
低温胁迫下4个景天科多肉品种的生理响应及耐寒性评价
2020-02-21 08:41陈艺荃林榕燕潘宏林兵叶秀仙
福建农业学报 2020年10期
陈艺荃 林榕燕 潘宏 林兵 叶秀仙
摘要:【目的】研究低温胁迫下景天科多肉植物的生理响应,评价不同品种的耐寒能力,筛选出耐寒性强的多肉品种,以繁育推广。【方法】以观赏性强的‘蓝苹果:‘火祭、‘丸叶松绿和‘筒叶花月等4个多肉品种为试材,通过人工低温(9、6、3、0和-3℃)胁迫处理后测定8项生理指标,将主成分分析与隶属函数法相结合,构建数学模型进行耐寒性综合评价,并通过恢复生长试验对评价结果进行验证。【结果】4个多肉品种应对低温胁迫的生理变化有显著差异,MDA和Pro含量变化呈上升趋势,而可溶性糖、相对含水量和光合色素含量变化因品种而异。-3℃时,‘火祭相对含水量增幅最大且MDA释放量最大,‘筒叶花月的光合色素含量最稳定,‘丸叶松绿的可溶性糖与Pro积累最迅速。8项生理指标可综合为2个主成分,累积贡献率达90.5 g%,生物学意义显著。耐寒性综合评价得出4个多肉品种耐寒能力大小为‘丸叶松绿>‘蓝苹果>‘筒叶花月>‘火祭:评价结果与恢复生长试验观测结果基本一致。【结论】‘丸叶松绿在-3℃低温胁迫下恢复室温生长成活率达85.5%,植株长势良好,形态性状稳定,可繁育推广;而‘火祭成活率仅为25 .5%,对低温适应性有限。
关键词:低温胁迫;生理指标;耐寒性;综合评价
中图分类号:S 682.33
文献标志码:A
文章编号:1008-0384( 2020) 10-1093-08
0 引言
【研究意义】多肉植物( Succulents)原产于北美洲和非洲干旱、半干旱地带,约50科334属,茎叶肥厚,耐旱、耐盐碱。景天科多肉植物( Crassulaceaeplants)外形秀美,叶色绮丽变化多端,花期和绿期较长,观赏性强,具食用、药用、生态和景观应用等多重价值,是最受市场欢迎的多肉种类之一。福建省是我国多肉四大产区之一,但商品化栽培起步较晚[1],产业基础尚未成熟。景天科植物适宜的生长温度为15~28℃,福建冬季与多肉原产地气候差异显著,最低气温低于5℃,部分山区在0℃以下,低于多肉最适生长温度的下限,因此亟需开展多肉耐寒性研究,筛选耐寒优种,降低引种种植的经济损失,對科学引种和繁育推广具有重要意义。【前人研究进展】景天科植物原产于热带、亚热带地区,不耐寒,在低温下易受冷害或冻害,引起膜结构改变、胞质环流缓慢、光合速率降低等生理代谢紊乱,严重影响生长发育[2-3],因此种质资源耐寒性鉴定一直是多肉植物抗逆性的重要研究领域[4]。前人在景天科植物分类[5]、观赏性评价[6]、繁育栽培技术[7-9]、活性物质提取[10-11]、光合特性和耐旱性[12-14]等方面已有较多报道。景天科植物耐寒性研究较多集中在景天属(SedumL.)、长生草属(Sempervivum)地被植物。研究表明,‘黄针叶万年草(Sedum rupestreAngelina)、‘六棱景天(Sedum sexangulare)的耐寒性强于‘胭脂红景天(Sedum spurium);‘观音莲(Semper-vivum tectorum)在-8℃低温胁迫后,依然能够恢复正常生长[15-16],说明不同多肉品种耐寒性存在显著差异。在低温胁迫生理响应方面,研究认为总叶绿素含量、MDA含量、游离脯氨酸(Pro)等生理指标的变化趋势和幅度在一定程度上能够反映多肉抵御低温的能力[17]。【本研究切人点】景天科多肉植物作为新兴的园林景观花卉,品种繁多,品种间耐寒性差异大,仍有大量观赏性强的多肉种质尚未进行低温胁迫生理响应研究和耐寒性鉴定与评价。【拟解决的关键问题】本研究在对景天科多肉植物观测、评价、筛选的基础上,以观赏性强、商品价值高的4个多肉品种为对象,分析其低温胁迫下的生理生化代谢差异,结合主成分分析、隶属函数分析法对其耐寒性进行评定,并通过恢复生长试验来验证评价结果,筛选耐寒性较强的品种,为多肉种质资源推广利用提供参考。
1 材料与方法
1.1 供试材料
供试材料为‘蓝苹果(x Sedeveria Blue Elf)、‘火祭(Crassula Campfire)、‘丸叶松绿(Sedumlucidum Obesum)和‘筒叶花月(Crassula obliqueGollum)4个景天科多肉品种,均引种自云南省昆明市。选取健康、长势一致的3年生盆栽植株,种植条件与栽培管理措施一致。
1.2 试验方法
试验在福建省农业科学院作物研究所花卉种苗生理与检测实验室进行。将4个品种盆栽植株置于人工气候培养箱(普朗特LRX-700C-LED)中预处理5d,昼夜温度150C/12℃,光照强度12 000 lx,光周期昼夜12 h/12 h,相对湿度45%。预处理结束后进行低温胁迫处理,光照强度、光周期和湿度同预处理,处理温度梯度为9、6、3、0、-3℃,其中9℃为对照(CK),每级温度处理12 h,各处理30盆,每盆4~6株,其中10盆用于形态观察。将经过不同低温处理后的试材立即测定生理指标,设3次重复。观测低温胁迫后植株恢复室温的生长状况和形态变化,统计成活率。
1.3 指标测定
参考《植物生理生化实验原理与技术》中的方法进行测定[18]。取低温胁迫处理后各品种第三轮数的叶片测定相对含水量( RWC)、丙二醛(MDA)、叶绿素a( Chla)、叶绿素b(Chlb)、总叶绿素(TC)、类胡萝卜素( Car)、可溶性糖(SS)和游离脯氨酸( Pro)等生理指标。按照饱和含水量法测定叶片相对含水量,丙酮浸提法(丙酮:乙醇=2:1)测定叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量,蒽酮比色法测定可溶性糖含量,硫代巴比妥酸( TBA)法测定丙二醛含量,酸性茚三酮比色法测定游离脯氨酸含量。叶绿素、可溶性糖、丙二醛、Pro等指标的含量均为鲜重的含量。
1.4 数据处理
用Excel 2016对原始数据进行统计整理和制图。用SPSS21.0軟件对所有指标进行方差分析、隶属函数计算和主成分分析[19-20],用Duncan新复极差法对各处理间差异进行显著性分析。
1.4.1 隶属函数分析 采用模糊数学中的隶属函数法,计算8个生理指标的隶属函数,所有数据标准化后再进行分析,计算公式为:
2 结果与分析
2.1低温胁迫下多肉植物的生理响应
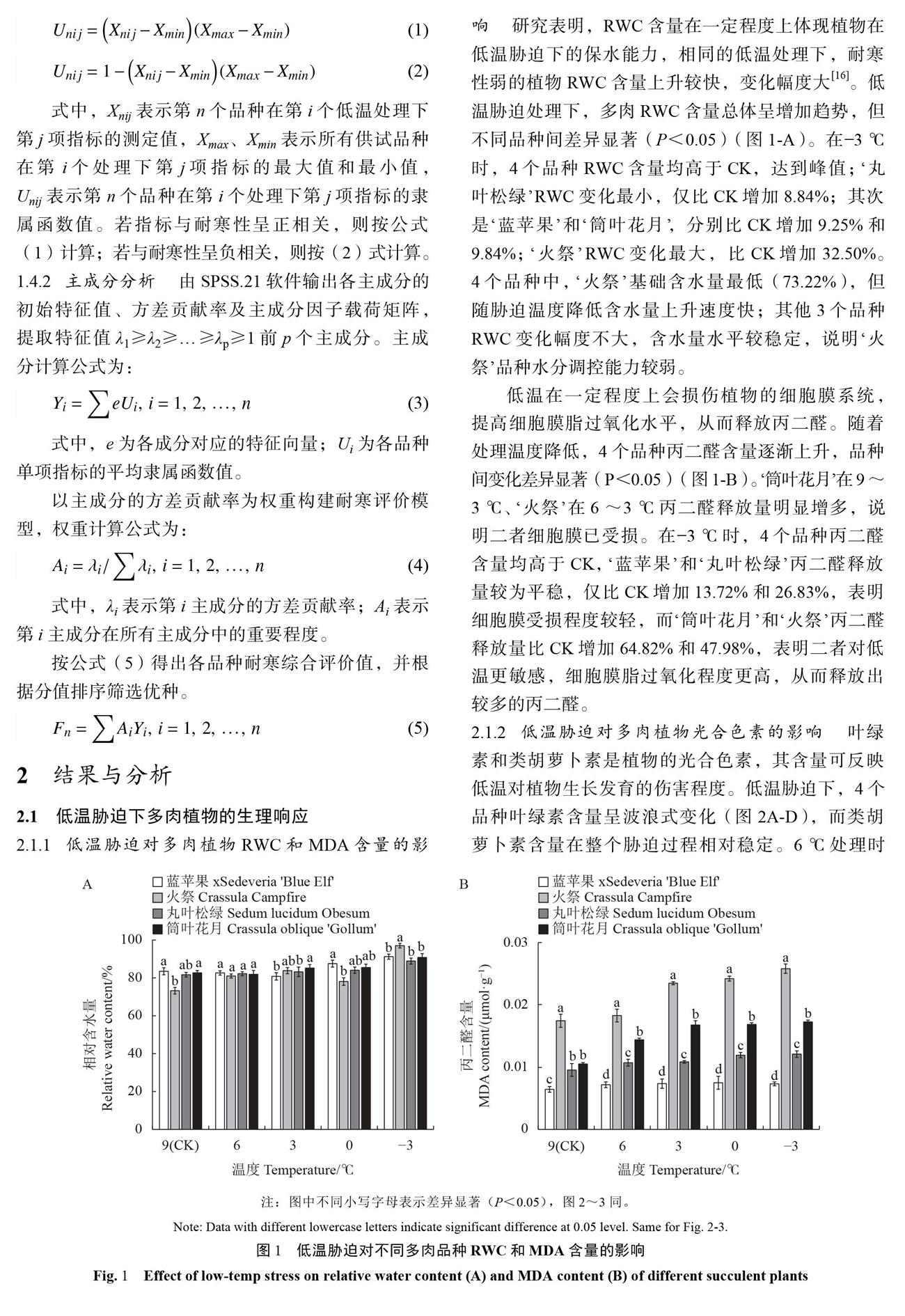
2.1.1低温胁迫对多肉植物RWC和MDA含量的影响 研究表明,RWC含量在一定程度上体现植物在低温胁迫下的保水能力,相同的低温处理下,耐寒性弱的植物RWC含量上升较快,变化幅度大[16]。低温胁迫处理下,多肉RWC含量总体呈增加趋势,但不同品种间差异显著(P<0.05)(图1-A)。在-3℃时,4个品种RWC含量均高于CK,达到峰值;‘丸叶松绿RWC变化最小,仅比CK增加8.84%;其次是‘蓝苹果和‘筒叶花月;分别比CK增加9.25%和9.84%;‘火祭RWC变化最大,比CK增加32.50%。4个品种中,‘火祭基础含水量最低(73.22%),但随胁迫温度降低含水量上升速度快;其他3个品种RWC变化幅度不大,含水量水平较稳定,说明‘火祭品种水分调控能力较弱。
低温在一定程度上会损伤植物的细胞膜系统,提高细胞膜脂过氧化水平,从而释放丙二醛。随着处理温度降低,4个品种丙二醛含量逐渐上升,品种间变化差异显著(P<0.05)(图1一B)。‘筒叶花月在9~3℃、‘火祭在6~3℃丙二醛释放量明显增多,说明二者细胞膜已受损。在-3℃时,4个品种丙二醛含量均高于CK,‘蓝苹果和‘丸叶松绿丙二醛释放量较为平稳,仅比CK增加13.72%和26.83%,表明细胞膜受损程度较轻,而‘筒叶花月和‘火祭丙二醛释放量比CK增加64.82%和47.98%,表明二者对低温更敏感,细胞膜脂过氧化程度更高,从而释放出较多的丙二醛。
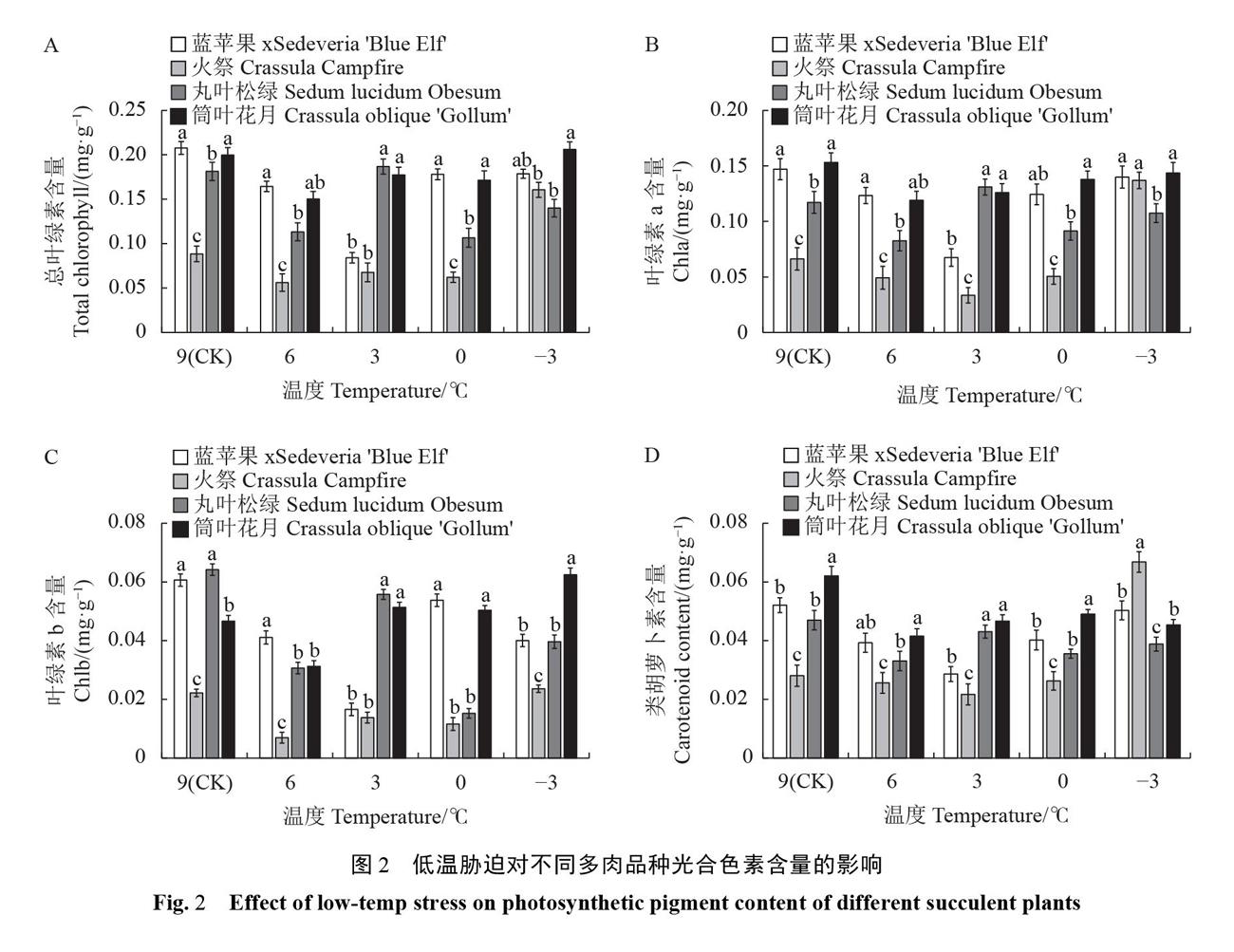
2.1.2低温胁迫对多肉植物光合色素的影响 叶绿素和类胡萝卜素是植物的光合色素,其含量可反映低温对植物生长发育的伤害程度。低温胁迫下,4个品种叶绿素含量呈波浪式变化(图2A-D),而类胡萝卜素含量在整个胁迫过程相对稳定。6℃处理时4个品种总叶绿素、叶绿素a、叶绿素b和类胡萝卜素含量均明显下降,与CK比较,‘火祭和‘丸叶松绿叶绿素a含量降低25.69%和29.49%、叶绿素b降低68.64%和52.21%,‘火祭和‘筒叶花月总叶绿素含量达到最低点,较CK降低36.47%和24.78%,‘筒叶花月类胡萝卜素含量降幅最大,较CK降低33.10%,说明6 ℃处理对多肉光合色素合成有明显影响。-3℃时,‘丸叶松绿的总叶绿素和类胡萝卜素含量均最低,‘火祭的变化幅度最大,其总叶绿素含量比CK增加81.57%,类胡萝卜素含量比CK增加137.93%。
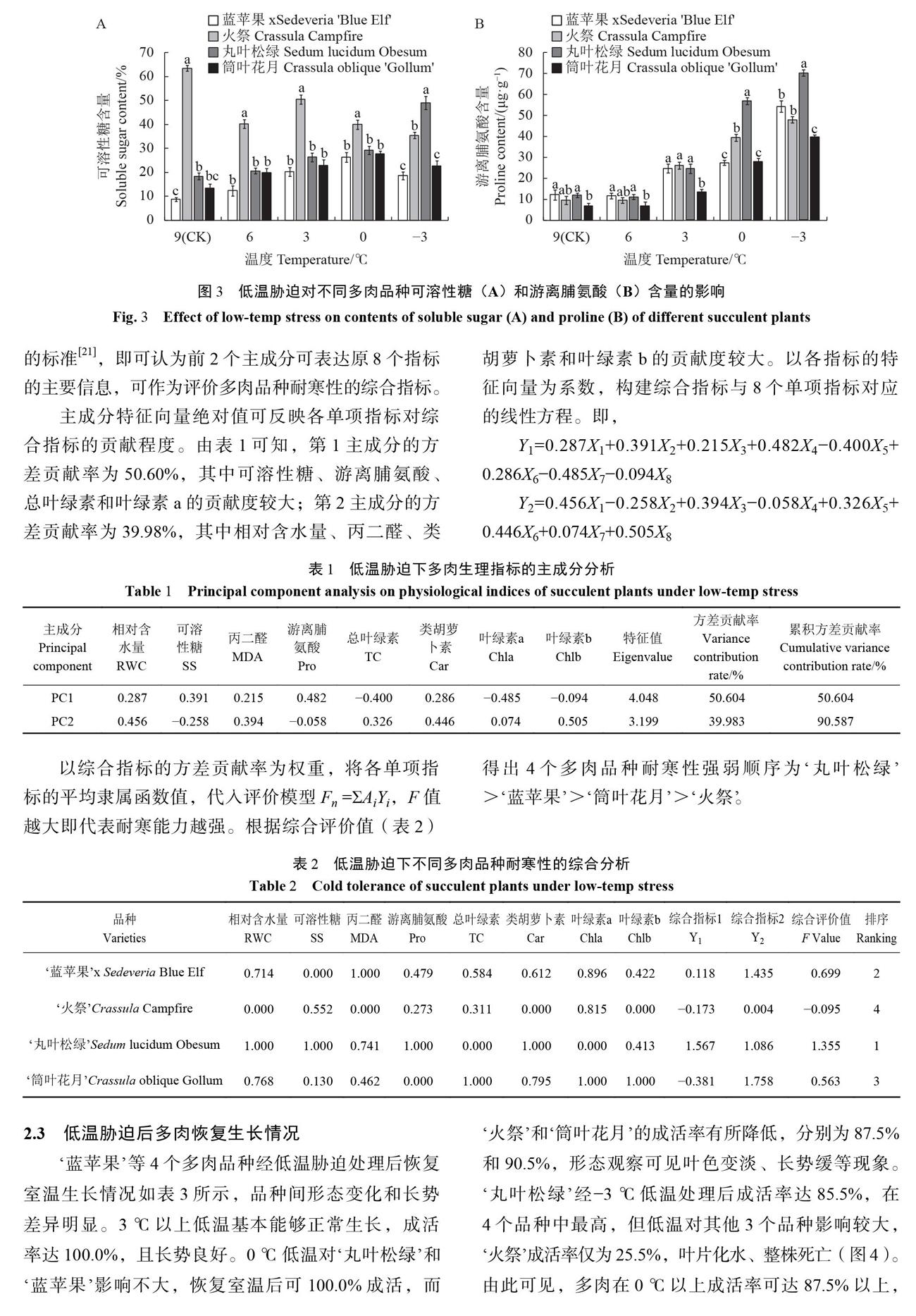
随胁迫程度加剧,4个品种光合色素含量的变化特征各不相同。‘蓝苹果和‘丸叶松绿的光合色素含量在6~0℃之间波动最大,0℃以下则趋于平稳;‘火祭的光合色素含量在0℃以下变化幅度最大;而‘筒叶花月的光合色素含量始终保持稳定,说明低温胁迫处理对不同多肉品种光合系统的影响有差异,0℃以下低温对‘火祭光合系统影响最大。2.1.3低温胁迫对多肉植物渗透调节物质的影响低温胁迫下,不同多肉品种可溶性糖含量变化趋势有差异(图3-A)。‘蓝苹果和‘筒叶花月的可溶性糖含量先上升,在0℃达到峰值后又迅速下降;‘火祭的可溶性糖含量总体呈下降趋势,仅在3℃时小幅度上升;‘丸叶松绿的可溶性糖含量则始终保持增长。在-3℃时,‘丸叶松绿的可溶性糖含量显著高于其他3个品种(P<0.05),比CK提高166.42%,积累速度最快;‘火祭可溶性糖含量降幅最大,比CK降低44.17%。图3-A还表明,‘火祭的基础可溶性糖含量最高,但随温度降低下降速度快;而‘丸叶松绿在0℃以下仍能大量积累可溶性糖,说明低温胁迫下不同多肉品种积累可溶性糖的能力有差异。
由图3-B可知,低温处理下多肉脯氨酸含量不断增加,温度越低,增幅越大。在9~6℃4个品种脯氨酸含量均较低,且品种间差异不显著。在0℃时,4个品种积累脯氨酸的速度明显加快,‘丸叶松绿脯氨酸含量增幅最大,是CK的4.79倍,增幅较小的是‘蓝苹果:是CK的2.25倍。-3℃时,4个品种仍在大量积累脯氨酸,其含量显著高于CK。‘丸叶松绿的脯氨酸积累最多,达70.29 μg.g-1,是CK的5.91倍;其次是‘蓝苹果和‘火祭:二者无显著差异;脯氨酸积累最少的是‘筒叶花月:显著低于其他3个品种(P<0.05)。
2.2 多肉耐寒性综合评价
对8个生理指标的数据标准化后进行主成分分析,提取各主成分的载荷系数、特征向量和方差贡献率,结果表明,前2个主成分的累积方差贡献率达90.59%(表1)。以主成分累积贡献率应≥85%的标准[21],即可认为前2个主成分可表达原8个指标的主要信息,可作为评价多肉品种耐寒性的综合指标。
主成分特征向量绝对值可反映各单项指标对综合指标的贡献程度。由表1可知,第1主成分的方差贡献率为50.60%,其中可溶性糖、游离脯氨酸、总叶绿素和叶绿素a的贡献度较大;第2主成分的方差贡献率为39.98%,其中相对含水量、丙二醛、类胡萝卜素和叶绿素b的贡献度较大。以各指标的特征向量为系数,构建综合指标与8个单项指标对应的线性方程。即,
以综合指标的方差贡献率为权重,将各单项指标的平均隶属函数值,代人评价模型Fn=∑AiYi,F值越大即代表耐寒能力越强。根据综合评价值(表2)得出4个多肉品种耐寒性强弱顺序为‘丸叶松绿>‘蓝苹果>‘筒叶花月>‘火祭'.而-3℃则成活率大幅度降低,表明多肉植物半致死温度在-3~0℃之间,需进一步试验探讨。
3 结论与讨论
低温冷害或冻害是限制亚热带植物生长的重要非生物因子。耐寒性強的种质在遭受低温胁迫时能迅速启动抵御低温的生理机制,诱导渗透调节基因表达[22],调控相关的代谢途径,以减轻或消除胁迫的伤害。试验结果表明,不同景天科多肉植物对低温胁迫的生理应答规律不同,抵御低温能力有显著差异;8项生理指标综合为2个主成分,累积贡献率达90.59%,生物学意义显著;综合评价得出耐寒性强弱顺序为‘丸叶松绿>‘蓝苹果>‘筒叶花月>‘火祭'.
MDA含量是反映细胞膜受损程度的重要生理指标,耐寒性强的种质在低温下MDA积累量与外渗率均较小。低温胁迫后‘丸叶松绿和‘蓝苹果丙二醛积累较少,细胞膜功能相对稳定;而‘火祭和‘筒叶花月丙二醛释放量较高,说明低温加剧二者细胞膜脂过氧化程度,导致生物膜受损较严重。耐寒性强的种质在低温胁迫下能维持更稳定的水分代谢水平,‘火祭的相对含水量增幅最快,胁迫解除后叶片失水无缓解,保水能力弱;‘丸叶松绿相对含水量变化幅度较小,叶片肉质饱满,保水能力强。
低温处理会抑制景天科植物叶绿素合成中的酶促反应,降低净光合速率(Pn)[23],限制植株生长。低温胁迫后4个品种叶绿素含量均下降,叶绿素b降幅明显大于叶绿素a,说明低温主要限制多肉叶绿素b的合成,从而影响光合作用。一3℃时叶绿素含量上升可能是因为在此温度下多肉进入濒死状态,叶绿素大量析出导致,其他物种也有类似的变化规律[24]。‘丸叶松绿,-3℃时叶绿素含量最低,但综合评价值表明其耐寒性最强,可能因为叶绿素一定程度降解有利于保护光合器官以减轻低温损伤[25-26],因此应结合多项生理指标综合判定多肉耐寒性。低温处理后类胡萝卜素含量低的种质耐寒性更强[20],0℃以下‘丸叶松绿的类胡萝卜素含量显著低于其他3个品种,耐寒性最强。
可溶性糖和Pro是植物应对低温胁迫重要的渗透调节物质。耐寒性强的种质在低温下能迅速积累可溶性糖和Pro,以提高细胞液浓度、降低冰点温度,保持细胞不至于遇冷凝固[27]。植物可溶性糖含量与耐寒性总体上呈正相关关系,而Pro含量与耐寒性的关系尚无统一定论[28]。多肉的可溶性糖和Pro含量在0℃以下积累迅速,说明0℃是多肉启动低温防御机制,积累渗透调节物质的关键点。-3℃时,‘丸叶松绿的可溶性糖与Pro含量较对照分别增加166.42%和491.17%,增幅最大,耐寒性最强;‘火祭与‘筒叶花月Pro积累较少,综合评价值也表明其较不耐寒,因此低温处理后叶片中Pro积累程度也可作为耐寒性评定的参考指标,在火龙果耐寒试验中也有此类结论[28]。
植物的耐寒性由许多数量或质量遗传基因综合决定[29],构建数学评价模型对多肉耐寒性进行综合评价,较单一的生理指标更能全面、系统反映多肉植物的耐寒性,评价结果与恢复室温生长试验观测结果基本一致。‘丸叶松绿经-3℃低温胁迫后仍保持85.5%的成活率,形态性状稳定,与‘冰花果(C.edulis)等耐寒型多肉在低温下的生理和形态表现类似[30],可繁育推广;‘火祭,耐寒能力低于其他3个品种,且胁迫解除后对其生长影响不可逆。在此评价基础上,可选择耐寒多肉种质作为育种亲本材料;还应发掘与多肉耐寒性密切关联的关键基因,在分子水平深层次分析多肉耐寒机理。
参考文献:
[1] 张先进,李素华,张旭等,我国多肉植物引种繁育与应用现状研究[J].安徽农学通报,2019,25 (7):94-95.
ZHANG X J,LI S H,ZHANG X,et al.Study on introduction,breeding and application of succulent plants in China[J].AnhuiAgricultural Science Bulletin, 2019,25(7):94-95.(inChinese)
[2] ERMN J E,HENSLEY J Plants with horticultural and ecologicalattributes for green roofs in a cool, dry climate[J]Hortscience, 2019,54 (10):1703-1711.
[3] 蔡庆生,植物生理学[M].北京:中国农业大学出版社,2014: 299-301.
[4] 胡增辉,王倩,冷平生,苔景天和联合勘察加景天对低温胁迫的生理响应[J].新疆农业科学。2013, 50 (1):77-83.
HU Z H,WANG Q, LENG P S.The physiological response of Sedumacre plant and S.kamtschaticum subsp. ellacombianum plant to lowtemperature [J].Xinjiang Agricultural Sciences. 2013, 50(1):77-83. (in Chinese)
[5] JOVANOVIC S e,ZLATKOVIC B K, STOJANOVIC G S.Chemotaxonomic approach to the central balkan Sedum species basedon distribution of triterpenoids in their epicuticular waxes [J].Chemistrv&Biodiversitv. 2016,13(4):459-465.
[6] 荆瑞,冯黎,张洁,等.基于AHP的北京地区景天属植物园林应用综合评价[J].河南农业科学,2016. 45 (10):128-133.
JING R,FENG L,ZHANG J,et al.AHP-based comprehensiveevaluation of landscape application of sedums in Beijing area EJl.Journal of Henan Agricultural Sciences, 2016, 45 (10):128-133.(in Chinese)
[7] CLARK M J,ZHENG Y B.Effect of fertilizer rate on plant growthand leachate nutrient content during production of Sedum-vegetatedgreen roof modules EJl.Hortscience, 2014, 49(6):819826.
[8] GETTER K L,ROWE D B.Substrate depth influences Sedum plantcommunity ona green roof [J].HortScience, 2009, 44(2):401-407.
[9]赵欢,刘克林,郑高言,等.多肉植物‘丽娜莲Echeveria lilacinakimnach& Moran再生体系的建立[J].分子植物育种,2018, 16 (18):6061-6067.
ZHAO H, LIU K L,ZHENG G Y, et al. Establishment of regenerationsystem of succulent plant Echeveria lilaeina kimnach&moran EJl.MolecularPlantBreeding, 2018, 16(18):6061-6067.( inChinese)
[10] MONIUSZKO-SZAJWAJ B, PECIO L, KOWALCZYK M, et al. Newbufadienolides isolated from the roots of Kalanchoe daigremontiana(Crassulaceae) [J].Molecules (Basel. Switzerland). 2016, 21 (3): 243.
[11] LOPEZ-ANGULO G, MONTES-AVILA J, DiAZ-CAMACHO S P,eta1. Bioactive components and antimutagenic and antioxidant activitiesof two Echeveria DC. species [J]. Industrial Crops and Products,2016. 85: 3848.
[12] STARRY O,LEA-COX J D, KIM J, et al.Photosynthesis and wateruse by two Sedum species in green roof substrate [J].Environmentaland Experimental Botany. 2014. 107: 105-112.
[13]岳莉然,岳樺.2种多肉植物在室内环境中的光合特性及含酸量研究[J].江苏农业科学,2012, 40 (7):171-173.
YUE L R,YUE H.Photosynthetic characteristics and acid content oftwo kinds of succulent plants in indoor environment [J]. JiangsuAgricultural Science. 2012. 40 (7):171-173.( in Chinese)
[14] KOZMINSKA A, AL HASSAN M. WISZNIEWSKA A, et aLResponses of succulents to drought: Comparative analysis of fourSedum (Crassulaceae) species [J].Scientia Horticulturae. 2019. 243:235-242.
[15] KIM I, HUH K Y, HUH M. Cold tolerance assessment of Sedumspecies for shallow-extensive green roof system EJl.Korean Journal ofHorticultural Science&Technology, 2010, 28(1):22-30.
[16]马英,许琪,谷战英,等,低温胁迫对五种景天科多肉植物生理指标的影响[J].北方园艺,2019 (1):97-102.
MA Y, XU Q, GU Z Y, et al.Effect of low temperature stress onphysiological index of five Crassulaceae succulents [J]. NorthernHorticulture. 2019 (1):97-102. (in Chinese)
[17]王璐珺,丁彦芬,田治国,低温胁迫对4种景天属植物生长和生理的影响[J].中国农业信息,2013( ll):117-120.
WANG L J,DING Y F,TIAN Z G.Effect of low temperature on physiological indexes of 4 Crassulaceae species [J]. ChinaAgriculture Information, 2013(1 1):1 17-120.( in Chinese)
[18]王学奎,黄见良,植物生理生化实验原理与技术:第3版[M].北京:高等教育出版社,2015: 171, 274-276.
[19]申惠翡,赵冰.杜鹃花品种耐热性评价及其生理机制研究[J].植物生理学报,2018, 54 (2):335-345.
SHEN H F,ZHAO B.Study on evaluation of heat tolerance and itsphysiological mechanisms in Rhododendron cultivars [J]. PlantPhysiologyCommunications, 2018, 54(2):335-345.( in Chinese)
[20]陈洁,金晓玲,宁阳,等.3种含笑属植物抗寒生理指标的筛选及评价[J].河南农业科学,2016, 45 (2):113-118.
CHEN J, JIN X L,NING Y. et al.Identification and comprehensiveevaluation of cold resistance indexes of three Michelia plants [J].Journalofllenan Agricultural Sciences. 2016, 45(2):113-1 18.( inChinese)
[21]郭英姿,贾文庆,刘会超,等.三十二个品种芍药观赏性状的主成分分析[J].北方园艺。2018 (4):110-116.
GUO Y Z,JIA W Q, LIU H C,et al. Principal component analysis ofornamental traits of thirty-two herbaceous peony varieties [J].Northern Horticulture. 2018 (4):1 10-1 16.( in Chinese)
[22]吴海波.枇杷幼果在低温胁迫下基因的差异表达[D].福州:福建农林大学,2010.
WU H B.mRNA differential expression of loquatfruit under chillingstress[D]. Fuzhou: Fujian Agriculture and Forestry University, 2010.(in Chinese).
[23] PAGTER M, PETERSEN K K, KJAER K H.Direct and indirecteffects of shoot- and/or root-chilling stress on growth, photosynthesis,and osmotic root water uptake in Kalanchoe blossfeldiana Poelln.‘Molly' [J].Journal ofHorticultural Science&Biotechnology, 20 13,88 (5):571-579.
[24]李静,陈秀龙,李志阳,等.低温胁迫对10个油棕新品种生理生化特性的影响[J].华南农业大学学报,2013. 34 (1):62-66.
LI JING", CHEN XIULONGI, LI ZHIYANG ZHENG., et alPhysiological and biochemical responses of 10 new varieties of Elaeisguineensis to low temperature stress [J]. Journal of South ChinaAgricultural University, 2013, 34(1):62-66.( in Chinese)
[25] GOLOVKO T K,DALKE I V,BACHAROV D S.Mesostructure andactivity of photosynthetic apparatus for three crassulacean speciesgrown in cold climate [J].Russian Journal ofPlant Physiology, 2008,55 (5):603-612.
[26]岳海,李国华,李国伟,等.澳洲坚果不同品种耐寒特性的研究[J].园艺学报,2010, 37 (1):31-38.
YUEH, LI G H,LI G W, et al. Studies on cold resistance of differentMacadamia cultivars [Jl.Acta Horticulturae Sinica, 2010. 37(1):31-38. (in Chinese)
[27]李付鹏,伍宝朵,秦晓威,等,可可种质资源抗寒性初步评价及低温胁迫下生理响应[J].热带作物学报,2019, 40 (11):2135-2141.
LI F P,WU B D,QIN X W. et al.Preliminary evaluation on coldresistance of cacao germplasm resources and physiological responseunder low temperature stress [J] Chinese Journal of Tropical Crops,2019, 40( 11): 2135-2141. (in Chinese)
[28]邓仁菊,范建新,王永清,等,低温胁迫下火龙果的半致死温度及抗寒性分析[J].植物生理学报,2014, 50 (11):1742-1748.
DENG R J,FAN J X,WANG Y Q,et al-Semilethal temperature ofpitaya under low temperature stress and evaluation on their coldresistance [J]. Plant Physiology Communications, 2014. 50( 11):1742-1748. (in Chinese)
[29]李倩,王月异,王玉祥,低温处理对白三叶叶绿素、可溶性糖和丙二醛等的影响[J].中国农学通报,2016, 32( 19):83-86.
LI Q,WANG Y Y,WANG Y X.Effects of temperature stress onchlorophyll content, soluble sugar and MDA in Trifolium repensL [J].
Chinese Agricultural Science Bulletin, 2016. 32 (19):83-86. (in Chinese)
[30]VAHDATI N, TEHRANIFAR A, KAZEMI F. Assessing chilling anddrought tolerance of different plant genera on extensive green roofs inan arid climate region in Iran [J]. Journal of EnvironmentalManagement, 2017. 192: 215-223.
(責任编辑:黄爱萍)
作者简介:陈艺荃(1989-),女,硕士研究生,研究实习员,研究方向:园林植物资源评价与栽培生理(E-mail:
yiquanchenl 107@sina.com)
通信作者:叶秀仙(1977-),女,副研究员,研究方向:花卉生物技术(E-mail: yxx7861@163.com)
猜你喜欢
江苏农业科学(2017年1期)2017-02-27
吉林农业·下半月(2017年1期)2017-02-16
安徽农学通报(2017年1期)2017-02-15
物联网技术(2016年12期)2017-01-21
科学与财富(2016年26期)2016-12-01
医学信息(2016年29期)2016-11-28
大学教育(2016年11期)2016-11-16
企业导报(2016年9期)2016-05-26
江苏农业科学(2015年8期)2015-09-10
江苏农业科学(2015年8期)2015-09-10
