
三维电镜在脑干耳蜗核神经元形态学研究中的应用
2022-04-22 06:03周佳蕾盛海斌王皓煜王方方皓1华云峰1
上海交通大学学报(医学版) 2022年2期
周佳蕾,盛海斌,王皓煜,鲁 岩,王方方,吴 皓1,2,,华云峰1,2,#
1.上海交通大学医学院附属第九人民医院耳鼻咽喉头颈外科,上海 200011;2.上海交通大学医学院耳科学研究所,上海市耳鼻疾病转化医学重点实验室,上海 200125;3.上海精准医学研究院,上海 200125;4.上海交通大学医学院附属儿童医院耳鼻咽喉头颈外科,上海 200062
耳蜗核(cochlear nucleus,CN)是哺乳动物听觉中枢的最低级核团,其对称分布于脑桥和延髓交界处,接受同侧耳蜗听神经投射。既往研究[1]发现,CN 神经元的轴突亦可投射到同侧及对侧的上橄榄核复合体(superior olivary complex,SOC)。因此,除能够对来自耳蜗的听觉信号作简单中继外,CN 还可作为双耳信号处理的起始核团,在声源定位、双耳增益校准及回声抑制等双耳功能中发挥重要作用。
连接耳蜗听觉毛细胞的听神经束在进入CN 后会形成分叉,分别投射至CN 的3 个亚区,即前腹侧耳蜗核(anteroventral cochlear nucleus,AVCN)、后腹侧耳蜗核(posteroventral cochlear nucleus,PVCN)以及背侧耳蜗核(dorsal cochlear nucleus,DCN)。研究[2-3]显示,位于AVCN 中的丛细胞(bushy cell,BC)是耳蜗听神经的主要支配对象之一,其胞体可与听神经形成根蕾状(endbulb of Held)突触。且相关研究[4-6]发现,根蕾状突触可能赋予了丛细胞与听神经放电产生锁相的特征,从而被认为是一种重合检测器,用于判断丛细胞上多个听觉信号间的相关性。同时,相邻丛细胞可形成具有同步放电特性的细胞簇[7-9],通过听神经上连体的根蕾状突触或丛细胞间的电耦合来实现相邻丛细胞的同步放电。此外,丛细胞还可接收来自AVCN 内部卫星细胞及来自同侧DCN、对侧CN、SOC 和体感神经的直接或间接投射的调控[1,7,10-13],而完整的投射至丛细胞的神经突触支配模式及特异性有待进一步明确。另有动物实验[5,14-15]发现,耳聋以及异常的听觉经历均可导致腹侧耳蜗核(ventral cochlear nucleus,VCN)发生突触可塑性变化,而这些改变对VCN 神经环路产生的具体影响尚不清晰。
研究[16]显示,脑组织神经环路的解析需要足够的尺度及必要的分辨率,才能获得单个突触分辨率的完整神经连接图谱。但目前,针对CN 神经环路的结构学研究方法仅局限在稀疏荧光标记和光学显微成像,由于其分辨率的限制,只能行孤立神经元的重构和分析。而孤立重构的神经元虽可用于形态分类和功能预测,但对明确神经环路中同类神经元的分工以及最小功能微环路的组成贡献较为有限。近年来,三维电镜技术在通量上的提升使得大尺度、高分辨重构生物样品成为可能,该新技术已逐渐用于各类脑组织和感官器官的神经连接组学研究[17-19]。基于前期对大脑皮层[20-21]和耳蜗神经组织[22-24]的三维电镜研究基础,本课题组拟制备小鼠完整CN 的电镜样品,通过X 射线显微镜和连续切片三维扫描电镜对VCN 神经环路进行跨尺度的形态学研究尝试,为系统开展CN神经连接组学研究奠定基础。
1 对象和方法
1.1 实验动物
7~8周龄SPF级正常听力的雄性CBA/Ca小鼠5只[购于上海西普尔-必凯实验动物有限公司,实验动物生产许可证:SCXK(沪)2017-0012],体质量为200~250 g。小鼠饲养于上海交通大学医学院附属第九人民医院实验动物中心的标准饲养笼中,实验动物使用许可证:SYXK(沪)2016-0016。温度约25 ℃、湿度60%~70%、12 h 光照与黑暗交替条件下,自由进食、饮水。本研究涉及的动物实验通过上海交通大学医学院附属第九人民医院动物伦理委员会审批(审批号:SH9H-2021-A728-1),所有动物的相关操作均遵循《实验动物管理条例》。
1.2 CN的固定和取材
小鼠经腹腔注射水合氯醛(500 mg/kg)进行麻醉后,用微量注射泵(以9 mL/min 的泵速)经心脏先后泵入15 mL 二甲胂酸钠缓冲液(0.15 mol/L)和30 mL 混合固定液(2%多聚甲醛、2.5%戊二醛及0.08 mol/L 二甲胂酸钠缓冲液,pH=7.4)行全身固定。随后,处死小鼠并进行断头,快速地从其颅骨中解剖、分离出脑组织(需注意在分离脑组织时,应使用精细眼科剪切断内听道至脑干间的听神经,以免破坏CN组织的结构),并浸没在混合固定液中,于4 ℃下静置24~48 h。取出脑组织后,将其转移至装有二甲胂酸钠缓冲液(0.15 mol/L)的解剖皿中(冰上)。于解剖显微镜下小心剔除部分小脑,暴露出脑干及其表面的CN。最终,从脑干中完整分离出CN。
1.3 CN样品的重金属块染
参照已建立的电镜样品制备流程中的染色方法[20]对采集的CN 样品进行染色,具体步骤如下:①使用二甲胂酸钠缓冲液(0.15 mol/L)清洗CN 样品2 次,每次30 min;随后,依次于锇酸(2%)、亚铁氰化钾(2.5%)、锇酸(2%)中分别浸没2、1.5、1 h。②使用二甲胂酸钠缓冲液(0.15 mol/L)和去离子纯净水清洗样品各1 次,每次30 min;再将样品转移至硫代甲肼(1%水溶液)和锇酸(2%水溶液)中浸泡1、1.5 h,并于每步之后用去离子纯净水清洗2次,每次30 min。③将样品浸没在乙酸铀(1%水溶液)中,于4 ℃过夜;次日转移至50 ℃烘箱中静置2 h,并用去离子纯净水清洗2 次,每次30 min。④再将样品转移到新鲜配制的天门冬氨酸铅溶液(0.03 mol/L)中,于50 ℃浸泡2 h,用去离子纯净水清洗2 次,每次30 min。
1.4 CN核样品的脱水和树脂包埋
采用梯度乙醇(50%、75%、90%和100%)及无水丙酮对上述已完成染色的CN 样品进行脱水,具体如下:①样品于预冷(4 ℃)的50%、75%、90%乙醇中脱水,每个梯度浸泡30 min。②将样品转移至100%乙醇中,于室温脱水30 min。③于室温下用无水丙酮脱水3次,每次45 min。
将脱水后的样品浸没于无水丙酮与低黏度Spurr树脂的混合液(体积比1∶1)中,于旋转仪上(转速为5 r/s)过夜混匀(离心管盖保持打开,以便丙酮挥发)。次日,将样品转移至纯树脂中静置8~12 h(离心管关盖),而后再置于模具中,70 ℃烘箱(需预热)内高温聚合72 h以上,完成包埋。
1.5 CN 样品的X 射线显微成像及其目标区域的定位和修样
利用TRIM2 修块机(Leica,美国)将脱水、包埋后的CN 样品修成横断面约为2.0 mm×2.0 mm 的长方体,并用强力胶将其固定于铝钉底座上,通过X射线显微镜进行扫描、成像。使用图像处理软件Dragonfly(Objective Research Systems,加拿大)对获得的图像进行三维重构,实现三维体积数据的可视化,用于对样品染色效果和均一度的初步评估以及CN目标区域的定位。
使用修块机去除CN 目标区域周围多余的树脂,利用UC7 超薄切片机(Leica,德国)对其表面进行抛光处理,再用Ted Pella-208C 离子溅射仪(Ted Pella,美国)对其做喷碳(厚度10 nm)处理后,用Gemini300 扫描电镜(Zeiss,德国)进行低分辨率快速预扫描(像素为1.59 μm,驻留时间为1.0 μs),用于和X射线显微成像后重构的图像进行比对,确定样品表面在CN的具体位置。
1.6 目标区域的三维电镜图像采集、三维重构和超微结构分析
将预扫的电镜图像与X射线显微成像后重构的三维图像进行结构比对,进一步确定目标区域(VCN的丛细胞簇区域)位置,必要时可通过超薄切片机进行再次修样,原则上样品横断面大小不大于600 μm×600 μm。而后,利用EM ACE600 高真空镀膜仪(Leica,德国)对修样抛光后的CN 样品表面行真空喷金处理(厚度30 nm),采用连续切片扫描电镜——Gemini300 场发射扫描电镜(Zeiss,德国)[内置了3ViewXP 超薄切片机(Gatan,美国)和Onpont背散射电子探测器(Gatan,美国)]对VCN 目标区域进行三维图像采集。
利用本课题组编写的MATLAB(Mathworks,美国)脚本对连续切片扫描电镜采集的原始图像的对比度与亮度进行匹配、三维对齐重构,并将数据集上传至开源图像标记工具WEBKNOSSOS以实现在线浏览和标记[25],包括对丛细胞树突的追踪标记以及支配于丛细胞胞体表面各类神经末梢突触的体积标记。所标记的结构的三维可视化和量化均使用MATLAB 脚本完成。
2 结果
2.1 CN样品的显微观察及其三维成像信息
在解剖显微镜下观察到,小鼠CN 位于脑桥和延髓交界处的两侧,呈现椭圆状隆起,其长轴自前下方至后上方依次为VCN 和DCN,而AVCN 和PVCN 分别位于耳蜗听神经残端的前背侧和后背侧区(图1)。通过进一步解剖分离,我们获得了包括完整CN 在内的脑干组织,尺寸约为2.0 mm×2.0 mm×3.0 mm。最终,共制备了5 个CN 样品用于后续的成像分析,X射线显微镜及连续切片扫描电镜三维成像信息见表1。
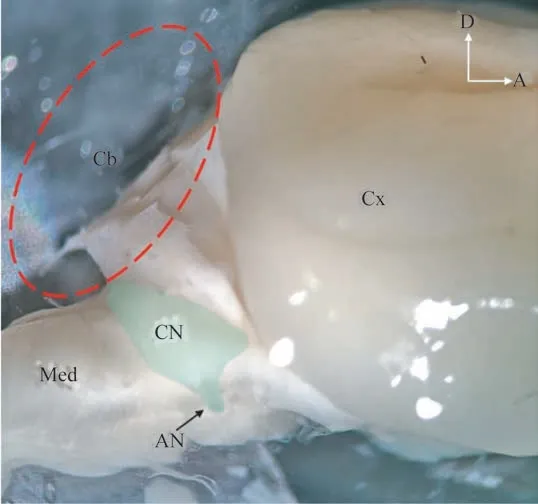
图1 显微镜下观察小鼠CN的解剖位置Fig 1 Observation of anatomical location of CN in mice under microscope
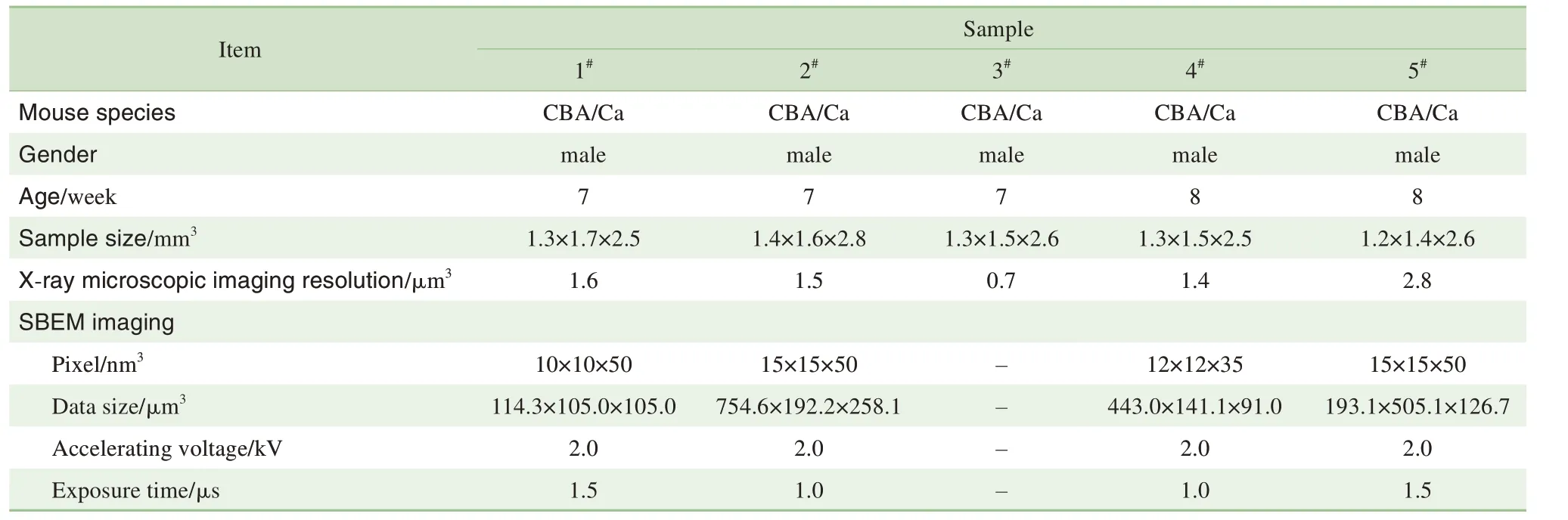
表1 小鼠CN样品的三维成像信息Tab 1 Three-dimensional imaging information of mouse CN samples
2.2 CN样品的X射线显微图像及其解剖分区
采用X 射线显微镜对CN 样品进行断层扫描,对所得图像进行三维重构,得到具有细胞分辨率的CN三维图像。CN 样品尺寸为1.3 mm×1.5 mm×2.6 mm、体素0.7 μm。根据听神经的特征分布和细胞密度,我们对CN 样品的各个亚区进行了划分,其中VCN、DCN 以及听神经(auditory nerve,AN)的空间分布如图2A 所示。从样品的矢状面(图2B)和冠状面(图2C)来看,听神经主干和CN 神经元胞体清晰可见;相比于其神经毡,由髓鞘包裹的听神经束经锇酸染色后呈现出较高的染色密度(即亮信号),而细胞胞体的染色密度相对较低(即暗信号),且样品的解剖分区清晰可辨:AVCN 位于听神经束较短的分支处,PVCN 位于长分支的起始端;而DCN 则位于长分支的末端,与PVCN由细胞密度较低的颗粒细胞亚区间隔。

图2 CN样品的X射线显微成像Fig 2 X-ray microscopic imaging of CN samples
2.3 X射线显微图像和扫描电镜图像的结构比对
在正式采集连续切片扫描电镜的数据之前,本研究将修样后CN 样品表面的电镜预扫描图像与已采集的X射线显微成像图像进行比对,结果显示,样品表面的电镜预扫描图像(图3A)与X 射线显微成像的相应层面图像(图3B)在特征结构上(胞体及听神经的特征分布)可完全吻合。继而说明,通过X射线显微成像可精确定位目标区域;同时,在其引导下可进一步优化连续切片扫描电镜采集的目标区域(VCN的丛细胞簇区域)。
2.4 VCN丛细胞及根蕾状突触的三维重构
采用连续切片扫描电镜采集并三维重构的VCN 丛细胞簇区域的高分辨三维图像数据集(图4A),所 采集的CN 体积为114.3 μm×105.0 μm×105.0 μm,三维分辨率为10 nm×10 nm×50 nm。在此成像条件下,突触的特征结构(包括神经递质囊泡、突触后致密物)清晰可辨(图4B),可作为突触识别的依据。随后,对丛细胞的树突结构进行追踪标记,结果(图4C)显示丛细胞有1 个树突,树突末端有丰富的分支,与文献[7]报道相一致。从丛细胞胞体上各类突触的体积标记(图4C、4D)和三维重构(图4E、4F)来看,该细胞胞体受5 个大小相当的根蕾状突触支配,共形成了348 个独立的突触活跃区(active zone);此外,其还受来自59 个非听神经来源的神经末梢的97 个突触支配。
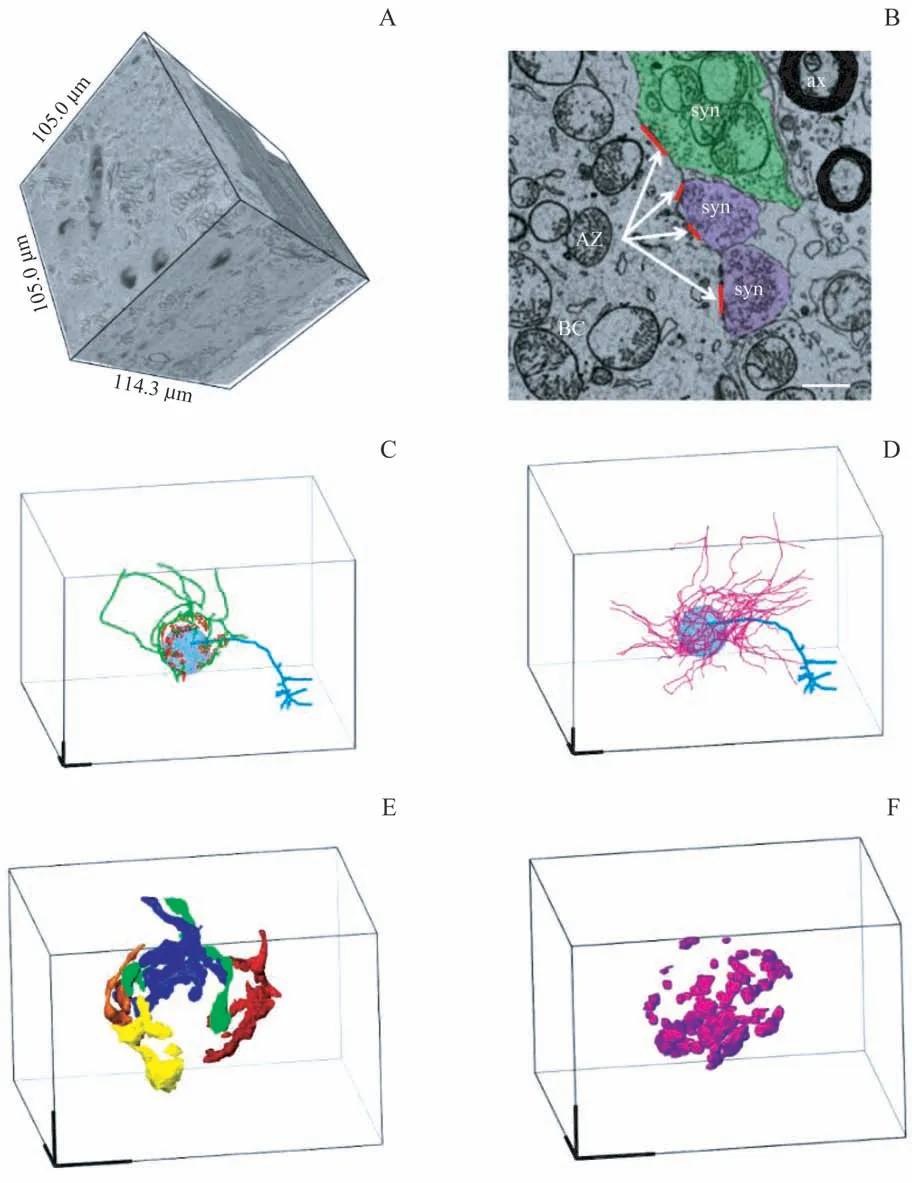
图4 VCN丛细胞簇的三维电镜图像数据集及相关神经结构的标记与重构Fig 4 Labeling and reconstruction of three-dimensional electron microscopic image dataset of VCN bushy cell cluster and related neural structures
3 讨论
听觉系统具有持续、高频、高保真传递信号的特点,这与一系列具有特殊结构和生理功能的突触紧密相关,其中包括耳蜗带状突触、CN 根蕾状突触以及斜方体内侧核(medial nucleus of the trapezoid body,MNTB)的花萼状巨突触(calyx of Held),因此对各类听觉突触的超微结构及电生理特性的研究对理解突触传递的机制具有重要的意义。对于听觉突触超微结构的研究,既往主要是通过透射电镜对手动收集的连续超薄切片进行成像及重构。由于受到切片质量和稳定性的影响,其重构体积(如厚度)非常有限,常常仅有1~2 个完整细胞。近年来,高通量三维电镜技术的发展大大提高了神经组织三维图像采集的效率,已在不同脑组织神经连接组学的研究中得到了越来越多的应用。如:SCHMIDT 等[26]利用三维电镜成像量化分析大鼠内侧嗅皮层中局部单个轴突的突触前突触类别,揭示了轴突径路长度依赖的突触分类现象;KARIMI 等[27]利用三维电镜成像量化小鼠大脑皮层不同层梭形细胞顶树突接受不同类型突触的独特输入图谱,揭示不同梭形细胞在其顶树突上所接受调控的神经连接组学规则;GOUR 等[21]利用三维电镜成像量化并揭示发育过程中小鼠体感皮层抑制性中间神经元突触产生的环路模式。同时,在神经环路超微结构的解析方面,光学成像的空间分辨率有限,电生理记录局限于单个脑组织切片中的少量连接,传统电镜则受限于成像尺寸;而高通量三维电镜成像具备大尺寸、高分辨率以及可三维成像等优势,可以获得树突径路上不同部位和不同类别的突触密度、轴突径路上所形成的突触分布图谱以及神经元内细胞器结构等超微结构的定量信息,因而能达到光学成像、电生理记录以及传统电镜等成像手段无法实现的目的。在听觉系统研究领域,三维电镜也越来越多地被应用于耳蜗毛细胞和NMTB 中花萼状巨突触的形态及发育研究[28-31],但将其应用于CN 的研究尚未有报道,其瓶颈之一即为相关组织电镜样品的制备。而本研究在前期工作[20]的基础上,首次实现了小鼠完整CN 三维电镜样品的制备,相比之前的工作其具有更大的体积以及组织的特异性。同时,本研究利用X射线显微成像,对其染色效果进行评估并对其目标区域进行初步定位;成功采集了VCN 丛细胞簇区域的三维电镜图像,并在突触分辨率下实现了VCN 中丛细胞及其表面的根状蕾突触和非听神经来源突触的追踪和重构。因此,本研究不仅证实了对小鼠CN 开展三维电镜研究的可行性,还提供了具体的实验细节,包括CN 样品的制备流程、X 射线显微成像辅助的目标区域定位以及三维电镜的成像条件。
作为VCN 的主神经元,丛细胞可通过根蕾状突触接受来自耳蜗听神经的信号,然而具有不同自发放电速率以及蛋白表达的耳蜗听神经在丛细胞上的整合方式目前仍不明确[8,32-33]。当外周听力受到损伤,丛细胞上的根蕾状突触会发生一系列显著的结构改变并伴随有突触支配力度的下降,而这样的改变是否具有听神经亚型的特异性、如何影响丛细胞对听觉信号的响应等有待深入的探索分析。已有证据表明,在成年正常听力以及老年性聋的小鼠CN丛细胞上,特异表达钙视网膜蛋白(calretinin)和非表达该蛋白的听神经可支配不同的丛细胞,但后者的占比在老年性聋小鼠中有大幅下降[34]。而在本研究中,我们对丛细胞的三维电镜重构不仅可以提供更具体的量化指标,包括单个丛细胞上根蕾状突触和活跃区的数量以及非听神经来源的突触支配信息,同时其更大的重构体积还可以将突触的量化扩展到丛细胞的树突结构或是可在丛细胞簇层面上开展进一步的研究。未来,如能将其他研究手段(如免疫金染色等)与三维电镜成像技术结合,将有望回答更多的关于CN神经环路的未解之疑。
耳鸣、耳聋、异常的听觉经历常伴随有CN 突触可塑性的改变[35-38]。利用三维电镜量化这一改变并在神经环路层面揭示其对听功能的影响,将有助于对相关疾病病理机制的研究。目前,对于因耳蜗严重畸形或蜗神经发育不良而无法接受人工耳蜗植入的患者,听性脑干植入是听觉重建的唯一手段[39],即可通过植入电极直接刺激CN 组织产生人工听觉。由于当前研究者们对相关神经环路的结构、听神经损伤后引起的CN 神经环路可塑性改变等都缺少足够的认识,使得精准刺激CN 神经元、对声信号进行有效编码的能力极为有限。但随着三维电镜技术的不断发展,其或将推动CN 神经环路的相关研究,从而填补这一空白。
猜你喜欢
无线互联科技(2022年4期)2022-05-11
当代陕西(2022年4期)2022-04-19
城市道桥与防洪(2022年1期)2022-02-25
摄影世界(2022年1期)2022-01-21
中国听力语言康复科学杂志(2021年6期)2021-12-21
中国听力语言康复科学杂志(2021年6期)2021-12-21
北京航空航天大学学报(2021年9期)2021-11-02
河北工业大学学报(2021年4期)2021-09-23
全球定位系统(2021年3期)2021-08-07
发明与创新·中学生(2019年3期)2019-03-20
