
蛋白酶制剂对虾酱发酵过程中理化性质和微生物区系的影响
2021-06-04 02:17:32李文亚郑晓卫桑亚新孙纪录
食品科学 2021年10期
李文亚,刘 洋,李 宁,郑晓卫,桑亚新,孙纪录,*
(1.河北农业大学食品科技学院,河北 保定 071000;2.中粮营养健康研究院有限公司,营养健康与食品安全重点实验室,北京 102209)
虾酱为毛虾等小型虾类及虾加工副产物经腌制、捣碎、发酵制成的糊状食品[1]。虾酱在东亚和东南亚很流行,是一种传统调味品[2-3]。虾酱味道鲜香,营养丰富,富含蛋白质、多种维生素及矿物质[4-5]。虾酱的发酵过程主要涉及来自虾类和细菌的蛋白酶对原料中蛋白质的水解,释放出氨基酸和肽,从而赋于产品特定的理化性质。因此,研究虾酱在发酵过程中的微生物多样性及其与主要理化性质的相关性,对了解参与发酵的优势微生物,以改进虾酱生产工艺非常重要[6]。
采取传统发酵方法生产虾酱,发酵时间长,虾酱质量不稳定。为了改进虾酱生产工艺,缩短发酵周期,稳定产品质量,已经开始尝试添加外源蛋白酶制剂。在虾酱发酵过程中加入各种蛋白酶,加速虾体蛋白质的分解转化,缩短发酵时间,是加酶发酵法制备虾酱的目标[1]。谢主兰等[7]通过对虾酱酶法模拟加工过程中各工序的细菌学分析,得出杀菌工艺条件直接影响虾酱的风味品质与细菌学品质。纂翠华等[4]以小型蜢子虾为原料,利用碱性蛋白酶、中性蛋白酶、木瓜蛋白酶探究虾酱酶法发酵工艺条件,最终确定最佳酶碱性蛋白酶及最佳工艺条件。刘树青等[8]通过外加酶制剂制得了虾酱,进行了工艺研究,考察酶种类、用盐量及温度对于虾酱理化性质的影响。在已有研究中,对虾酱中加入酶制剂往往只是工艺上的研究,而外加酶制剂对虾酱微生物区系和生物胺的影响尚鲜有报道。近年来,基于16S rDNA基因的高通量测序已被广泛用于分析食品发酵体系中的微生物组成[9]。利用高通量测序分析虾酱的微生物多样性可行,而且十分必要。
因此,为了阐明蛋白酶制剂对虾酱发酵过程中理化性质和微生物区系动态变化的影响,本研究使用高效液相色谱等方法测定虾酱的生物胺、氨基酸态氮、挥发性盐基氮(total volatile base nitrogen,TVB-N)等重要理化指标,通过高通量测序分析虾酱的细菌群落组成和结构,使用Spearman分析优势菌群与理化性质之间的相关性。旨在为进一步改善虾酱的酶法辅助发酵工艺提供理论依据。
1 材料与方法
1.1 材料与试剂
虾购自河北保定市科技市场,品种为蜢子虾,产地为河北黄骅。
蛋白酶制剂 北京润泽康生物科技公司;硫代巴比妥酸 国药集团化学试剂有限公司;乙腈(色谱纯)福晨(天津)化学试剂有限公司;丙二醛乙缩醛、生物胺标准品(组胺二盐酸盐、酪胺盐酸、2-苯乙胺、色胺、尸胺二盐酸盐、腐胺、精胺、亚精胺)、丹磺酰氯(纯度≥99%) 上海源叶生物科技有限公司;E.Z.N.ATMMag-Bind Soil DNA Kit 美国Omega公司;Qubit 2.0 DNA检测试剂盒 美国Life公司;TaqDNA Polymerase 美国Thermo公司;Agencourt AMPure XP美国Beckman公司。
1.2 仪器与设备
Breeze高效液相色谱仪 美国Waters公司;3-18K型高速冷冻离心机 美国Sigma公司;HD-850桌上式洁净工作台 苏州净化设备有限公司;PHS pH计 合肥桥斯仪器设备有限公司;721G可见分光光度计 上海仪电分析仪器有限公司;XHF-DY高速分散器 山东博科生物产业有限公司;Qubit®2.0荧光计 美国Invitrogen公司;ABI GeneAmp®9700型聚合酶链式反应(polymerase chain reaction,PCR)仪 美国Bio-Rad公司。
1.3 方法
1.3.1 虾酱的制作
挑选新鲜蜢子虾,洗净沥水并称质量,捣碎搅拌,打浆后放入发酵罐(每罐200 g),同时加入蜢子虾质量20%的食盐,用2 层纱布封口,室温下自然发酵(2019年9月20日—10月17日,河北省保定市)。加酶组虾酱在添加食盐的同时加入0.3%中性蛋白酶[10]。
1.3.2 理化指标分析
1.3.2.1 pH值测定
在试管中,将2 g虾酱和10 mL蒸馏水混合,振荡2 min,充分混匀,室温静置5 min,取出上清液,测定pH值,每种样品重复测定3 次[11]。
1.3.2.2 TVB-N值的测定
参照GB 5009.228—2016《食品中挥发性盐基氮的测定》,采用半自动凯氏定氮法。
1.3.2.3 丙二醛的测定
参照GB 5009.181—2016《食品中丙二醛的测定》,采用分光光度法。
1.3.2.4 氨基酸态氮的测定
参照GB/T 5009.235—2016《食品中氨基酸态氮的测定》,采用比色法。
1.3.2.5 生物胺的测定
采用高效液相色谱法,色谱柱为WondaSil C18(4.6 mm×250 mm),流动相A为0.1 mol/L乙酸铵,流动相B为乙腈,柱温40 ℃,流速1 mL/min,紫外检测器波长254 nm,进样量20 μL。采用梯度洗脱程序,洗脱程序参照李大伟等[12]的方法。
1.3.3 微生物多样性分析
1.3.3.1 DNA提取和PCR扩增
根据E.Z.N.A.®Soil DNA Kit说明书进行虾酱中微生物群落总DNA抽提,使用1%琼脂糖凝胶电泳检测DNA的提取质量,使用NanoDrop2000测定DNA浓度和纯度;使用338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)对16S rRNA基因V3-V4可变区进行PCR扩增,扩增程序:95 ℃预变性3 min,27 个循环(95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s),然后72 ℃终延伸10 min,最后在4 ℃进行保存。PCR体系:5×TransStart FastPfu缓冲液4 μL,2.5 mmol/L dNTPs 2 μL,上游引物(5 μmol/L)0.8 μL,下游引物(5 μmol/L)0.8 μL,TransStart FastPfuDNA聚合酶0.4 μL,模板DNA 10 ng,补足至20 μL。每个样本3 个重复[13]。
1.3.3.2 Illumina MiSeq测序和数据处理
将同一样本的PCR产物混合后,使用2%琼脂糖凝胶回收PCR产物,利用AxyPrep DNA Gel Extraction Kit进行回收产物纯化,2%琼脂糖凝胶电泳检测,并用Quantus™Fluorometer对回收产物进行检测定量。使用NEXTFLEX®Rapid DNA-Seq Kit进行建库,原始数据上传至NCBI SRA数据库。
使用Trimmomatic软件对原始测序序列进行质控,使用FLASH软件进行拼接。使用的UPARSE软件(version 7.1),根据97%的相似度对序列进行可操作分类单元(operational taxonomic units,OTU)聚类并剔除嵌合体。利用RDP classifier对每条序列进行物种分类注释,比对Silva数据库(SSU128),设置比对阈值为70%。
1.4 统计学分析
使用Origin 2017处理理化指标,结果用折线图呈现;利用mothur计算不同随机抽样下的α多样性指数,利用R语言工具制作曲线图;利用LEfSe进行物种差异分析;使用SPSS(Version 22)进行相关性分析。
2 结果与分析
2.1 虾酱发酵期间的理化指标分析

图1 虾酱在发酵期间的理化指标变化Fig.1 Changes in physicochemical properties of shrimp paste during fermentation
由图1A所示,发酵期间,两种虾酱的pH值均呈波动上升趋势,可能是由于在微生物的作用下,部分氨基酸、蛋白质等被分解为碱性物质,如氨、生物胺等[14];两种虾酱的pH值差异不显著,该结果与尹超[15]的研究类似。SB/T 10525—2009《虾酱》规定虾酱中TVB-N值不得超过450 mg/100 g[16]。由图1B可见,在发酵过程中,两种虾酱的TVB-N值均呈上升趋势,加酶组TVB-N值上升较多,但其含量远低于限量标准。吴小禾等[11]检测成品黄骅虾酱所测得TVB-N值最高为401 mg/100 g,远高于本研究数值。由图1C可以看出,发酵期间,两种处理的虾酱中,丙二醛含量都在逐渐降低。氨基酸态氮是评价鱼露、酱油、虾酱等产品发酵程度的首要指标。由图1D可以看出,在发酵期间,加酶组的氨基酸态氮含量在7 d后一直高于对照组,在一定程度上可以缩短虾酱的发酵周期。但是,两组虾酱中氨基酸态氮含量差异并不大。这可能是因为,氨基酸在虾酱中既是外加蛋白酶制剂和微生物产生的蛋白酶分解产物,同时又是许多微生物可以吸收利用的氮源类营养物质。虾酱作为一种自然发酵的产品,其中的微生物区系复杂多变,使得发酵过程受到许多因素的影响,氨基酸态氮的积累速度也是如此。为了确保主要的发酵菌群在虾酱发酵过程中一直处于优势地位,以稳定虾酱的质量稳定性,功能微生物的强化发酵技术近年来已开始引入虾酱研究领域。因此,不同的加工方式会影响虾酱的理化性质。
2.2 虾酱发酵期间的生物胺分析

图2 虾酱在发酵期间的生物胺变化Fig.2 Changes in biogenic amine content in shrimp paste during fermentation
对两种处理的虾酱样品进行了8 种生物胺的测定,分别是色胺、腐胺、尸胺、苯乙胺、组胺、酪胺、精胺和亚精胺。其中检测出的只有色胺和酪胺。由图2可见,在虾酱发酵过程中,加酶组和对照组中色胺含量不断升高,酪胺呈现微小波动变化;加酶组和对照组并无明显差异,这说明外加蛋白酶制剂并不会影响虾酱中生物胺的含量。此外,与安全限量相比,两种虾酱中生物胺含量都很低,符合质量安全要求。这与两种虾酱中产生物胺细菌属的相对丰度较低一致。如,假单胞菌属(Pseudomonas)能够产生物胺,其在对照组和加酶组两组虾酱中相对丰度均比较低。四联球菌属(Tetragenococcus)在发酵食品中较为常见,且与生物胺的产生有关,但其在本研究中的两种虾酱中含量也很低。
2.3 虾酱中的微生物多样性分析
2.3.1 虾酱样品的α多样性指数

表1 虾酱发酵过程中微生物多样性Table 1 Microbial community diversity indices in shrimp paste during fermentation
在全部虾酱样品中共获得313 864 条有效序列和1 279 个OTU。如表1所示,对照组的ACE指数和Chao1指数大于加酶组,说明对照组中物种丰富度比较高。估算微生物多样性的常用指数为Shannon和Simpson指数,如表1所示,对照组的微生物多样性高于加酶组。对照组比加酶组虾酱样品中的细菌种类丰富。随着发酵的进行,对照组样品中细菌的多样性在后14 d呈现减少的趋势,加酶组样品在前14 d呈增加趋势,而后逐渐减少。每个样品的覆盖率,即抽样完整性指数均在0.99以上,表明测序结果获得了足够可靠的数据质量[17-18]。

图3 两种虾酱样品细菌稀疏曲线和OTU丰度曲线Fig.3 Rarefaction curves and OTU rank curves for bacterial community in shrimp paste samples with different fermentation times
由图3A可见,基于OTU水平的样品相似性为97%的稀疏性曲线分析,表明测序深度符合测序和分析的要求。上述虾酱样品的曲线均未趋于平坦,这表明虾酱样品中还有细菌继续生长的空间,说明虾酱中的细菌种类十分丰富。从图3B可以看出,对照组的曲线更长也更加平滑,这说明相较于加酶组,在对照组中有更加丰富的细菌种类,并且细菌的种类分布也更加均匀。而加酶组无论是细菌的丰度,还是菌种分布的均匀度,都相对较小。这可能是因为加入的蛋白酶制剂会加速分解大分子蛋白质,在虾酱发酵初期,外加的蛋白酶制剂迅速降解原料中的蛋白质,导致在加酶组中芽孢杆菌属(Bacillus)相对丰度明显增加,成为一种新的优势菌属,因而导致加酶组菌种分布均匀度及细菌丰度相应有所下降。
2.3.2 虾酱细菌菌群门水平分布

图4 虾酱细菌组成在门水平上的分布Fig.4 Bacterial community composition in shrimp paste at the phylum level
两组虾酱样品中共鉴定出29 个菌门,由图4可见,主要为变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)、厚壁菌门(Firmicutes)、软壁菌门(Tenenricutes)、放线菌门(Actinobacteria)和Epsilonbacteraeota。在虾酱的整个发酵过程中,优势门一直为变形菌门,这与泰国传统发酵虾酱中优势门为厚壁菌门有所不同[6],而与连云港虾酱中优势菌门结果相似[13],由此可见,不同地区的虾酱微生物区系有所差异。在发酵之初(C0),各菌门依照相对丰度由高到低为变形菌门(51.83%)、拟杆菌门(23.46%)、Epsilonbacteraeota(6.63%)、厚壁菌门(5.78%)、放线菌门(5.77%)、软壁菌门(4.99%)。在对照组虾酱发酵过程中,相对丰度排在前3 位的菌门依次为变形菌门、拟杆菌门和软壁菌门,发酵结束时软壁菌门的相对丰度出现了下降。在加酶组中,相对丰度排在前3 位的菌门依次为变形菌门、拟杆菌门和厚壁菌门;在发酵过程中,拟杆菌门减少,而厚壁菌门增加。由此可见,加入中性蛋白酶制剂后,虾酱中拟杆菌门的相对丰度减少而厚壁菌门增加。变形菌门是革兰氏阴性菌的主要类群之一,在人体[19]和虾类[20]中广泛存在。对渤海等地区的养殖池塘中的水体研究也发现变形菌门为优势类群[21-23]。本研究中虾酱的优势菌门是变形菌门,很有可能与原料虾有关。变形菌门是细菌中最大的一门,包括很多病原菌,如大肠杆菌、沙门氏菌、幽门螺杆菌等[24]。因此,变形菌门对虾酱的品质影响极大,在虾酱的发酵生产和贮存过程中需严格控制变形菌门的生长。
2.3.3 虾酱细菌菌群科水平分布

图5 虾酱细菌组成在科水平上的分布Fig.5 Bacterial community composition in shrimp paste at the family level
在虾酱中共检出了305 个菌科,如图5所示,主要为红细菌科(Rhodobacteraceae)、莫拉菌科(Moraxellaceae)、黄杆菌科(Flavobacteriaceae)、杆状菌科(Bacillaceae)、虫原体科(Entomoplasmatales-Incertae-Sedis)、伯克氏菌科(Burkholderiaceae)、全孢菌科(Holosporaceae)、脱硫弧菌科(Desulfovibrionaceae)、拟南芥科(Arcobacteraceae)、希瓦氏菌科(Shewanellaceae)和弧菌科(Vibrionaceae)。在发酵初时(C0),相对丰度由高到低为黄杆菌科(18.75%)、弧菌科(13.09%)、希瓦氏菌科(7.07%)、拟南芥科(6.61%)、莫拉菌科(4.85%)。在对照组发酵过程中,前3 位的优势科是红细菌科、黄杆菌科和脱硫弧菌科。与戴玲瑛等[13]研究所得的连云港虾酱优势菌科为弧菌科有差异。在整个发酵过程中,红细菌科相对丰度增加,而黄杆菌科和脱硫弧菌科呈现波动变化。在加酶组中,前3 位的优势科是红细菌科、莫拉菌科和杆状菌科;在整个发酵过程中,红细菌科和杆状菌科均有所增加,而莫拉菌科呈现减少趋势。因此,加入蛋白酶制剂后,虾酱中莫拉菌科、杆状菌科和全孢菌科增加,而黄杆菌科和脱硫弧菌科减少。
2.3.4 虾酱细菌菌群属水平分布
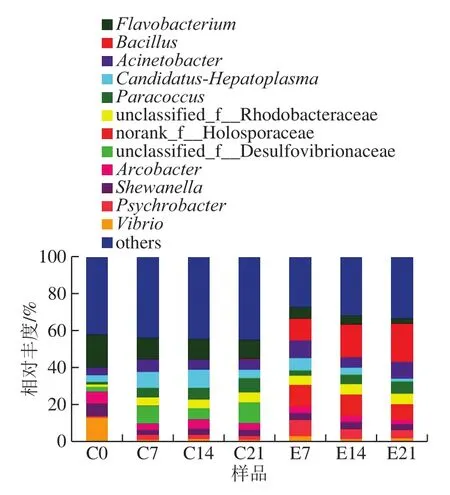
图6 虾酱细菌组成在属水平上的分布Fig.6 Bacterial community composition in shrimp paste at the genus level
本研究中共得到了642 个菌属,如图6所示,主要为黄杆菌属(Flavobacterium)、芽孢杆菌属、不动杆菌属(Acinetobacter)、Candidatus-Hepatoplasma、副球菌属(Paracoccus)、弓形杆菌属(Arcobacter)、希瓦氏菌属(Shewanella)、嗜冷杆菌属(Psychrobacter)和弧菌属(Vibrio)。在虾酱发酵的初始阶段(C0),相对丰度由高到低为黄杆菌属(17.78%)、弧菌属(13.05%)、希瓦氏菌属(7.07%)。这与尹超[15]研究结果中发酵初始阶段优势菌属为中慢生根瘤菌属(Mesorhizobium)、苯基杆菌属(Phenylobacterium)、慢生根瘤菌属(Bradyrhizobium)和盐渍微菌属(Salimicrobium)有所不同,这可能是因为原料产地和发酵方式的不同。在对照组发酵过程中,前3 位的优势菌属为黄杆菌属、Candidatus-Hepatoplasma和副球菌属。Jung等[25]发现,在15℃条件下,含25 g/100 mL食盐发酵的虾制品中优势菌属为盐厌氧菌属(Halanaerobium)。Lee等[26]证明,在15 ℃发酵时,盐沼虾中最终加盐量为20 g/100 mL和24 g/100 mL的虾酱中优势菌属均为盐厌氧菌属,而在28 g/100 mL加盐量的样本中,盐渍微菌最终成为占主导地位的菌属。Lee等[27]研究显示,在25 g/100 mL盐渍虾样品中,盐渍微菌在10 ℃发酵时未出现,但在20 ℃和25 ℃发酵时为优势菌属。本结果与之前研究的虾制品中优势菌属存在差异,这说明原料及制作方式会对虾制品微生物区系产生影响[28-29]。黄杆菌属在发酵过程中出现缓慢下降,由最初的17.78%减少至10.45%,但是其在整个过程中都被检测到,说明它可能是发酵过程中主要的微生物,但在泰国传统发酵虾酱中并未检测出[6]。芽孢杆菌属由最初的0.06%缓慢减少至0.01%,不动杆菌属也出现了轻微增长,Candidatus-Hepatoplasma在发酵过程中相对丰度增加。在加酶组发酵过程中,排名前3的优势菌属为芽孢杆菌属、不动杆菌属和嗜冷杆菌属。在整个发酵过程中,芽孢杆菌属相对丰度增加。在加入蛋白酶制剂后黄杆菌属相对丰度降低,至发酵末期,所占比例为2.75%,与对照组明显不同的是,芽孢杆菌属含量显著增加,在发酵末期所占比例为20.69%。芽孢杆菌属为产乳酸菌属,所以在整个发酵中加酶组的pH值低于对照组。不动杆菌属较对照组也有所增加,而Candidatus-Hepatoplasma在发酵结束时相对丰度降低至1.6%。弧菌属在虾肠道中普遍存在[30-31],对凡纳滨对虾肠道和三疣梭子蟹肠道进行多样性分析后也可得出相同的结论[32-33]。在整个发酵过程中四联球菌属相对丰度较低,四联球菌属在高渗发酵食品中较为常见[34],但在本研究结果中丰度较低,这在Kobayashi等[35]研究中已有报道。
通过对样品进行主成分分析(principal component analysis,PCA)可以看出,加酶组和对照组均分别聚集,表现出与C0样品明显的组成成分差异,表明对照组中虾酱样品细菌组成的相似性。其余样品中E7、E14、E21形成聚集,表明加酶组样品中类似的细菌菌落结构。图7表明,对照组和加酶组样品之间存在显著差异(P<0.05),说明加入蛋白酶制剂后对虾酱的微生物区系影响显著。

图7 虾酱发酵不同时期的PCAFig.7 Principal composition analysis plot for discrimination of shrimp paste samples with different fermentation times
2.3.5 不同处理虾酱差异菌群分析
使用线性判别分析(LEfSe)找出对组间差异存在显著影响的微生物类群,如图8所示,拟杆菌门、拟杆菌纲(Bacteroidia)、黄杆菌目(Flavobacteriales)、黄杆菌属、Deltaproteobacteria、脱硫弧菌目(Desulfovibrionales)等在对照组中相对丰度显著高。杆状菌科、芽孢杆菌属、厚壁菌门、Holosporales、Alfhaproteobacteria等在加酶组中相对丰度显著高,对菌群组间差异存在显著影响。值得注意的是,在加酶组虾酱中检测到高丰度的芽孢杆菌属,这可能引起加酶组虾酱品质下降。

图8 虾酱微生物群落结果直方图(阈值为4)Fig.8 Histogram of the microbiota of shrimp paste with a threshold value of 4

图9 虾酱在属水平组间差异菌群图Fig.9 Differential bacterial genera between the two types of shrimp paste
图9 是在属水平上探索影响较大的差异菌群,其中芽孢杆菌属、乳球菌属在组间差异为显著。证明加入蛋白酶制剂后,芽孢杆菌属显著增加,而乳球菌属显著减少。芽孢杆菌属在海洋环境中大量分布[36],通常情况下,添加淀粉质原料于发酵性鱼虾产品中,会提高芽孢杆菌属的相对丰度,本研究中,添加商品化中性蛋白酶后芽孢杆菌属相对丰度增加,成为优势细菌,与前者结果一致。由于两种处理下的虾酱原料相同,故排除原料对微生物区系的影响后可以得出,添加商品化酶制剂会显著增加芽孢杆菌属的含量[37]。此外,本研究添加酶制剂后希瓦氏菌属相对丰度增加,希瓦氏菌属往往会引起水产品腐败及组胺的积累,所以在安全方面对于加酶虾酱应更为重视。
2.4 细菌菌群与理化性质之间的相关性
2.4.1 主要优势菌群与理化性质的相关性分析
为了对细菌菌群与理化性质进行相关性分析,通过Spearman分析构建了理化性质与主要菌属之间的相关性热图。由图10可知,与弓形杆菌属有较高相关性的为TVB-N值,该菌属与TVB-N值呈显著负相关(R2=-0.79),与丙二醛含量呈正相关(R2=0.75)。芽孢杆菌属与TVB-N值呈正相关(R2=0.64),这说明芽孢杆菌属可能是导致加酶组虾酱品质下降的原因。黄杆菌属与丙二醛含量呈正相关(R2=0.96),与TVB-N值和氨基酸态氮含量呈负相关,对于黄杆菌属在虾酱发酵中的作用还尚未可知,需要以后多加研究。副球菌属和pH值、氨基酸态氮含量和色胺含量呈正相关。乳球菌属与TVB-N值呈负相关(R2=-0.82),在之后研究中可以考虑通过添加乳球菌属提高虾酱样品的品质。据目前已有的报道,乳酸菌是产生生物胺的主要微生物,其中肠球菌、乳杆菌、乳球菌、明串珠菌、片球菌和链球菌是生物胺的强力生产者,然而,本研究结果中生物胺含量与乳球菌属并没有呈现高度相关性,它也证实了生物胺的形成高度依赖于细菌菌株而不是微生物的种类[38]。

图10 细菌属与理化性质的相关分析Fig.10 Correlation analysis between bacterial genera and physicochemical properties
2.4.2 虾酱样品理化性质与主要物种的冗余分析(redundancy analysis,RDA)
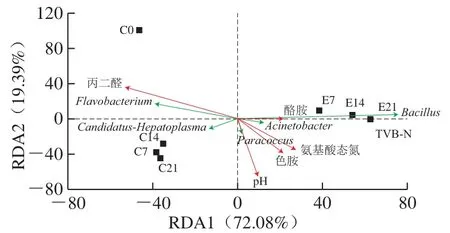
图11 虾酱样品理化性质与细菌属之间的RDAFig.11 RDA analysis between physicochemical properties and bacterial genera of shrimp paste samples
RDA使用物种水平的OTU数据显示了不同处理虾酱样品中微生物群落的差异。由图11可以看出,添加酶制剂使虾酱样品在微生物群落中出现明显差异。从RDA可以看出3 类微生物群落结构,第1组是主要由黄杆菌属组成的初始发酵虾酱样品C0,第2组是主要由Candidatus-Hepatoplasma和黄杆菌属组成的对照组(C7、C14和C21),第3组是主要由芽孢杆菌属组成的加酶组(E7、E14和E21)。同时,结果显示添加酶制剂后导致TVB-N值高于对照组,这可能造成虾酱的品质下降。因此,对于外加蛋白酶制剂所制得的虾酱,应该对其质量进行更为全面的评价,以更好地保证虾酱的质量与安全。
3 结 论
本研究利用高通量测序技术分析外加酶制剂对于虾酱微生物区系的影响,对虾酱进行微生物多样性分析后共得到29 个门、305 个科、642 个属。在门水平上,所有样品中变形菌门为优势菌门,对照组中拟杆菌门丰度较高,而加酶组中厚壁菌门丰度较高;在科水平上,对照组中黄杆菌科为优势菌科(18.75%),加酶组中杆状菌科为优势菌科(20.69%);在属水平上,对照组中黄杆菌属为优势菌属,而加酶组中芽孢杆菌属为优势菌属。通过LEfSe分析得到外加酶制剂使得芽孢杆菌属和乳球菌属在两种处理虾酱之间产生显著差异,通过理化性质可以得出,添加中性蛋白酶可能使虾酱的品质有所下降,但对于生物胺并无显著影响。所以,在以后的工业生产中,要对不同处理得到的虾酱进行质量的严格控制,可以考虑从原料新鲜度、发酵温度及添加剂进行改进,将生产标准化、规范化以降低风险。这对于保证虾酱的安全与质量具有重要意义。
猜你喜欢
河南医学研究(2022年19期)2022-10-19 00:44:18
中国比较医学杂志(2020年4期)2020-05-26 05:47:22
水生生物学报(2019年4期)2019-07-20 08:08:10
生态学报(2019年11期)2019-07-08 06:18:58
生物安全学报(2019年3期)2019-02-15 16:54:12
川北医学院学报(2019年6期)2019-02-10 10:48:32
美食(2018年11期)2018-12-05 11:29:54
农村百事通(2018年6期)2018-03-29 09:02:14
意林(2014年6期)2014-05-05 15:24:40
食品工业科技(2014年23期)2014-03-11 18:19:08
