
白藜芦醇通过调控自噬减轻乳大鼠心肌细胞低氧复氧损伤
2020-12-09 11:26张金芳刘梦思孙秀玉刘立亚李志勇黄秀兰
中国药理学与毒理学杂志 2020年8期
张金芳,刘梦思,孙秀玉,刘立亚,李志勇,黄秀兰
(中央民族大学1.药学院,2.生命与环境科学学院,北京 100081)
据《中国心血管病报告2018》报道,冠心病患病率和死亡率持续升高,在心脑血管疾病中仅次于卒中[1]。心肌缺血再灌注损伤(myocardial ischemia reperfusion injury,MIRI)是由冠状动脉完全或局部闭塞后,血液供需失衡,在一定时间内恢复灌注致使心肌损伤加重的病理过程[2]。目前,自噬在MIRI的发生发展过程中越来越受到关注,其调控机制可能与自噬的状态有关[3]。自噬包括自噬体形成、自噬体与溶酶体融合和自噬溶酶体降解3个阶段[4]。目前认为心肌缺血时自噬可减少心肌细胞死亡,对心肌具有保护作用;再灌注时,自噬加重心肌细胞损伤,其机制是自噬过度还是自噬流受阻则存在着争议[3,5-6]。另有研究发现,小鼠MIRI可激活细胞自噬,不同缺血时间和不同再灌注时间导致自噬的状态不同[7]。可见,MIRI过程中自噬的改变值得深入研究。
白藜芦醇(resveratrol,Res)是一种存在于花生、葡萄和中药虎杖等植物中的多酚化合物,具有抗氧化、抗炎和抗肿瘤等药理活性,主要用于防治心血管疾病、糖尿病、高血压和肾疾病等[8]。研究表明,Res可通过调控自噬减轻MIRI,目前尚不清楚其机制是否与自噬的改变有关[9]。本研究通过体外培养乳大鼠心肌细胞,制备心肌细胞低氧复氧(hypoxia/reoxygenation,H/R)损伤模型同时加用自噬抑制剂氯喹(chloroquine,CQ),以探讨Res对H/R损伤心肌细胞的保护作用和机制。
1 材料与方法
1.1 药品、试剂和主要仪器
Res(纯度>99%,长沙康隆生物制品有限公司赠予);CQ和5-BrdU(美国Sigma公司);青、链霉素和DMEM/F12培养基(美国Corning公司);胎牛血清(美国Gibco公司);兔抗大鼠P62多克隆抗体(美国Abcam公司);兔抗大鼠β肌动蛋白单克隆抗体和兔抗大鼠Beclin 1单抗(美国CST公司);兔抗大鼠微管相关蛋白1轻链3(microtubule associated protein light chain 3,LC3)多抗(美国Invitrogen公司);FITC标记的山羊抗兔IgG抗体(美国Millipore公司);乳酸脱氢酶(lactate dehydrogenase,LDH)试剂盒和CCK-8试剂盒(日本株式会社东仁化学研究所);AnnexinⅤ-FITC凋亡检测试剂盒(美国BD公司)。
三气培养箱(美国Thermo公司);CO2细胞培养箱(德国Binder公司);倒置相差显微镜和倒置荧光显微镜(日本Olympus株式会社);垂直板电泳槽(北京六一生物技术有限公司);湿式转移电泳槽(美国Bio-Rad公司);化学发光成像仪(上海天能科技有限公司);多功能酶标仪和流式细胞仪(美国Beckman Coulter公司)。
1.2 动物和原代心肌细胞制备培养
出生24 h内SPF级SD乳大鼠,实验动物使用许可证:SCXK-(军)0007-004(军事医学科学院实验动物中心)。按文献[10]分离乳大鼠心肌细胞,接种于含10%胎牛血清和1%双抗(青霉素和链霉素)的DMEM/F12培养基中,置CO2细胞培养箱(37℃,5% CO2)中培养。待细胞融合度约80%时,用于实验。
1.3 心肌细胞H/R损伤模型制备
心肌细胞常规用含10%胎牛血清的DMEM/F12的培养基培养。低氧处理时,换用三气培养箱中预饱合1 h的Hank液,置于三气培养箱(94% N2,5% CO2,1% O2)中培养12 h,复氧处理时,换成常规培养液,置CO2培养箱中培养12 h。
1.4 细胞分组和处理
乳大鼠心肌细胞分为5组:细胞对照组,常规培养基培养12 h后,换新鲜培养液继续培养24 h;H/R模型组,常规培养基培养12 h,然后低氧12 h,复氧12 h;模型+Res 5 和 20 μmol·L-1组,Res 5 和20 μmol·L-1预处理细胞12 h,然后低氧12 h,复氧12 h;模型+Res 20 μmol·L-1+CQ 10 μmol·L-1组,Res 20 μmol·L-1预处理细胞12 h,然后换含终浓度为CQ 10 μmol·L-1的 Hank液低氧培养12 h,再换用终浓度为CQ 10 μmol·L-1复氧12 h。
1.5 CCK-8法测定心肌细胞存活率
将心肌细胞5×108L-1接种于96孔板,每孔100 μL,每组设6复孔,按1.5分组处理细胞H/R结束后,加入CCK-8溶液置于CO2培养箱中孵育4 h,用酶标仪在450 nm处检测吸光度值(A450nm),计算细胞存活率。细胞存活率(%)=(给药组A450nm-空白组A450nm)/(细胞对照组A450nm-空白组A450nm)×100%。
1.6 比色法测定心肌细胞LDH漏出率
密度为5×108L-1心肌细胞接种于96孔板,每孔100 μL,每组设6复孔,按1.5分组处理细胞。H/R结束后,按LDH试剂盒说明书操作,同时设不经药物处理的高对照组(含培养基、细胞悬液和裂解缓冲液)和低对照组(含培养基和细胞悬液)。用酶标仪测定490 nm处的吸光度值(A490nm),计算LDH漏出率。LDH漏出率(%)=(样品A490nm-低对照A490nm)/(高对照A490nm-低对照A490nm)×100%。
1.7 AnnexinⅤ/Pl双染法检测细胞凋亡
按照1.5分组处理细胞,H/R结束后,胰蛋白酶消化法收集细胞并调整密度,取细胞悬液100 μL,按AnnexinⅤ-FITC凋亡检测试剂盒说明书操作,加AnnexinⅤ-FITC染色液5 μL避光孵育15 min,再加PI染色液5 μL,用结合缓冲液终止反应,1 h内用流式细胞仪检测分析。
1.8 免疫荧光法检测心肌细胞LC3阳性细胞率
将密度为5×108L-1心肌细胞接种于激光共聚焦小皿,分别按1.5分组处理细胞,H/R结束后,4%多聚甲醛固定,0.2%Triton X-100室温透化,5%BSA-PBS封闭2 h。兔抗大鼠LC3多抗(稀释倍数为1∶200)4℃孵育过夜。FITC标记的羊抗兔IgG抗体(二抗,稀释倍数为1∶500)于CO2培养箱中湿盒避光孵育1 h,室温下DAPI避光孵育10 min,随机选取5个高倍(400×)视野,计数心肌细胞总数和LC3阳性细胞数(细胞至少有5个LC3斑点),计算LC3阳性细胞率。LC3阳性细胞率(%)=LC3阳性细胞数/细胞总数×100%。
1.9 Western印迹法检测心肌细胞Beclin 1,LC3Ⅱ和P62蛋白表达水平
分别按1.5分组处理细胞,H/R结束后,加含蛋白酶抑制剂cocktail的RIPA的裂解液,冰上裂解30 min后,9120×g,4℃离心20 min,提取细胞总蛋白。BCA法测定总蛋白浓度,SDS-PAGE凝胶电泳分离蛋白。5%脱脂奶粉封闭2 h,一抗(抗Beclin 1、LC3和β肌动蛋白1∶1000稀释,抗P62为1∶500稀释)4℃孵育过夜,二抗(1∶10 000稀释)室温孵育1 h,加ECL发光液避光反应,用显影仪拍照。采用Image J软件分析条带积分吸光度值,以目标蛋白与内参蛋白条带积分吸光度比值表示蛋白的相对表达水平。
1.10 统计学分析
用SPSS 20.0软件进行统计分析,实验结果数据以±s表示,实验重复3次。组间比较使用单因素方差分析(one-way ANOVA)进行数据统计,方差齐时采用LSD(L)检验,方差不齐时采用TamhaneT2(M)检验,以P<0.05为差异具有统计学意义。
2 结果
2.1 Res对H/R损伤乳大鼠心肌细胞形态的影响
倒置相差显微镜观察心肌细胞形态,细胞对照组心肌细胞融合度较高,细胞多呈梭形、三角形或星形,轮廓清晰且折光性强(图1A)。H/R组细胞融合度明显降低,细胞间隙较大,折光度低(图1B)。H/R+Res 5 和20 μmol·L-1组心肌细胞生长状态明显改善,融合度升高,折光度增强(图1C、图1D)。H/R+Res 20 μmol·L-1加用 CQ 10 μmol·L-1后,细胞间隙增大,状态与H/R组相似(图1E)。

Fig.1 Effect of resveratrol(Res)on morphology of myocardial cells of newborn rats after hypoxia/reoxygenation(H/R).Model+Res 5 and 20 μmol·L-1group:cardiomyocytes were pretreated with Res 5 and 20 μmol·L-1for 12 h,and then processed with hypoxia for 12 h,reoxygenation for 12 h.Model+Res 20 μmol·L-1+CQ 10 μmol·L-1group:after cardiomyocytes were pretreated with Res 20 μmol·L-1for 12 h,processed with Hank medium containing the final concentration of CQ 10 μmol·L-1 to H for 12 h,and then replaced conventional DMEM/F12 culture medium containing the final concentration of CQ 10 μmol·L-1to reoxygenation for 12 h.
2.2 Res对H/R损伤乳大鼠心肌细胞存活率和LDH漏出率的影响
与细胞对照组相比,H/R组心肌细胞存活率显著下降(P<0.01),LDH漏出率显著升高(P<0.01);与H/R组比较,H/R+Res 5 和20 μmol·L-1组心肌细胞存活率显著升高(P<0.05),LDH漏出率显著下降(P<0.01)(表1)。与H/R+Res 20 μmol·L-1组比较,H/R+Res 20 μmol·L-1+CQ 10 μmol·L-1组的细胞存活率明显降低(P<0.05),LDH漏出率显著升高(P<0.01),接近H/R组水平。

Tab.1 Effect of Res on cell viability and leakage rate of lactate dehydrogenase(LDH)of myocardial cells of newborn rats after H/R
2.3 Res对H/R损伤乳大鼠心肌细胞凋亡率的影响

Fig.2 Effect of Res on myocardial cells apoptosis of newborn rats after H/R.See Fig.1 for the cell treatment.±s,n=3.**P<0.01,compared with cell control group; #P<0.05,##P<0.01,compared with H/R group;ΔP<0.05,compared with H/R+Res 20 group.
与细胞对照组相比,H/R组细胞早期凋亡率和总凋亡率均显著升高(P<0.01)(图2)。与H/R组相比,H/R+Res 5和20 μmol·L-1组细胞早期凋亡率和总凋亡率均显著降低(P<0.01,P<0.05)。与H/R+Res 20 μmol·L-1组比较,H/R+Res 20 μmol·L-1+CQ 10 μmol·L-1组细胞早期凋亡率明显上升(P<0.05),总凋亡率无明显变化。
2.4 Res对H/R损伤乳大鼠心肌细胞内LC3阳性细胞率的影响
与细胞对照组相比,H/R组心肌细胞的LC3荧光点聚集不明显,LC3阳性细胞率无明显变化(图3)。与H/R组相比,H/R+Res 5和20 μmol·L-1组心肌细胞的LC3荧光点聚集较大、较亮,LC3阳性细胞率显著增加(P<0.05);H/R+Res 20 μmol·L-1+CQ 10 μmol·L-1组LC3荧光点聚明显多且亮,LC3阳性细胞率显著增加(P<0.01)。与H/R+Res 20 μmol·L-1组比较,H/R+Res 20 μmol·L-1+CQ 10 μmol·L-1组的LC3荧光点聚集更大、更亮;LC3阳性细胞率无明显变化。
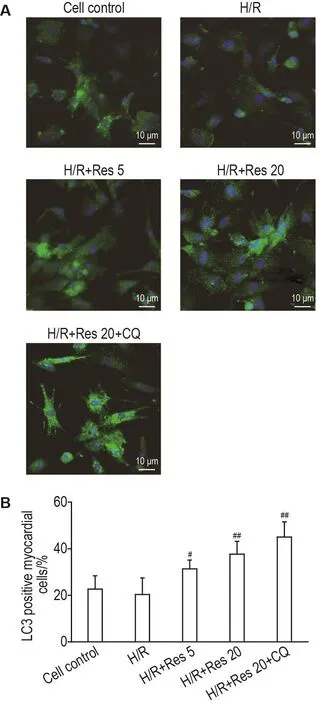
Fig.3 Effect of Res on C3 positive myocardial cells rate in myocardial cells of newborn rats after H/R by immunofluorescence.See Fig.1 for the cell treatment.±s,n=3.#P<0.05,##P<0.01,compared with H/R group.
2.5 Res对H/R损伤乳大鼠心肌细胞Beclin 1,LC3Ⅱ和P62蛋白表达水平的影响
如图4所示,与细胞对照组相比,H/R组Beclin 1蛋白表达水平无明显变化;LC3Ⅱ和P62表达蛋白水平显著下调(P<0.05)。与H/R组相比,H/R+Res 5和20 μmol·L-1组Beclin 1,LC3Ⅱ和P62蛋白表达水平均显著上调(P<0.05);H/R+Res 20 μmol·L-1+CQ 10 μmol·L-1组Beclin 1 无明显变化,LC3Ⅱ和P62蛋白表达水平均显著上调(P<0.01)。与 H/R+Res 20 μmol·L-1组比较,H/R+Res 20 μmol·L-1+CQ 10 μmol·L-1组Beclin 1蛋白表达水平下调,LC3Ⅱ和P62蛋白表达水平无明显变化。
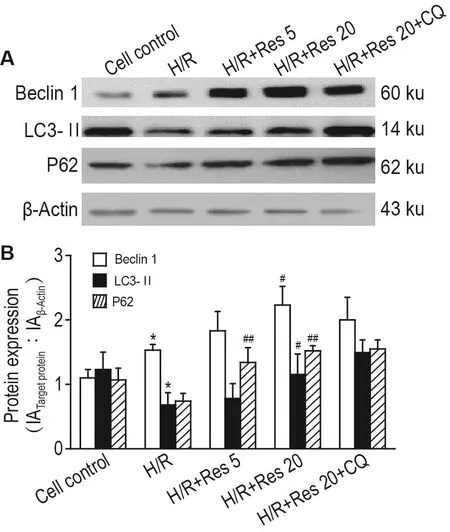
Fig.4 Effect of Res on protein expressions of Beclin 1,LC3Ⅱand P62 in myocardial cells of newborn rats after H/R by Western blotting.See Fig.1 for the cell treatment.IA:integrated absorbance.±s,n=3.*P<0.05,compared with cell control group;#P<0.05,##P<0.01,compared with H/R group.
3 讨论
本研究结果表明,经Res预处理后,H/R损伤心肌细胞的细胞活力升高,LDH漏出率下降,凋亡率降低,而加用CQ后,上述结果呈相反现象,提示Res对H/R损伤的心肌细胞具有保护作用,并推测Res对心肌细胞的保护作用可能与自噬有关。
自噬在MIRI中发挥重要作用,MIRI的机制可能与自噬的改变有关[11]。自噬是细胞清除受损或老化的细胞器和错误折叠的蛋白质,以维持细胞内稳态的适应性机制,其动态过程又称为自噬流[4]。Beclin 1是自噬启动的标志蛋白,自噬标志蛋白LC3存在于自噬的整个过程中,P62表达降低可能预示着自噬溶酶体的降解顺畅,因此Beclin 1,LC3和P62是目前公认的反映自噬的重要蛋白[12]。自噬被激活后,胞浆内的LC3Ⅰ脂化成为位于自噬体膜上的LC3Ⅱ,LC3Ⅱ和自噬小泡结合形成自噬体[13]。自噬底物蛋白P62和LC3Ⅱ相互作用形成自噬溶酶体并在自噬后期降解,而缺少Atg基因或自噬体与溶酶体的融合过程受到阻碍,均会使P62堆积,因此通过检测P62的表达量来监测自噬的变化[13-14]。CQ可升高溶酶体内pH,降低溶酶体酶活性,不利于自噬体与溶酶体融合,阻碍自噬溶酶体的形成,从而抑制自噬,常用作评价自噬的工具药[15]。由于LC3Ⅱ水平升高存在2种情况,即可能是自噬被激活引起自噬体增多也可能是自噬体与溶酶体融合受阻或自噬溶酶体降解受阻,因此需要加用CQ,进一步明确细胞内自噬的状态[16]。研究发现,单纯低氧和不同时间H/R的大鼠心肌细胞LC3Ⅱ/LC3Ⅰ水平均显著提高,单纯低氧能激活自噬,H/R则进一步激活自噬,且H/R会加重低氧引起的细胞损伤[17]。本研究结果发现,心肌细胞H/R损伤后引起Beclin 1表达升高,LC3Ⅱ和P62表达下降,表明H/R可能通过抑制自噬,诱发心肌细胞损伤。
Res的抗MIRI作用可能与调控细胞自噬有关。研究证实,Res可通过升高LC3Ⅱ蛋白表达、ATP含量和自噬小泡,以及抑制西罗莫司(雷帕霉素)和p70S6激酶,激活心肌细胞自噬,抑制心梗面积扩大,改善心功能,减轻小鼠MIRI[18]。研究发现,Res后处理能通过增加自噬体形成、提高LC3Ⅱ和Beclin 1的水平降低H/R诱导的H9c2心肌细胞损伤[19]。本研究结果显示,Res预处理可激活H/R损伤的心肌细胞内Beclin 1蛋白,LC3Ⅱ和P62表达水平明显上调,LC3阳性细胞率增加,提示Res可活化自噬;而加用CQ后LC3Ⅱ和P62的表达水平和LC3阳性细胞率均进一步增加,提示可逆转Res的心肌保护作用。以上结果提示,Res对H/R损伤的心肌细胞的保护作用机制可能是通过活化自噬发挥的。
综上所述,本研究采用乳大鼠心肌细胞H/R损伤模拟MIRI模型,发现Res可通过活化自噬以降低H/R诱导的心肌细胞LDH漏出率和凋亡率,减轻心肌细胞损伤,表明Res对H/R造成的心肌细胞损伤具有改善作用,为Res在MIRI的潜在临床应用提供实验依据。
猜你喜欢
中国现代医生(2022年23期)2022-09-21
河南科技(2022年6期)2022-04-22
现代仪器与医疗(2021年6期)2022-01-18
口腔颌面外科杂志(2021年3期)2021-07-15
中国比较医学杂志(2020年11期)2020-12-09
数码世界(2020年4期)2020-06-18
野生动物学报(2020年1期)2020-02-21
食品工程(2020年3期)2020-01-05
读与写(2019年35期)2019-11-05
现代职业教育·高职高专(2018年7期)2018-05-14
