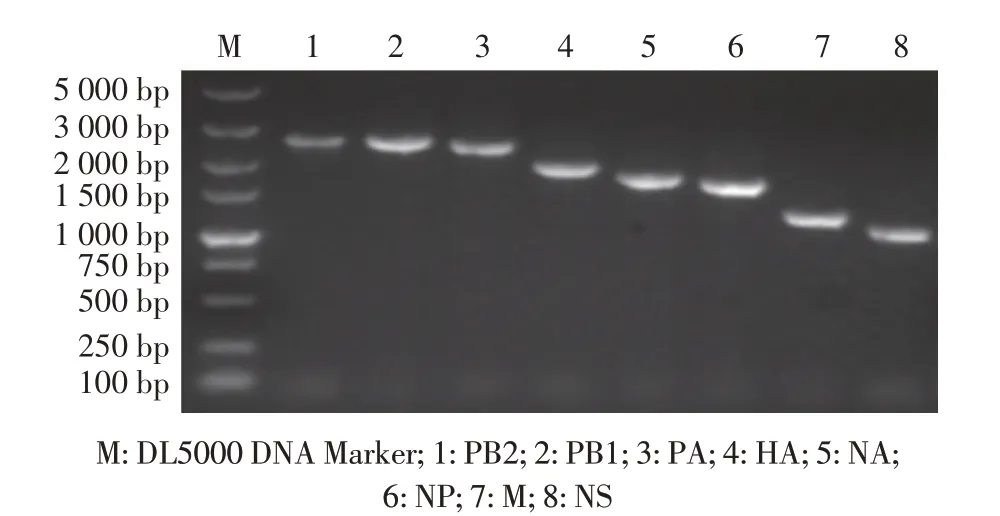
图1 CIV各基因节段重组质粒的鉴定结果Fig.1 Amplification of each gene segment of H3N2 CIV by RT-PCR
2.2 重组病毒的拯救及鉴定结果 将构建的8 个重组质粒共转染293T 细胞,48 h 后收获细胞培养液,接种9 日龄SPF 鸡胚,48 h 后收获尿囊液,测定病毒的HA 效价。经PCR 扩增病毒的8 个基因节段后测序鉴定。构建的突变质粒pHW-PB1-F2-M1R 及pHW-PB1-F2-S11Stop 经测序鉴定正确后,分别替换pHW-PB1 以相同步骤转染、接种鸡胚、测HA 效价并经PCR 和测序鉴定拯救的重组病毒。利用Snap⁃Gene 软件对3 株重组病毒的测序结果与亲本病毒的全基因组进行比对。结果显示,拯救的rH3N2 与亲本病毒序列完全一致;rH3N2-M1R 的PB1 序列第120位碱基由T 变成G,致其第二个ORF 的aa1 由M 突变为R,其他序列与亲本病毒基因组序列均一致;rH3N2-S11Stop 的PB1 序 列 第153 位 碱 基 由C 变 成A,致其第二个ORF 的aa11 由S 突变为终止氨基酸序列,其他序列与亲本病毒基因组序列均一致。结果表明重组病毒rH3N2、rH3N2-M1R 和rH3N2-S11Stop 均构建正确。经HA 测定,rH3N2 和rH3N2-S11Stop 的HA 效价均为25;rH3N2-M1R 的HA 效价为26。
2.3 重组病毒的效价及遗传稳定性的测定结果 根据Reed-Muench 法计算各病毒的TCID50,并将拯救的3 株重组病毒在MDCK 细胞中连续传15 代。收集第1 代、第5 代、第10 代和第15 代的细胞培养液,采用PCR 方法扩增病毒的8 个基因节段并测序分析各重组病毒的遗传稳定性。结果显示,3 株重组病毒 的TCID50分 别 为106.37TCID50/0.1 mL、107.53TCID50/0.1 mL 和106.62TCID50/0.1 mL。
遗传稳定性试验结果显示,3 株重组病毒的第1代、第5 代、第10 代和第15 代的测序结果均相同;其中,各代次的rH3N2 序列均与亲本病毒基因组序列一致;rH3N2-M1R 和rH3N2-S11Stop 各代次的PB1-F2 序列均与2.2 中的各突变序列一致,其他序列均与亲本病毒基因组序列一致。结果表明rH3N2、rH3N2-M1R 和rH3N2-S11Stop 均 能 在MDCK 细 胞 中 稳定传代,且PB1-F2 中的M1R 和S11Stop 突变均具有良好的遗传稳定性。
2.4 病毒RNP 聚合酶活性的测定结果 将重组质粒pHW-PB1、pHW-PB1-F2-M1R、pHW-PB1-F2-S11Stop,分别与pHW-PB2、pHW-PA、荧光素酶报告质粒pPolI-Luci-NP 和海肾荧光素酶表达质粒共转染293T 细胞48 h 后,利用双荧光素酶检测试剂盒检测荧光素酶活性。结果显示,阴性对照组未能检测到聚合酶活性;pHW-PB1-F2-S11Stop 组的聚合酶活性与pHW-PB1 组相比无明显变化;而pHW-PB1-F2-M1R 组的聚合酶活性约为pHW-PB1 组和pHWPB1-F2-S11Stop 组的2 倍,显著高于后两者(P<0.05)(图2)。表明,PB1-F2 的不表达能够显著提高H3N2 CIV 的转录效率(P<0.05),而PB1-F2 长度的截短则对病毒的转录效率基本无影响。

图2 病毒RNP聚合酶活性的检测结果Fig.2 Polymerase activity detection of viral RNP polymerase
2.5 重组病毒在MDCK 细胞中生长曲线的测定 3株重组病毒均以MOI 0.1 的剂量接种MDCK 细胞。每隔12 h 收获细胞培养液,并测定病毒的TCID50。结果显示,3 株重组病毒在MDCK 细胞中的复制趋势基本相似;rH3N2-M1R 在各个时间段的病毒滴度均显著高于rH3N2 和rH3N2-S11Stop(P<0.05);rH3N2-S11Stop在感染后12 h~24 h的病毒滴度显著高于rH3N2(P<0.05),在36 h~48 h的病毒滴度与rH3N2无明显差异(图3)。可见rH3N2-M1R 和rH3N2-S11Stop 在MDCK细胞中的复制效率均高于rH3N2,且rH3N2-M1R的复制效率比rH3N2-S11Stop 高。表明,不同长度的PB1-F2 影响H3N2 CIV 在MDCK 中的复制效率,且以不表达PB1-F2 蛋白的H3N2 CIV 复制效率最高。
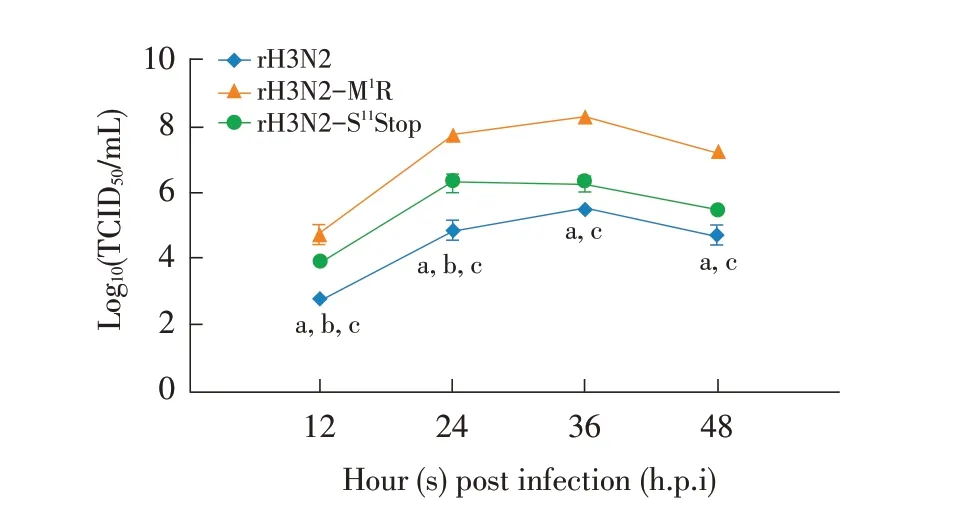
图3 各重组病毒在MDCK细胞中的生长曲线Fig.3 The growth curve of recombinant in viruses on MDCK cells
2.6 重组病毒对小鼠致病性的试验结果
2.6.1 感染病毒后小鼠的体质量测定 用无菌DMEM将3 株重组病毒分别稀释至106TCID50/50 μL,并通过鼻腔感染小鼠。每天定时对各组小鼠称重并观察小鼠临床症状。结果显示,在14 d 实验期内,各组小鼠均无死亡,存活率为100%。感染重组病毒后的小鼠均表现出被毛粗乱、精神不振、食欲减退等临床症状,对照组各小鼠均表现正常。体质量变化曲线显示,rH3N2组与rH3N2-S11Stop组小鼠的体质量在感染后5 d 内持续下降,5 d 后体质量逐渐增加;rH3N2-M1R 组小鼠的的体质量在感染后的3 d 内持续下降,随后逐渐上升;对照组小鼠体质量则稳定上升(图4)。结果表明,H3N2 CIV PB1-F2 长度的改变对小鼠体质量的影响不大,且3 株重组病毒均不具有致死性。
图4 感染各重组病毒后小鼠的体质量的测定结果Fig.4 Body weight change of mice infected with each recombinant virus
2.6.2 小鼠肺脏组织病理学的观察 小鼠感染各重组病毒后的第5 d,每组随机迫杀3 只小鼠,对采集的肺脏制作病理切片并观察。结果显示,rH3N2-M1R 组小鼠气管及血管周边的炎性细胞增多,肺泡间质增宽,出现肺泡融合现象(图5B);rH3N2-S11Stop 组小鼠肺泡出现融合并伴有充血、出血的现象(图5C)。rH3N2 组小鼠气管及血管周边的炎性细胞增多,有肺泡融合现象,肺泡壁增厚,并伴有少量出血的现象(图5D);对照组无明显的病理变化(图5A)。表明,各重组病毒均能对小鼠的肺脏造成轻微病理损伤,且表达不同长度PB1-F2 的H3N2 CIV 对小鼠肺脏造成的病理损伤无明显差异。

图5 感染各重组病毒后小鼠肺脏组织病理变化的观察Fig.5 Lung histopathological changes of mice infected with recombinant viruses
3 讨 论
H3N2 CIV 的流行时间较短,与其他宿主流感病毒发生基因组重组的几率较小,其进化过程相对简单,是研究流感病毒跨宿主传播的理想模型[10]。目前,H3N2 CIV 在国内快速传播,但缺乏关于其毒力因子和跨宿主传播因素的研究,例如针对PB1-F2的研究。因此,本研究通过反向遗传操作技术,拯救了3 株分别表达不同长度PB1-F2 的重组病毒,并初步探究了PB1-F2 长度改变对H3N2 CIV 的影响。
不同宿主来源流感病毒表达PB1-F2 蛋白的长度不同。哺乳动物流感病毒常表达长度少于62 个氨基酸的PB1-F2 蛋白,如人流感pdm09 病毒表达长11 个 氨 基 酸 的PB1-F2 蛋 白[11-12]。然 而,AIV 常 表 达长87 个氨基酸或以上的PB1-F2 蛋白[13]。2013 年的禽流感流行病学调查发现,约61%高致病性H5N1 AIV PB1-F2 的长度发生改变,其PB1 序列第二个ORF 的aa1 由M 突变为R,从而导致PB1-F2 蛋白无法表达[14],这可能是AIV 致病性增强的原因之一。在本研究中,禽源H3N2 CIV 能表达长90 个氨基酸的PB1-F2 蛋白,具有弱致病性,且致死率不高。结合禽源H3N2 CIV 的两个特点,一是该病毒不仅能感染犬类(犬属于哺乳动物),也能感染禽类;二是该病毒在国内的广泛传播,进而推测,随着该病毒在犬中的稳定传播,其PB1-F2 蛋白也可能发生类似突变,从而增强H3N2 CIV 的致病性,并促进CIV 的跨种间传播。
不同氨基酸长度的PB1-F2 蛋白能使甲型流感病毒的致病性产生差异。Vidic 等研究发现,表达长90个氨基酸的PB1-F2 蛋白时,甲型流感病毒的细胞毒性和损伤细胞膜的能力减弱[15]。随后,Deventhiran 等将H3N2猪流感病毒PB1序列第二个ORF的aa1由M突变为R,致使PB1-F2 无法表达,从而增强了该病毒对禽类的致病性[16]。本研究结果表明,M1R 突变后的PB1-F2 能够显著提高H3N2 CIV在MDCK细胞中的复制效率及病毒的转录效率(P<0.05),这与前人的研究结果相一致[15];本研究结果还表明,S11Stop 突变后表达11 个氨基酸的PB1-F2 能够提高H3N2 CIV 在MDCK 细胞中的复制效率,但并不影响病毒RNP 的活性即对病毒的转录效率无明显影响,表明PB1-F2 蛋白在H3N2 CIV 的转录复制过程中起着重要作用,且截短后尤其是长11 个氨基酸的PB1-F2 蛋白可能具有与全长PB1-F2 蛋白相似的功能,并与病毒的复制功能相关。有研究结果显示,PB1-F2 蛋白对IAV 感染小鼠后造成的肺脏病理损伤基本无影响[17]。本研究中,3 株重组病毒对小鼠的致病性无明显差异,且均对小鼠不具有致死性;虽然携带不同长度PB1-F2 蛋白的H3N2 CIV 均能对小鼠肺脏造成病理损伤,但是各感染组之间的病理损伤无明显差异,表明表达不同长度PB1-F2 蛋白的H3N2 CIV 仍为低致病性流感病毒。
综上所述,本研究分别拯救了3 株表达不同长度PB1-F2 蛋白的重组CIV,首次证明了PB1-F2 长度的改变能影响H3N2 CIV 在细胞中的转录与复制过程,但对病毒在小鼠中致病性的影响较小。本研究为阐明PB1-F2蛋白在H3N2 CIV中的致病作用提供了参考,也为该病毒病的防控提供了一定参考依据。
猜你喜欢
铁道建筑技术(2022年10期)2022-10-28
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
中国典型病例大全(2022年12期)2022-05-13
中国典型病例大全(2022年9期)2022-04-19
天津医科大学学报(2021年4期)2021-08-21
江西农业学报(2021年4期)2021-04-20
广东蚕业(2021年1期)2021-03-18
三农资讯半月报(2020年11期)2020-06-21
山东医药(2020年9期)2020-05-20
