
二倍体与四倍体苦荞的充实差异研究
2020-02-21 00:18张余何佩云罗庆华吴兴慧孔德章黄小燕黄凯丰
福建农业学报 2020年10期
关键词:苦荞
张余 何佩云 罗庆华 吴兴慧 孔德章 黄小燕 黄凯丰


摘要:【目的】比较分析二倍体和四倍体苦荞材料的根系、灌浆特性等相关生理性状,明确四倍体苦荞充实差的生理原因。【方法】以二倍体苦荞品种小黄荞和六苦2081及其四倍体苦荞材料TB193、TB198为试验材料,比较分析了四倍体和二倍体苦荞间灌浆特性、淀粉合成酶活性、根系形态生理、农艺性状、充实度及产量的差异。【结果】2个苦荞品种及其四倍体苦荞材料在花后25 d百粒重几乎不再增加,且二倍体苦荞的百粒重高于四倍体苦荞;四倍体苦荞的灌浆起始势、最大灌浆速率和平均灌浆速率低于二倍体苦荞,达最大灌浆速率的天数高于二倍体;灌浆前期二倍体苦荞籽粒的AGPase和SSS活性强于四倍体苦荞,中后期则低于四倍体苦荞;四倍体苦荞的根系长度、根系表面积、根系体积、根系活力、根系伤流液总体显著高于二倍体苦荞;二倍体苦荞的株高、主茎分枝数、主茎节数、1~2节节间粗度、1~2节节间粗度总体均低于四倍体苦荞,充实度和产量则分别是四倍体苦荞的2.97和1.23倍(两品种的平均值)。【结论】较低的灌浆起始势、灌浆速率,以及灌浆前期较低的淀粉合成酶活性是四倍体苦荞充实较差的重要生理原因。
关键词:苦荞;四倍体;灌浆特性;淀粉合成酶活性;根系形态生理;充实度
中图分类号:S 517
文献标志码:A
文章编号:1008-0384( 2020) 10-1043-07
0 引言
【研究意義】荞麦(Fagopyrum Miller)起源于中国,在世界上分布广泛,遍及亚洲和欧洲,主要有苦荞和甜荞2个栽培种。其中苦荞(Fagopyrumtataricum),又名鞑靼荞麦(Tartary buckwheat),为我国的主要栽培种。Rout[1]、Li and Zhang[2]和陈庆富[3]等的研究表明:苦荞含有丰富的黄酮类物质、D-手性肌醇(D-CI)、活性蛋白及活性多肽、丰富的矿质元素等[4],“三降”功能明显,还对癌症等疾病有较好的预防效果[5]。苦荞具有较高的研究和开发价值。但生产上苦荞的产量较低[6],实现苦荞的高产亟待解决[7]。研究表明,四倍体苦荞是由二倍体苦荞加倍而成,隐含着更高的增产潜力和更广的适应性[8-9],但是实际生产上四倍体苦荞的产量较低,其主要原因是籽粒充实度较差。因此,明确四倍体苦荞充实差的生理机理,对于提出切实可行的栽培调控措施、提高苦荞的产量等有重要的理论和实际意义。【前人研究进展】赵钢等[10]的研究发现,四倍体苦荞的细胞和组织器官比二倍体明显增大,且根系发达,茎秆粗壮,叶面积增大,叶厚度增加,籽粒增大,产量明显超过原种二倍体。廉立坤等[8]的研究发现,四倍体苦荞与二倍体苦荞相比具有生长势强、生物学产量高、籽粒大、籽粒品质好和抗逆性强等特点。李振宙等[9]的研究发现,四倍体苦荞的充实度较差,一般在20%~30%,严重影响了其在生产上的推广和利用,适宜的钾肥施用量,能促进四倍体苦荞的生长发育,同时还能提高其籽粒的充实度,促进了四倍体苦荞产量的提高。【本研究切人点】有关四倍体苦荞充实较差的生理机制还未有明确定论。【拟解决的关键问题】为了明确四倍体苦荞充实差的生理原因,本研究以二倍体苦荞品种小黄荞和六苦2081及其四倍体TB193和TB198为试验材料,比较分析了四倍体和二倍体苦荞间灌浆特性、根系形态生理、农艺性状、充实度及产量的差异,以期明确四倍体苦荞充实差的生理机制,为指导苦荞的高产栽培等提供理论和实践意义。
1 材料与方法
1.1 试验材料
供试苦荞品种为二倍体苦荞品种小黄荞和六苦2081,及其对应的四倍体苦荞材料TB193和TB198(利用秋水仙素对二倍体苦荞品种进行加倍而获得),分别以E1、E2和SI、S2表示,上述材料由贵州师范大学荞麦产业技术研究中心提供。
1.2 试验方法
试验于2018年4月10日在贵州师范大学荞麦产业技术研究中心黄泥塘试验基地进行。池栽,池子长5m,宽2m,深0.3 m。池中装有新移的黄壤土,前作空闲,土壤有机质34.45 g.kg-1,全氮1.10 g.kg-1,水解氮118 mg.kg-1,有效磷21.46 mg.kg-1,速效钾112.28 mg.kg-1,pH5.77。氮磷钾肥分别以100 kg.hm-2(尿素)、69 kg.hm-2(过磷酸钙)和5.1 kg.hm-2(氯化钾)的最适施用量混匀后作为基肥1次施人[11]。每份苦荞资源种植3个水泥池,每个水泥池作为1次重复,共计12个水池。条播,行距0.33 m,播种量为3.65 g.m-2,基本苗约90~100株.m-2。各小区中有70%籽粒成熟时采收(2018年7月12日)。
1.3 取样方法
从苦荞开花起选择相似的典型植株进行挂牌标记,第1次取样为开花后第5d,此后每隔5d取样1次,直到籽粒成熟。将所取的苦荞籽粒去壳后,取其中1/2籽粒置液氮冷冻1 min后放入-70℃冰箱保存,用于测定淀粉合成酶活性,另1/2置105℃烘箱中杀青10 min后,于60℃烘干至恒重,用于测定苦荞的灌浆特性。参考李振宙等[9]的方法取样,用于测定苦荞的根系形态、根系活力、伤流液及农艺性状。
1.4 指标测定
1.4.1 灌浆特性测定 参照朱庆森等[12]的方法利用Richards方程对籽粒生长过程进行拟合:W=A(1+BeKt(-l=1/N)。参考杨志远等[13]的方法计算灌浆前、中、后期时间段内灌浆物质对总灌浆物质的贡献率。
1.4.2淀粉合成相关酶活性的测定 参考Yang等[14]的方法测定腺苷二磷酸葡萄糖焦磷酸化酶( ADP-glucose pyrophosphorylase,AGPase)、可溶性淀粉合酶( Soluble starch synthase,SSS)和淀粉分支酶(Starchbranching enzyme,SBE)的活性。
1.4.3根系形态生理测定 参考李振宙等[9]的方法测定根系形态指标;参考金成忠等[15]的方法测定苦荞一昼夜伤流强度的变化情况。采用TTC法测定根系活力[16]。
1.4.4农艺性状及产量的测定 参考张宗文等[17]的方法测量株高、主茎分枝数、主茎节数、1~2节节间粗度、1~2节节间长度、子叶节高度;参照李振宙等[9]的方法测定籽粒的充实度和产量。
1.5 数据分析
采用Excel 2003软件进行数据处理,SPSS 20.0软件对数据进行差异性分析。
2 结果与分析
2.1 四倍体和二倍体苦荞的百粒重
由表1可以看出,4个供试苦荞材料的百粒干重随生育期的推进呈持续增加的变化趋势,花后15d之前增幅较大,其后增幅有所下降,花后30d以后4个苦荞材料的百粒重趋于稳定。苦荞材料间比较,同时期四倍体苦荞的百粒干重均低于二倍体苦荞,其中花后5、10、15 d的差异达到显著水平。
2.2 Richards方程模拟苦荞籽粒的灌浆过程
由表2可以看出,各苦荞材料的曲线方程的决定系数R2=0.995~1.000,说明用Richards方程拟合苦荞的灌浆过程是可行的。四倍体苦荞的A值均低于二倍体苦荞。4个苦荞的Ⅳ值均小于1,且以四倍体苦荞的Ⅳ值高于二倍体苦荞。四倍体苦荞的灌浆起始势( Ro)显著低于二倍体苦荞,E1和S1的最大灌浆速率( Gmax)和平均灌浆速率(Gmean)无明显差异,E2的Gmax和G mean显著高于S2;达最大灌浆速率的时间( Tmax.G)则明显高于二倍体苦荞;灌浆速率为最大时的生长量与籽粒生长终值量比值(I)苦荞材料间的差异不显著。
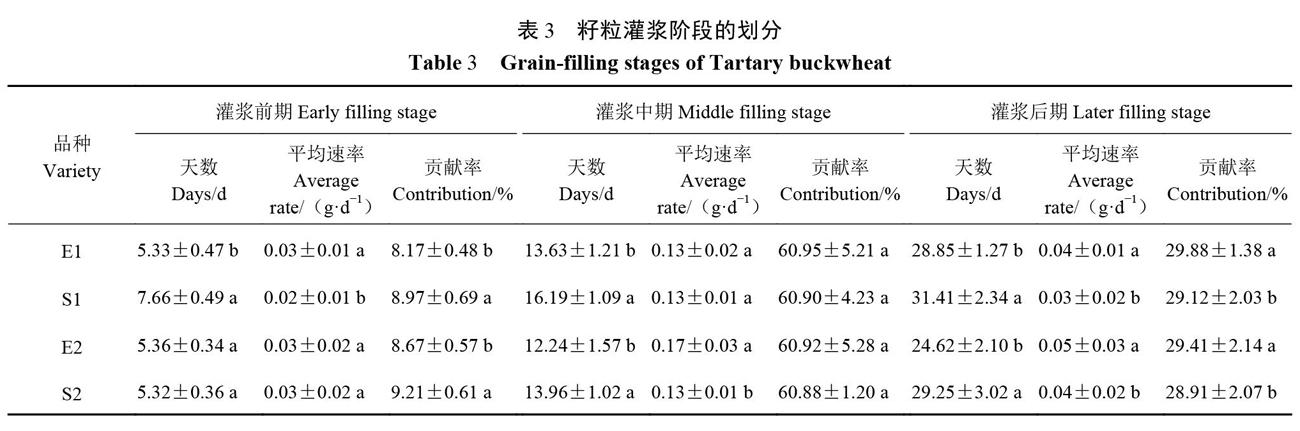
2.3 四倍体和二倍体苦荞籽粒灌浆阶段划分
由表3可以看出,灌浆前期持续的时间E1显著小于S1,E2和S2间则无显著差异;灌浆中期和灌浆后期持续的时间则都以二倍体苦荞显著小于四倍体苦荞。灌浆前期的平均速率S1显著小于E1,E2和S2间无明显差异;灌浆中期和后期总体以四倍体苦荞显著小于二倍体苦荞。籽粒灌浆前期对籽粒粒重的贡献率二倍体苦荞显著小于四倍体苦荞,灌浆中期对籽粒粒重的贡献率以四倍体苦荞小于二倍体苦荞,但差异不显著,灌浆后期对籽粒粒重的贡献率则以四倍体苦荞显著小于二倍體苦荞。由表3还可以看出,各灌浆时期对苦荞籽粒粒重的贡献率存在差异,表现为灌浆中期>灌浆后期>灌浆前期。
2.4 四倍体和二倍体苦荞淀粉合成酶活性
由表4可以看出,苦荞籽粒的AGPase活性随生育期的推进呈先增加后降低的趋势,其中二倍体苦荞的AGPase活性在花后10 d达最大,四倍体苦荞的AGPase活性则在花后15 d达最大;灌浆前期(花后10 d之前)以二倍体苦荞的AGPase活性较大,灌浆中后期(15 d以后)则以四倍体苦荞的AGPase活性较大。4个苦荞材料的SSS活性在花后10 d达最大,其后快速下降;花后10 d之前以二倍体苦荞的SSS活性高于四倍体,其后则低于四倍体苦荞。苦荞籽粒的SBE活性随生育期的推进呈先增加后降低的趋势,以二倍体苦荞的SBE活性显著高于四倍体。2.5四倍体和二倍体苦荞的根系形态生理比较
由表5可以看出,苗期S1的根系长度、根系表面积、根系体积显著高于E1,根系平均直径、根系伤流液、根系活力及根冠比则无明显差异;S2的根系形态生理指标则均显著高于E2。开花期,四倍体苦荞的根系长度、根系表面积、根系体积、根系伤流液显著高于二倍体苦荞,根系平均直径无明显差异;根系活力以四倍体苦荞高于二倍体苦荞,其中S1和E1的差异达到显著水平。灌浆期,四倍体苦荞的根系长度显著低于二倍体苦荞.而根系表面积、根系体积、根系伤流液和根系活力则显著高于二倍体苦荞;四倍体苦荞的根系平均直径高于二倍体,其中S2和E2的差异达到显著水平。成熟期,四倍体苦荞的根系长度、根系表面积、根系体积、根系伤流液均显著高于二倍体苦荞,根系平均直径和根系活力以四倍体苦荞较高,S1的根系活力显著高于E1。
2.6 四倍体和二倍体苦荞的农艺性状及产量
由表6可以看出,四倍体苦荞的株高、主茎节数、主茎分枝数、1~2节节间长度、1~2节节间粗度均显著高于二倍体苦荞;四倍体苦荞和二倍体苦荞的子叶节高度无明显差异;二倍体苦荞的充实度和产量均显著高于四倍体苦荞。
3 讨论与结论
3.1 四倍体与二倍体苦荞的根系形态生理差异
根系的形态影响植物对水分和养分的吸收,进而影响植株地上部的生长、养分供应[18]、产量和品质[19]形成。潘晓华等[20]、张成良等[21]的研究发现水稻根数、根长、根系表面积、根系体积、根系活力、根系伤流量与籽粒灌浆强度、充实度、千粒重和产量呈显著或极显著的正相关关系。本试验的研究发现,四倍体苦荞的根系长度、根系表面积、根系体积、根系直径、根系活力和根系伤流量总体均明显大于二倍体苦荞,但四倍体苦荞的充实度和产量则显著低于二倍体苦荞,这与杨建昌等[22]和Samejima等[23]在水稻上的研究结果一致,可能是因为根系虽然是水分和养分的吸收器官,但同时又要为根系的建成而消耗地上部提供的光合产物以维持生长,如果根量过大反而会造成无效消耗而对产量形成产生不利的影响[24-25]。也有可能是因为四倍体苦荞相对二倍体苦荞来说,根系的干物质积累量大、根系活力高、根系代谢旺盛会形成与籽粒灌浆物质竞争的“次库”:从而影响籽粒的增重[26],导致充实度较差,产量较低。具体机制有待于进一步的研究证实。
3.2 四倍体与二倍体苦荞的灌浆特性与淀粉合成酶活性的差异
Richards方程生长曲线是由N值大小决定的一簇曲线,当01时,曲线拐点大于0.500 A,并随N的增大而逐渐接近于A,灌浆速率曲线右偏[27-28]。本试验的研究结果发现参试苦荞品种的N值均<1,表明其灌浆受库容的限制较大,即灌浆物质相对充分[12,29],表现为灌浆前期生长迅速,其后逐渐减弱。从本试验的研究结果还可以看出,四倍体苦荞的灌浆起始势(R0)均明显低于二倍体苦荞品种,这可能与四倍体苦荞相对于二倍体苦荞而言,籽粒具有较大的库容(即籽粒大小),灌浆初期生长速率较慢,胚乳细胞分裂周期较长,分裂速度较慢有关[12]。四倍体苦荞的R0较低,暗示着籽粒灌浆启动较晚,得到的光合产物不足,导致开花后仍需较长的时间才能达到最大灌浆速率,充实较差。结合四倍体苦荞具有较低的最大灌浆速率和平均灌浆速率,以及具有较高的达最大灌浆速率天数的研究结果,认为这些可能是导致四倍体苦荞充实较差的主要生理原因。
苦荞籽粒中的淀粉含量在75%左右[30],因此,苦荞籽粒的充实过程,其实就是淀粉的累积过程。源器官的光合同化物以蔗糖的形式经韧皮部运输到籽粒,之后在一系列酶的作用下形成淀粉[31-32]。焦磷酸化酶( ADPG)、可溶性淀粉合酶(SSS)、淀粉分支酶( SBE)等是淀粉合成代谢过程中的关键酶,这些酶对籽粒淀粉的合成、积累起着重要的调节作用。从本试验的研究结果可以看出,灌浆前期二倍体苦荞的AGPase和SSS活性高于四倍体苦荞,灌浆中后期则低于四倍体苦荞,这可能与四倍体苦荞灌浆前期光合同化物从“源”输送到“库”的转运能力弱于二倍体苦荞有关,致使四倍体苦荞灌浆前期籽粒的生理活性较低,蔗糖降解的速度慢,导致籽粒淀粉合成慢、累积少,最后籽粒充实不良[33]。结合四倍体苦荞具有较低灌浆速率的研究结果,认为四倍体苦荞在灌浆前期具有较低的淀粉合成酶活性是其灌浆速率小、粒重轻的一个重要原因[34]。
综上,较低的灌浆起始势、灌浆速率,以及灌浆前期较低的淀粉合成酶活性是四倍体苦荞充实较差的重要生理原因。
参考文献:
[1] ROUT M K,CHRUNGOO N K,RAO K S.Amino acid sequence ofthe basic subunit of 13S globulin of buckwheat [J]. Phytochemistry,1997, 45 (5):865-867.
[2] L1 S Q,ZHANG Q H.Advances in the development of functionalfoods from buckwheat [J]. Critical Reviews in Food Science andNutrition, 2001, 41(6):451-464.
[3] 陈庆富,荞麦生产状况及新类型栽培荞麦育种研究的最新进展[J].贵州师范大学学报(自然科学版),2018. 36 (3):1-7.
CHEN Q F.The status of buckwheat production and recent progressesof breeding on new type of cultivated buckwheat EJl. Journal ofGuizhou Normal University (Natural Science Edition), 201 8,36(3):1-7. (in Chinese)
[4]HUANG X Y,ZELLER F J,HUANG K F,et al.Variation of majorminerals and trace elements in seeds of Tartary buckwheat (Fagopyrum tataricum Gaertn.) EJl. Genetic Resources and CropEvolution. 2014, 61 (3): 567-577.
[5] 武素平,魯纯静.荞麦面对血脂及脂肪肝影响的动物实验[J].食品科学,1988.2 (2):10-11.
WU S P,LU C J.Animal experiment on the effect of buckwheat onblood lipid and fatty live EJl. Food Science. 1988,2 (2):10-11. (in Chinese)
[6]宋毓雪,郭肖,杨龙云,等.不同氮磷钾肥料处理对苦荞籽粒充实度及产量的影响[J].浙江农业学报,2014. 26 (6):1568-1572.
SONG Y X,GUO X,YANG L Y,et al.Effects of different NPKtreatments on the yield and plumpness of Tartary buck,Arheat [J].Acta Agriculturae Zhejiangensis, 2014. 26 (6):1568-1572. (inChinese)
[7] 黄凯丰,李振宙,王炎,等.我国荞麦高产栽培生理研究进展[J].贵州师范大学学报(自然科学版),2019, 37(1):115-120.
HUANG K F,LI Z Z,WANG Y,et al.Research progress onphysiology of buck.Arheat under high-yield cultivation EJl.Journal ofGuizhou Normal University (Natural Science Edition), 2019,37(1):115-120. (in Chinese)
[8] 廉立坤,陈庆富.二倍体和四倍体苦荞种子蛋白质含量和黄酮含量比较研究[J].种子,2013, 32 (2):1-5.
LIAN L K,CHEN Q F.A comparative study of seed protein contentand seed flavonoid content between diploid and tetraploid Tartarybuck,Arheat [J]. Seed. 2013, 32 (2): 1-5. (in Chinese)
[9] 李振宙,吴兴慧,张余,等.钾肥用量对四倍体苦荞籽粒灌浆特性、充实度的影响[J].福建农业学报,2019, 34 (8):883-888.
LI Z Z,WU X H,ZHANG Y,et al.Effect of potassium fertilizationson grouting and plumpness of tetraploid Tartary buckwheat grains EJl.Fujian Journal of Agricultural Sciences, 2019, 34 (8):883-888.(in Chinese)
[10]趙钢,唐宇,同源四倍体苦荞新品系与原种主要性状的比较研究[J].科技通报,1994, 10 (5):321-325.
ZHAO G,TANG Y.Comparative study on main characters of a newstrain of autotetraploid and its autodiploid parent stock in Tartarybuck,Arheat [J]. Bulletin of Science and Technology, 1994, 10 (5):321-325. (in Chinese)
[11]宋毓雪,陈小娥,魏让,等,不同肥料配比对甜荞产量和品质的影响[J].中国土壤与肥料,2014 (3):49-53.
SONG Y X,CHEN X E,WEI R et al.Effects of different ratios ofNPK fertilizer on yield and quality of common buckwheat EJ] Soilsand Fertilizers Sciences加China, 2014 (3):49-53. (in Chinese)
[12]朱庆森,曹显祖,骆亦其.水稻籽粒灌浆的生长分析[J].作物学报,1988, 14 (3):182-193.
ZHU Q S,CAO X Z,LUO Y Q.Growth analysis on the process ofgrain filling in rice EJl. Acta Agronomica Sinica, 1988. 14 C3):182-193. (in Chinese)
[13]杨志远,孙永健,徐徽,等,栽培方式与免耕对杂交稻II优498灌浆期根系衰老和籽粒灌浆的影响[J].中国农业科学,2013, 46 (7):1347-1358.
YANG Z Y. SUN Y J,XU H,et al_Influence of cultivation methodsand no-tillage on root senescence at filling stage and grain-fillingproperties of eryou 498 [J]. Scientia Agricultura Sinica. 2013.46 (7): 1347-1358. (in Chinese)
[14] YANG J C,ZHANG J H,WANG z Q, et al.Activities of enzymesinvolved in sucrose-to-starch metabolism in rice grains subjected towater stress during filling EJ] Field Crops Research, 2003. 81 (1):69-81.
[15]金成忠。许德威作为根系活力指标的伤流液简易收集法[J].植物生理学通讯.1959 (4):51-53.
JINCZ,XUD W.A simple collection method of injured fluid as anindex of root activity EJl. Plant Physiology Journal., 1959 (4):51-53. (in Chinese)
[16]张志良,植物生理学实验指导[M].北京:高等教育出版社,1990.
[17]张宗文,林汝法,荞麦种质资源描述规范和数据标准[M].北京:中国农业出版社,2007.
[18]陈磊,王盛锋,刘荣乐,等.不同磷供应水平下小麦根系形态及根际过程的变化特征[J].植物营养与肥料学报,2012. 18 (2):324- 331.
CHEN L,WANG S F,LIU R L,et al_Changes of morphology andthizosphere processes of wheat under different phosphate supply EJl.Plant Nutrition and Fertilizer Science.,2012,18(2):324331(inChinese)
[19]杨建昌.水稻根系形态生理与产量、品质形成及养分吸收利用的关系[J].中国农业科学,2011. 44 (1):36-46.
YANG J C.Relationships of rice root morphology and physiologywith the formation of grain yield and quality and the nutrientabsorption and utilization EJl. Scientia Agricultura Sinica, 2011,44(1):36-46. (in Chinese)
[20]潘晓华,王永锐,傅家瑞.水稻根系生长生理的研究进展[J]植物学通报,1996 (2):13-20.
PANXH,WANGYR,FU JR.Advance in the study on the growth-physiology in rice of root system (Oryza sativa) EJl.Chinese BulletinofBotany, 1996 (2): 1320. (in Chinese)
[21]张成良,姜伟,肖叶青,等.水稻根系研究现状与展望[J].江西农业学报,2006. 18 (5):23-27.
ZHANG C L,JIANG W, XIAO Y Q, et al_Status and prospects ofresearch on rice root systems EJl. Acta Agriculturae Jiangxi, 2006,18 (5): 2327. (in Chinese)
[22]杨建昌,徐国伟,仇明,等.新株型水稻生育特性及产量形成特点的研究[J].扬州大学学报(农业与生命科学版),2002. 23 (1):45-50.
YANG J C,XU G W, QIU M, et al.Growth and developmentcharacteristics and yield formation of new plant type rice [J]. Journalof Yangzhou University (Agricultural and Life Sciences Edition), 2002,23 (1): 4550. (in Chinese)
[23] SAMEJIMA H,KONDO M, ITO O,et al_Characterization of rootsystems with respect to morphological traits and nitrogen-absorbingability in the new plant type of tropical rice lines [J] Journal of PlantNutrition, 2005, 28 (5):835-850.
[24]蔡昆爭,骆世明,段舜山,水稻根系在根袋处理条件下对氮养分的反应[J].生态学报,2003, 23(6):1109-1116.
CAI K Z,LUO S M, DUAN S S.The response of the rice root systemto nitrogen conditions under-root confinement [J]. Acta EcologicaSinica, 2003, 23(6):1109-1116. (in Chinese)
[25]刘文兆,李秧秧断伤作物根系对籽粒产量与水分利用效率的影响研究现状及问题[J].西北植物学报,2003, 23(8):1320-1324.
LIUWZ,LIY Y. Effect of crop root-cutting on grain yield and wateruse efficiency:a review EJl. Acta Botanica Boreali-OccidentaliaSinica, 2003, 23(8):1320-1324. (in Chinese)
[26]蔡永萍,杨其光。黄义德,水稻水作与旱作对抽穗后剑叶光合特性、衰老及根系活性的影响[J].中国水稻科学,2000. 14 (4):219- 224.
CAI Y P,YANG Q G, HUANG Y D.Effect of rice cultivated underpaddy and upland condition on photosynthesis and senescence of flagleaf and activity of root system afier heading EJl Chinese Journal ofRice Science, 2000, 14 (4): 219224. (in Chinese)
[27] LIANG C G. SONG Y X. GUO X. et al.Characteristics of the grain-filling process and starch accumulation of high-yield common buckwheat 'Cy. Fengtian l' and Tartary buckwheat 'Cy. Jingqia0 27[J]Cereal Research Communications, 2016, 44(3):393-403.
[28] WANG Y, SONG Y X, ZHANG Y, et al_Characteristics of starchsynthesis and grain filling of common buck,Arheat[J]Journal ofCereal Science, 2017, 73:1 1 6- 121.
[29]陈昱利,张海军,葛道阔,等,小麦籽粒灌浆速率的模拟[J].江苏农业学报,2014. 30 (3):480-485.
CHEN Y L,ZHANG H J,GE D K. et al_Simulation of grain fillingrate for wheat EJl. Jiangsu Journal of Agricultural Sciences, 2014.30 (3): 480-485. (in Chinese)
[30] STEADMAN K J,BURGOON M S,LEWIS B A, et aL Minerals,phytic acid, tannin and rutin in buckwheat seed milling fractions EJl.Journal of the Science of Food and Agriculture, 2001, 81( 11):1094-1100.
[31] NAKAMURA Y, YUKI K. Changes in enzyme activities associatedwith carbohydrate metabolism during the development of riceendosperm EJl. Plant Science, 1992, 82 (1): 15-20.
[32] JENGTL,WANGC S,CHENC L,et al_ Effects of grain position onthe panicle on starch biosynthetic enzyme activity in developing grainsof rice cultivar Tainung 67 and its NaNi-induced mutant EJl. TheJournal ofAgricultural Science. 2003. 141( 3/4):303-311.
[33]劉立军,王志琴,杨建昌,等,亚种间杂交稻籽粒灌浆特性及其生理机制[J].扬州大学学报(自然科学版),2000,3 (3):35-40.
LIU L J,WANG Z Q,YANG J C,et al.Grain-filling characteristicsand physiological mechanism of intersubspecific hybrid rice [Jl.Journal of Yangzhou University (Natural Science Edition), 2000.3 (3): 35—40. (in Chinese)
[34]付景,徐云姬,陈露,等,超级稻花后强、弱势粒淀粉合成相关酶活性和激素含量变化及其与籽粒灌浆的关系[J].中国水稻科学,2012. 26 (3):302310.
FU J, XU Y J,CHEN L,et al_Post-anthesis changes in activities ofenzymes related to starch synthesis and contents of hormones insuperior and inferior spikelets and their relation with grain filling ofsuper rice EJl. Chinese Journal of Rice Science, 2012,26 (3):302-310. (in Chinese)
(责任编辑:张梅)
作者简介:张余(1999-),女,硕士研究生,主要从事荞麦栽培生理研究(E-mail: zhangyu19990000@163.com)
通信作者:黄凯丰(1979-),男,博士,教授,主要从事荞麦栽培生理研究(E-mail: hkfl979@163.com)
猜你喜欢
诗潮(2021年11期)2021-11-24
农产品市场周刊(2021年15期)2021-10-08
农产品市场周刊(2020年15期)2020-09-22
农民致富之友(2020年26期)2020-09-12
现代农业科技(2020年13期)2020-08-04
小品文选刊·印象大同(2020年6期)2020-06-29
食品界(2017年9期)2017-09-30
农业知识·百姓新生活(2017年1期)2017-03-06
科学种养(2015年9期)2015-09-11
农村农业农民·B版(2014年12期)2015-01-04
