
不同氮素供应水平对菘蓝生长及药材质量的影响
2019-08-29 01:52关佳莉张梦蕊曹艺雯唐晓清王康才
核农学报 2019年10期
关佳莉 王 刚 张梦蕊 陈 曦 曹艺雯 唐晓清 王康才
(南京农业大学中药材研究所,江苏 南京 210095)
氮素作为植物生长所必需的营养元素之一,是植物体内叶绿素、蛋白质、核酸和部分激素等物质的组成成分,在药用植物生长发育、产量与品质的形成过程中发挥着重要作用[1]。菘蓝(IsatisindigoticaFort.)是大宗药材板蓝根与大青叶的基原植物。过量施用氮肥是其栽培生产上存在的主要问题之一,不仅导致氮素利用效率下降、生产成本增加,引发生态环境污染等问题,而且会对药材产量及药用品质产生不利影响[2-4]。因此,合理施用氮肥是获取高产优质药材的重要环节,即对药用植物而言,应在不显著降低其产量的条件下,适当减少氮肥施用量,以保证药材品质,降低生产成本和保护环境。
减量施氮对作物的影响已有大量报道。研究发现适当减量施氮不仅未影响甘蓝[5]、小麦[6]、水稻[7]等作物的产量,还减少了氮肥损失,提高了氮肥利用率。目前,有关菘蓝营养生理的研究主要集中在氮素形态、施氮方式对其的影响[8-10],而关于低氮营养下菘蓝生长与活性成分积累响应的研究尚鲜见报道。本研究采用盆栽试验,研究不同供氮水平对菘蓝生物量积累、光合参数、碳氮代谢及药材质量的影响,以期阐明菘蓝对低氮环境的生理响应,为生产实践中科学合理施氮、获取优质药材提供理论依据。
1 材料与方法
1.1 试验材料
试验材料为来自山西的菘蓝栽培居群,经南京农业大学中药材研究所王康才教授鉴定为十字花科植物菘蓝(IsatisindigoticaFort.)的角果(生产上称为种子)。
1.2 试验设计
试验于2017年7-12月在南京农业大学温室内进行。采用育苗移栽方式,挑选籽粒饱满、大小均匀的种子播于穴盘中,出苗后每10 d浇施一次1/4 Hoagland营养液(氮浓度为3.75 mmol·L-1),每次500 mL。基础营养液中,大量元素与微量元素采用改良Hoagland营养液配方,基础营养液pH值 6.0。待幼苗真叶长至4~5片时,选择长势基本一致的植株,转移至盆栽(外口径29.6 cm,底直径 17.8 cm,高19.7 cm)中,栽培基质由蛭石和珍珠岩(蛭石∶珍珠岩=2∶1)混合而成。每盆定苗5株。各处理重复8次,共40盆,随机排列。播种40 d后开始处理,每隔7 d浇灌1次处理营养液,于2017年12月10日收获。
试验设5个氮素水平,分别为0(记作N0)、2.5(记作N1)、5.0(记作N2)、10.0(记作N3)、15.0 mmol·L-1(记作CK)。处理营养液均加入硝化抑制剂双氰胺,用量为处理液中纯氮含量的0.4%。不同处理中大量元素配比如表1所示。
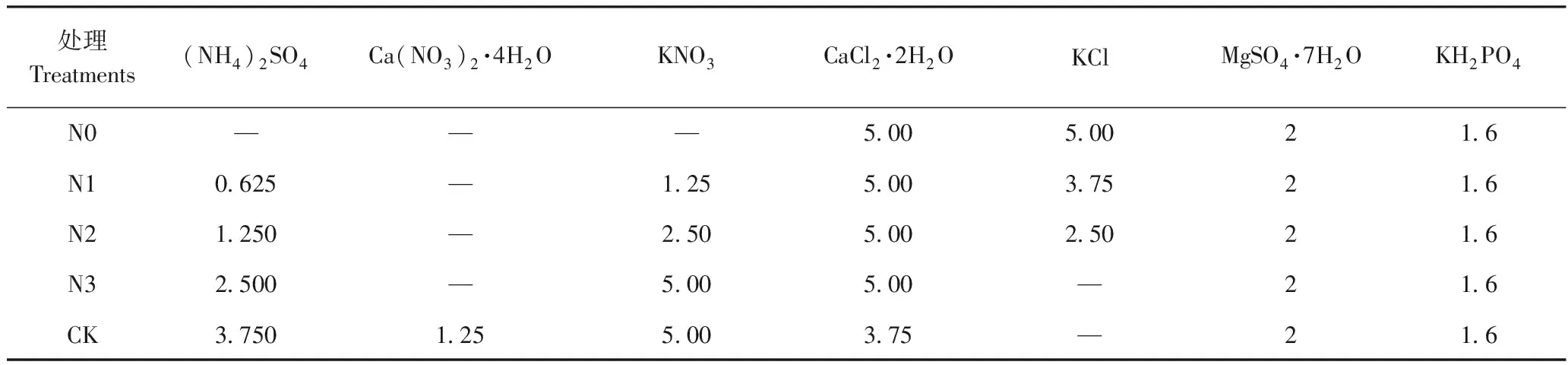
表1 不同处理中大量元素配比Table 1 The major elements ratio in different treatments /(mmol·L-1)
注:“—”表示处理营养液中不含列表中对应的化学成分。
Note: ‘—’ indicates that the chemical components in the nutrient solution are not included in the list.
1.3 测定指标及方法
1.3.1 生物量测定 各处理随机选取10株,清水洗净后用灭菌的滤纸吸干水分,将地上部分与地下部分分开,根和叶均经105℃杀青15 min,然后60℃烘干至恒重,分别称量单株根与叶的干重。并将叶、根的干样分别粉碎后过60目筛,备用。按照公式计算根冠比:
根冠比=单株根干质量/单株叶干质量
(1)。
1.3.2 游离氨基酸含量的测定 采用茚三酮显色法[11]。分别称取菘蓝根、叶的干样各0.1 g于试管中,每处理3次重复,加入10 mL蒸馏水,然后沸水浴20 min,冷却后过滤,保留上清液。向沉淀中加入5 mL蒸馏水,然后沸水浴10 min,过滤并反复冲洗残渣,合并2次的上清液并定容至25 mL。取4支洁净干燥的试管,其中3支分别加入0.5 mL稀释过的样品提取液,1支加入0.5 mL蒸馏水,分别在上述4支试管中加入醋酸缓冲液、水合茚三酮各0.5 mL,混匀后盖塞并沸水浴12 min,冷却后再分别加入5 mL 95%乙醇,摇匀。以空白作参比,于波长570 nm下测定吸光度值,并计算游离氨基酸含量。
1.3.3 硝态氮含量的测定 采用水杨酸-硫酸法[11]。样品提取步骤与游离氨基酸相同。吸取0.1 mL样品液,加入0.4 mL 5%水杨酸-硫酸溶液,混匀后于室温下放置20 min,缓慢加入9.5 mL 8% NaOH溶液,待冷却至室温后,以空白作参比,于410 nm波长下测定其吸光度值。用标准曲线上计算出0.1 mL样品提取液中硝态氮含量,再计算样品中的硝态氮含量。
1.3.4 可溶性糖含量的测定 采用蒽酮比色法[11]。样品提取步骤与游离氨基酸相同。吸取稀释过的样品提取液0.5 mL于20 mL刻度试管中,重复3次,加入1.5 mL蒸馏水。然后加入0.5 mL蒽酮乙酸乙酯试剂,再缓慢加入5 mL浓硫酸,充分振荡后立即放入沸水浴中准确保温1 min,(比色空白用2 mL蒸馏水与0.5 mL蒽酮试剂混合,并一同于沸水浴保温)。取出后冷却至室温,以空白作参比,在630 nm波长下测定其吸光度值,并计算可溶性糖含量。
1.3.5 光合参数的测定 于晴朗、阳光充足的天气,选择菘蓝植株由内至外的第3片功能叶,于2017年12月9日上午9:00-11:00进行光合参数测定。采用 LI-6400便携式光合作用测量系统(LI-COR公司,美国)测定叶片的净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、胞间CO2浓度(intercellular CO2concentration,Ci)和蒸腾速率(transpiration rate,Tr)。测定时使用开放式气路,采用红蓝光源,光通量密度为1 000 μmol·m-2·s-1。
1.3.6 叶中靛蓝和靛玉红含量的测定 参照2015年版《中华人民共和国药典》[4]的方法提取菘蓝叶中的靛蓝、靛玉红,应用超高效液相色谱法测定其含量。色谱条件:Agilent 超高效液相色谱仪。流动相为v(甲醇)∶v(水)=72∶28;流速0.300 mL·min-1;检测波长为289 nm;柱温30℃;进样体积2 μL。采用外标法计算叶中靛蓝和靛玉红含量。
标准曲线绘制:准确称取靛蓝和靛玉红标准品各1.0 mg,用甲醇溶解后均定容至50 mL,摇匀后用孔径0.45 μm微孔滤膜过滤。临用前,用甲醇溶液采用逐级稀释法将标准品母液配制成一系列质量浓度的单标标准品溶液。分别吸取靛蓝、靛玉红标准品溶液各2 μL进行测定。以色谱峰面积为纵坐标、浓度为横坐标进行标准曲线拟合。靛蓝标准曲线为y=16.022x+21.742,r2=0.999 4(n=3),线性范围0~2 μg·mL-1;靛玉红标准曲线为y=157.32x-9.329 4,r2=0.999 8(n=3),线性范围0~10 μg·mL-1。
供试品溶液制备:准确称取0.25 g叶粉末于索氏提取器中,加入适量三氯甲烷,浸泡15 h,加热回流提取至提取液无色。回收溶剂蒸干,残渣加甲醇使其溶解,然后转移至100 mL容量瓶中,补充甲醇至刻度,摇匀,过滤,即得供试品溶液。
1.3.7 根中(R,S)-告依春含量的测定 参照2015年版《中华人民共和国药典》[4]的方法提取菘蓝根中的(R,S)-告依春,并采用高效液相色谱法测定其含量。色谱条件:HITACHI D2000高效液相色谱系统(日立公司,日本)。流动相为 v(甲醇)∶v(0.02%磷酸溶液)=20∶80;流速0.600 mL·min-1;检测波长245 nm;柱温30℃;进样体积20 μL。采用外标法计算根中(R,S)-告依春含量。
标准曲线绘制:准确称取1.0 mg(R,S)-告依春标准品,用甲醇溶解并定容至25 mL,配制成40 μg·mL-1标准品母液。微孔滤膜(0.45 μm)滤过,备用。临用前,用甲醇溶液采用逐级稀释法将标准品母液配制成浓度分别为1.0、2.5、5.0、10.0、20.0、40.0 μg·mL-1的(R,S)-告依春标准品溶液,分别吸取20 μL进行测定。以色谱峰面积为纵坐标、浓度为横坐标进行标准曲线拟合。(R,S)-告依春标准曲线为y=116.93x-10.091,r2=0.999 8(n=3),线性范围0~40 μg·mL-1。
供试品溶液制备:准确称取0.5 g根粉末于圆底瓶中,加入25 mL蒸馏水,称定重量,煎煮2 h,待冷却后称定其重量,用水补足减失的重量,摇匀,滤过,取续滤液,即得供试品溶液。
1.3.8 叶中总黄酮含量的测定 准确称取0.100 0 g菘蓝叶粉末,加入10 mL 70%乙醇,超声波振荡1 h后过滤,将滤液定容至25 mL,混匀。吸取2 mL滤液于试管内,加入0.5 mL 50 g·L-1NaNO2,摇匀后静置6 min,再加入0.5 mL 100 g·L-1Al(NO3)3,摇匀后静置6 min,然后加入4 mL 40 g·L-1NaOH,最后用70%乙醇定容至10 mL。摇匀后静置15 min,于510 nm波长下比色,测定吸光度值。以芦丁为标准品绘制得到标准曲线为y=0.003 7x+0.002 2,r2=0.999 5(n=3),线性范围0~40 μg·mL-1,计算叶中总黄酮含量。
1.3.9 数据处理与分析 采用 Microsoft Office Excel 2013、SPSS 20.0进行数据统计分析,多重比较采用Duncan′s新复极差法。
2 结果与分析
2.1 不同供氮水平对菘蓝生物量积累的影响
由表2可知,随着氮素浓度的增加,菘蓝叶干重逐渐增加,其中CK显著高于其他处理(P<0.05)。根干重随着氮素浓度的增加而增加,但N0与N1之间、N3与CK之间均无显著差异(P>0.05)。单株干重的变化规律与叶、根干重的变化趋势基本一致,N0、N1、N2、N3处理的单株干重与CK相比,分别显著降低了91.0%、82.4%、52.8%、14.6%。根冠比随着氮素浓度的增加呈先增加后减少的趋势,依次表现为N1>N2>N3>N0>CK。表明在低氮条件下,菘蓝根与叶的生长均受到一定程度的抑制,氮素浓度降低到一定水平时促使植株的根冠比增加,对菘蓝地下部分生长的促进作用大于地上部分。
2.2 不同供氮水平对菘蓝可溶性糖、游离氨基酸、硝态氮含量的影响
由表3可知,N0处理下的菘蓝叶中可溶性糖含量最高,且显著高于其他处理(P<0.05),随着氮素浓度的增加,叶中可溶性糖含量呈先减少后增加的趋势,而根中可溶性糖含量的变化规律与叶中存在差异;在N1~CK范围内,根中可溶性糖含量呈先增加后减少的趋势,N2处理下根中可溶性糖含量显著高于其他处理(P<0.05),N0与N3之间无显著差异。叶中游离氨基酸含量随着氮素浓度的增加呈先增加后减少的趋势,在N3处理下达到最大值,N3与CK间无显著差异(P>0.05);根中游离氨基酸含量变化趋势与叶中相同,于N3处理下达到最大值。叶中硝态氮含量随着氮素浓度的增加呈先减少后增加的趋势,其中N3处理的硝态氮含量显著低于其他处理;根中硝态氮含量变化趋势与叶中一致,但同一氮素浓度下,叶中硝态氮含量均高于根。

表2 不同供氮水平对菘蓝根与叶干重及根冠比的影响Table 2 Effect of different nitrogen levels on dry weight and root to shoot ratio of I. indigotica
注:同列不同字母表示各处理间在5%水平差异显著。下同。
Note: Different letters in the same column indicate significant difference among different treatments at 0.05 level. The same as following.
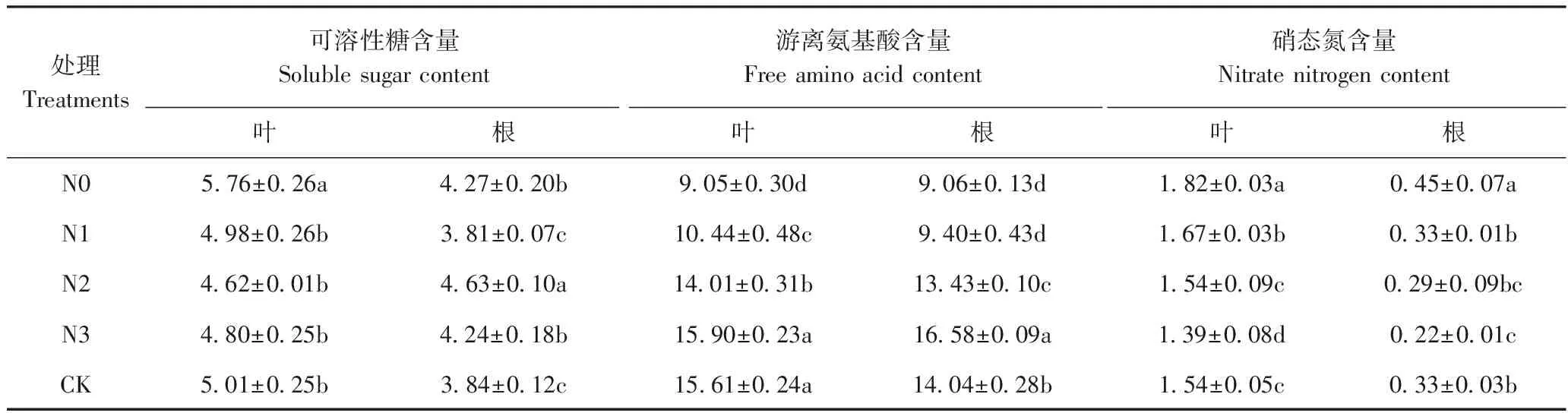
表3 不同供氮水平对菘蓝根与叶中可溶性糖、游离氨基酸、硝态氮含量的影响Table 3 Effect of different nitrogen levels on contents of soluble sugar, free amino acid and nitrate nitrogen in leaves and roots of I. indigotica /(mg·g-1DW)
2.3 不同供氮水平对菘蓝光合参数的影响
由图1可知,随着氮素浓度的增加,菘蓝叶片的Pn呈先升高后降低的趋势,N2、N3、CK间差异不显著,但均显著高于N0、N1(P<0.05);Gs也呈先升高后降低的趋势;叶片Tr的变化规律与Gs一致;而Ci则呈逐渐降低的趋势。表明氮素浓度过低不利于菘蓝植株进行光合作用。
2.4 不同供氮水平对菘蓝活性成分的影响
由表4可知,不同供氮水平对菘蓝叶中靛蓝、靛玉红含量的影响存在差异。靛蓝含量随着氮素浓度的增加逐渐增加,N3和 CK下的靛蓝含量均显著高于其他处理(P<0.05),但二者间无显著差异。靛玉红含量则随着氮素的浓度增加呈先减少后增加的趋势,低氮条件下的靛玉红含量较高。根中(R,S)-告依春的含量随着氮素浓度的增加呈先增加后减少的趋势,在N3处理下达到最高。叶中总黄酮含量随着氮素浓度的增加而逐渐减少,N0处理下达到最高,N0与N1间无显著差异(P>0.05),但二者的总黄酮含量均显著高于其他处理(P<0.05),且N2、N3、CK间无显著差异(P>0.05)。
由表5可知,随着氮浓度的增加,叶中靛蓝、靛玉红、总黄酮单株产量均呈上升的趋势,且均在CK处理达到最高。与CK相比,N3处理下的靛蓝、靛玉红、总黄酮单株产量分别下降了23.4%、20.4%、15.4%;N0处理下的靛蓝、靛玉红、总黄酮单株产量分别下降了97.5%、89.7%、90.5%。根中(R,S)-告依春单株产量则随着氮浓度的增加呈先增加再减少的趋势,在N3处理下达到最高,与CK相比,N3处理的(R,S)-告依春单株产量增加了43.2%。
2.5 菘蓝生物量与活性成分指标间相关性分析
对各处理的菘蓝生物量与活性成分指标间进行Pearson 相关性分析。由表6可知,叶干重与根干重呈极显著正相关,相关系数为0.991。叶干重与总黄酮含量呈极显著负相关,相关系数为-0.990。靛蓝含量与靛玉红、总黄酮含量之间均呈极显著负相关,相关系数分别为-0.974、-0.969。综上表明,氮素对于不同活性成分的调控特点存在差异。

注:不同小写字母表示差异显著(P<0.05)。Note: Different lowercase letters indicate significant different at 0.05 level.图1 不同供氮水平对菘蓝光合参数的影响Fig.1 Effect of different nitrogen levels on photosynthetic parameters of I. indigotica
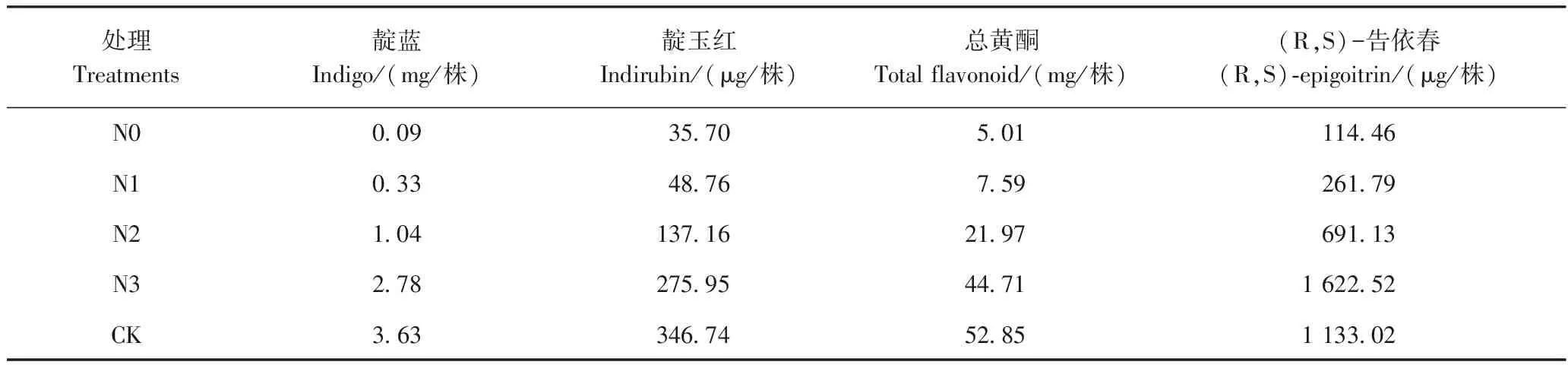
表5 不同供氮水平下菘蓝主要活性成分的单株产量Table 5 The yield per plant of the main active components under different nitrogen levels
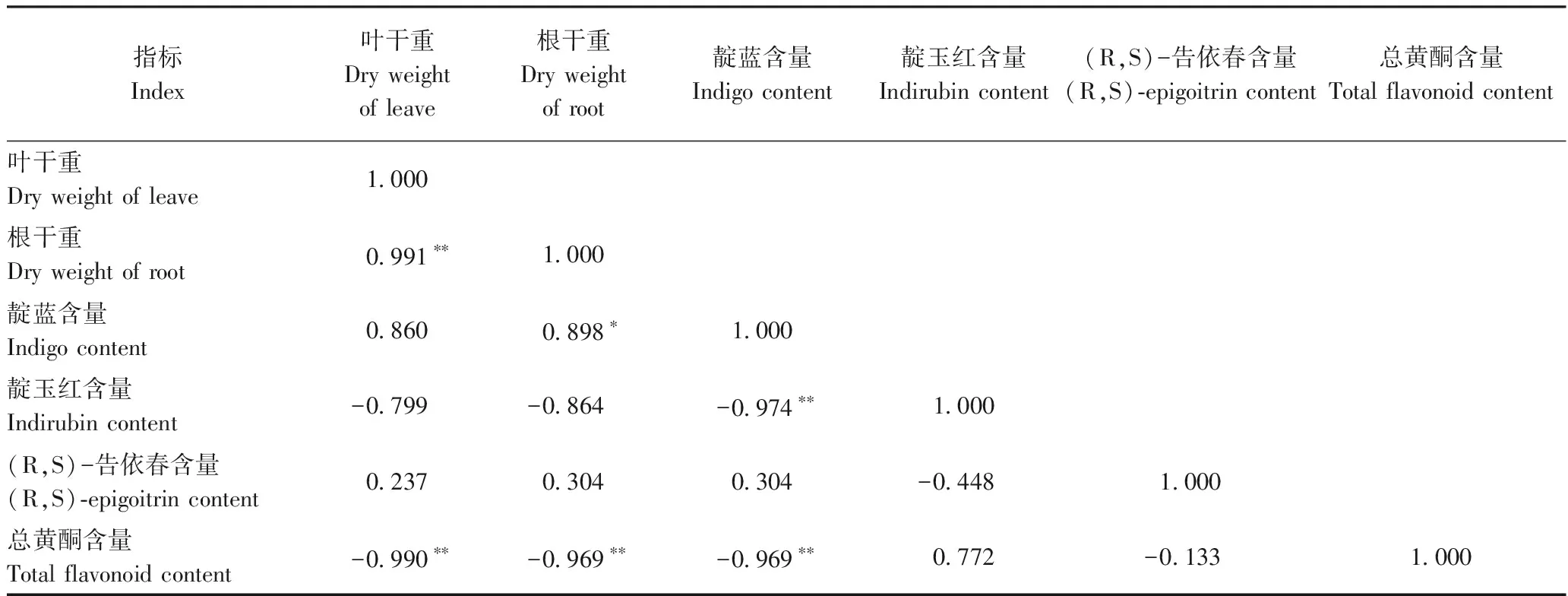
表6 菘蓝生物量与活性成分指标间相关性分析Table 6 Correlation analysis among biomasses and active components indexes in I. indigotica under different nitrogen levels
注:*表示在0.05水平相关性显著;**表示在0.01水平相关性极显著。
Note:*indicates correlation is significant at 0.05 level.**indicates correlation is extremely significent.
3 讨论
3.1 菘蓝生长对不同供氮水平的响应
氮素是植物生长与代谢过程中的重要影响因子,氮素不足时植株矮小且生长缓慢,氮素充足时植株生长较为旺盛,但氮素过量会对生长产生不利影响[12]。研究表明,氮素对植物生物量积累和产量形成具有促进效应。张云风等[13]研究发现随着氮浓度的增大,拟巫山淫羊藿的单株生物量和叶干重均显著升高。卢丽兰等[14]研究表明,高氮有利于广藿香生长,可显著提高其单株重量,这与本研究结果相同。本研究中,菘蓝根与叶的干重均随着氮素浓度的增加而增加,且CK的叶干重显著高于其他处理,但CK的根鲜重、根干重与N3之间无明显差异,表明适当减少氮素供给,对菘蓝根生物量积累影响较小,但氮素供应过少会抑制根与叶的生物量积累。本研究还发现菘蓝地上部分与根系在低氮营养下的响应存在差异,其根冠比随着氮素浓度增加呈先升高后降低的趋势,在N1处理下达到最大值。这是由于在氮素供应水平较低时,植物分配至地下部分的光合产物比例增加,相对提高根系生物量来适应环境变化,以获取更多养分,从而增大根冠比。低氮条件下,植株根冠比增加可能属于植物耐氮素胁迫的机制之一。
3.2 菘蓝碳氮代谢对不同供氮水平的响应
供氮水平对于植物体的碳氮代谢也存在显著影响。孙兴祥等[15]研究表明,菠菜中的可溶性糖含量随氮素浓度的降低呈上升趋势;张梅等[16]研究发现甜菜生长发育前期,随着施氮量的增加,叶片可溶性糖积累量整体上呈先降低后升高的趋势,中期逐渐下降,这与本研究结果相似。本研究中,菘蓝叶的可溶性糖含量随着氮素浓度的增加而呈先减少后增加的趋势,在N0处理达到最大值,根中可溶性糖含量在2.5~15.0 mmol·L-1范围内呈先升高后降低的趋势。这可能是由于氮素浓度较高时,植株代谢较旺盛,大量可溶性糖转化为碳架,导致可溶性糖含量下降。硝态氮和氨基酸含量在一定程度上可反映植物的氮素代谢情况。前人研究发现,低氮胁迫下,大豆叶与根系中硝态氮、游离氨基酸含量均低于正常氮素处理[17],这与本研究结果相似。本研究中,菘蓝叶与根中游离氨基酸含量变化趋势一致,均随着氮素浓度的增加而呈先增加后减少的趋势,在N3处理下达到最大值;菘蓝叶与根中的硝态氮含量则表现出先减少后增加的趋势,N3处理的硝态氮含量显著低于其他处理。氮素浓度较大时硝态氮含量低,可能是由于氮素浓度高,植物体内氮代谢关键酶活性较高,促进了无机氮向有机氮的转化,使得硝态氮的还原量大于吸收量。而在低氮条件下,氮代谢酶活性较低,硝态氮积累相对较多,导致其含量升高。同一供氮水平下叶中硝态氮含量高于根中硝态氮含量,可能是由于被吸收的硝态氮部分被根系直接转化,大量硝态氮被转移至叶片中。
3.3 菘蓝光合参数对不同供氮水平的响应
供氮水平对于植物的光合作用有较大影响。匡鹤凌等[18]研究表明,过低的氮素水平会导致喜树幼苗光合作用能力下降;李强等[19]研究发现在低氮胁迫下,苗期玉米叶片的Pn、Gs、Tr显著降低,Ci显著增加。本研究中,菘蓝的Pn、Tr、Gs均随着氮素浓度的增加呈先升高后降低的趋势,Ci呈逐渐降低趋势,这与许楠等[20]的研究结果类似。植物碳代谢和氮代谢相互影响与制约,氮素供应水平过高导致光合能力下降,可能是由于植物碳代谢与氮代谢均需要同化力及碳架。植物中氮素含量升高使氮同化作用增强,与光合碳同化竞争同化力,导致CO2同化速率降低。此外,氮同化加强后,呼吸作用向光合碳同化提供碳架的能力减弱,使CO2同化速率下降[17]。植物光合作用属于碳代谢的一部分,与同化产物在植物体内的分配与积累过程有一定内在联系。当植物氮素缺乏时,叶片Pn较低,叶中可溶性糖含量积累较多;在氮素充足时,植物向叶片投入较多氮营养,促进光合能力的提高,植物对于营养的需求增加,进而向根系分配较多光合产物以促进根系生长,增强根系对氮素的吸收,植株氮代谢活动旺盛,硝态氮含量降低。影响植物光合作用的主要因素为气孔限制和非气孔限制,叶片Pn、Gs和Ci同时下降,说明Pn下降主要因为气孔因素,而Pn、Gs下降、Ci升高说明非气孔限制是Pn降低的主要限制因素[21]。本研究中,N0~N3范围内,随着氮浓度的减小,菘蓝Pn、Gs、Tr均降低,Ci呈升高趋势,Pn与Tr、Gs均呈极显著正相关,与Ci间呈显著负相关,说明菘蓝光合作用主要受非气孔因素限制。
3.4 菘蓝活性成分对不同供氮水平的响应
药用植物的药效成分大多数为次生代谢产物。次生代谢产物是植物在与外界环境相互作用的过程中逐渐形成的,其形成除与遗传因素有关外,主要还受到生态因子的影响[22]。氮素可能通过影响植物中的碳氮代谢,也可能通过影响次生代谢过程的酶系统等方面,来调控植物的次生代谢途径,影响次生代谢产物含量[23]。兼顾药用植物产量及品质是生产实践中的关键环节,施用氮肥应考虑菘蓝生长与其次生代谢产物积累的平衡。
大多数生物碱属于含氮化合物,其代谢途径较为复杂。研究发现氮素对于生物碱的生物合成与积累有促进作用。丁丽洁等[24]研究表明,在一定氮素浓度范围内,黄檗中小檗碱、药根碱和掌叶防己碱均随着氮素浓度增加而增加。朱孟炎等[25]研究表明,低氮水平下,长春花叶片中的文朵灵、长春质碱、长春碱含量均显著低于正常和高氮水平。本研究中,菘蓝叶中靛蓝含量随着氮素浓度的增加逐渐增加,根中(R,S)-告依春含量随着氮素浓度的呈先增后减少的趋势,在N3处理下达到最高。但也有研究表明,氮素胁迫条件刺激了植物体内某些次生代谢物质的合成,使部分生物碱成分含量提高。孙世芹等[26]研究发现适当低氮胁迫能够显著增加喜树幼苗中的喜树碱含量。本研究中,菘蓝叶中靛玉红含量随着氮素浓度的增加表现出先减少后增加的趋势。靛玉红和靛蓝属于吲哚类生物碱,(R,S)-告依春为含硫类生物碱,这3类成分均为含氮次生代谢物。适当的氮素供应有利于菘蓝中靛蓝与(R,S)-告依春的积累,而低氮条件促进了靛玉红含量增加。根中(R,S)-告依春单株产量在N3处理下达到最高,表明适当降低施氮量,可以获得活性成分含量较高的板蓝根。通过调节氮素水平可以调控药用植物部分次生代谢产物的水平,为生产上获得高品质药材提供了可能的途径。
氮素对黄酮类化合物的合成代谢过程也有一定调控作用。菘蓝叶中总黄酮含量随着氮素浓度的降低呈逐渐增加的趋势,0~5 mmol·L-1氮浓度范围内的总黄酮含量显著高于其他处理。研究表明,在低氮环境下,黄酮与异黄酮合成相关基因的转录水平升高,可以通过氮素来调控黄酮类化合物的含量[27]。Løvdal等[28]研究发现在低氮条件下,番茄叶片中黄酮成分槲皮素、山奈酚及鼠李素的含量均有所升高,这与本研究结果相似。Liu等[29]研究表明,缺氮可使菊花叶片的黄酮含量保持在较高水平;向达兵等[30]研究发现施氮会显著降低苦荞麦中的芦丁含量,但对槲皮素含量无显著影响。上述结果表明,降低氮素浓度有利于植物中黄酮类成分的合成。在低氮条件下,不同植物中生物碱与黄酮类成分的响应存在差异,且菘蓝中靛蓝含量与靛玉红、总黄酮含量之间均呈极显著负相关,这可能与植物自身营养特性有关,也可能与次生代谢产物复杂的合成过程有关,但相关机制还有待进一步研究。
4 结论
本研究结果表明,氮素供应水平对菘蓝生长及药材质量有显著影响。氮素浓度在0~5 mmol·L-1范围内,菘蓝根与叶生物量积累及光合作用均受到一定程度的抑制。适当的氮素供应有利于菘蓝中靛蓝与(R,S)-告依春积累,而降低氮素浓度有利于增加靛玉红与总黄酮含量。根中(R,S)-告依春单株产量在N3(10 mmol·L-1)处理下达到最高,可以通过适当降低氮素浓度获得活性成分含量较高的板蓝根。在实际田间栽培生产中,为获得产量稳定、品质较好的药材,可以考虑适当降低氮肥用量,调控菘蓝部分次生代谢产物的水平,以获得高品质药材,但具体减量施氮措施还有待进一步研究。
猜你喜欢
中国民族民间医药(2023年18期)2023-10-28
广州化工(2020年20期)2020-11-02
广州化工(2016年11期)2016-09-02
环境污染与防治(2016年12期)2016-03-13
中国医药导报(2012年2期)2012-09-17
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年2期)2011-10-26
植物营养与肥料学报(2011年4期)2011-10-26
植物营养与肥料学报(2011年3期)2011-10-24
植物营养与肥料学报(2010年3期)2010-11-16
