
一种新型抗菌肽Hydramacin-1 在毕赤酵母中的重组表达、纯化及其抗菌活性
2019-08-22 01:55孟德梅石林李文娟孙雪晴郭雅君贾雪霞樊振川
天津科技大学学报 2019年4期
孟德梅,石林,李文娟,孙雪晴,郭雅君,贾雪霞,樊振川
(省部共建食品营养与安全国家重点实验室,食品营养与安全教育部重点实验室,天津科技大学食品工程与生物技术学院,大健康生物技术国际科技合作基地,天津市大健康生物技术国际联合研究中心,天津 300457)
抗菌肽(antimicrobial peptides,AMPs)是一种来源于动物、植物及真菌等先天免疫组织的有抑菌活性的小分子短肽[1],它可有效御防病原体入侵,大部分抗菌肽具有广谱抗菌活性和抑制细菌、真菌、寄生虫和病毒生长的功能,有些抗菌肽对肿瘤具有选择性杀伤作用[2-4].抗菌肽与传统抗生素相比,其作用机理独特,不易产生耐药性,因此成为了有望替代抗生素的理想药物[5].目前,基因工程法是生产抗菌肽的主要方法之一[6],其中利用毕赤酵母表达体系生产抗菌肽是一种重要方式.该表达体系相比于原核表达体系具有诸多优点[7-9],如可以将蛋白分泌到胞外,在蛋白质的加工、折叠、翻译后修饰等方面具有诸多优势,从而能较好地保持目的蛋白的生物活性.现在已有诸多来源的抗菌肽在毕赤酵母表达体系中成功表达,如人源抗菌肽human cathelicidin[10]、鸡源抗菌肽Fowlicidin-3[11]和植物源抗菌肽PaDef[12]等.
抗菌肽Hydramacin-1 来源于大乳头水螅的上皮防御组织,氨基酸序列与已发现的抗菌肽氨基酸序列具有极低的相似性,是一种新型抗菌肽[13].该抗菌肽含有60 个氨基酸残基,净电荷为+6,结构中具有一个二硫键桥.二硫键桥将该肽中的一个α-螺旋和β-折叠连接在一起,形成一个结扣式的结构,有利于维持该结构的稳定.疏水性氨基酸的比例为33%,分布在两侧,由于带正电荷的残基被夹在两个疏水区域的中间,疏水区域的基团与细胞膜上的疏水基团相互作用插到磷脂双分子层中,从而使得细胞聚集在一起,因此它的抑菌机制被描述为聚集效应模型.Jung等[13]鉴定了由大肠杆菌表达系统表达的重组抗菌肽Hydramacin-1 的抗菌活性,结果显示Hydramacin-1具有广谱抗菌特性,对革兰氏阴性菌具有较好的抑制效果,但对革兰氏阳性菌的抑制效果并不理想,其中对金黄色葡萄球菌的最小抑菌浓度高于100 μg/mL.因此,本文将抗菌肽Hydramacin-1 在毕赤酵母系统中表达,获得重组抗菌肽Hydramacin-1,并检测其表达产量和抗菌活性.
1 材料与方法
1.1 材料
1.1.1 菌株和质粒
毕赤酵母(Pichia pastoris)GS115 和质粒pPICZαA 购于Invitrogen 公司;大肠杆菌(Escherichia coli)O157 ATCC35150、大肠埃希氏菌(Escherichia coli)ATCC10305、单增李斯特氏菌(Listeria monocytogenes)ATCC21633 和金黄色葡萄球菌(Staphylococcus aureus)ATCC25923 均购于美国菌种保藏中心;枯草芽胞杆菌(Bacillus subtilis)LZZ-133 由中国农业大学申琳教授惠赠.
1.1.2 主要试剂
博来霉素(Zeocin),Invitrogen 公司;庆大霉素(Gen)、超低分子质量蛋白质marker 和BCA Protein Assay Kit,北京索莱宝科技有限公司;限制性核酸内切酶EcoRⅠ、KpnⅠ和Taq DNA 聚合酶,Fermentas公司;限制性核酸内切酶SacⅠ、dNTPs、1 kbp DNA Ladder、100 bp DNA Ladder 和DNA marker Trans 2 K,全式金生物技术有限公司;其他试剂均为国产分析纯.
1.2 方法
1.2.1 Hydramacin-1 重组表达载体的构建
依据毕赤酵母密码子的偏好性[14]优化抗菌肽Hydramacin-1 的DNA 序列,在Hydramacin-1 DNA序列的 5′端添加酶切位点 EcoRⅠ,起始密码子ATG,3′端添加终止密码子TAA,酶切位点KpnⅠ,最终顺序为:EcoRⅠ-ATG-Hydramacin-1 目的基因-TAA-KpnⅠ.将以上设计好的目的基因送往苏州金唯智生物科技有限公司进行全基因合成,并通过EcoRⅠ和KpnⅠ两个酶切位点将合成的目的基因克隆至载体pPICZαA 上,最终得到含有重组表达质粒pPICZαA-Hydramacin-1 的穿刺菌.
挑取含有重组表达质粒pPICZαA-Hydramacin-1的穿刺菌接种于含有 Zeocin(终质量浓度为25 μg/mL)的LB 培养基中,37 ℃、200 r/min 振荡培养过夜.使用北京索莱宝生物科技有限公司的质粒小提试剂盒提取质粒.提取的质粒用限制性核酸内切酶EcoRⅠ和KpnⅠ对其进行双酶切验证,并送往北京奥科生物公司测序验证.
1.2.2 重组抗菌肽Hydramacin-1 的诱导表达与鉴定
使用限制性核酸内切酶SacⅠ对重组表达质粒pPICZαA-Hydramacin-1 在37 ℃进行单酶切2 h,使其线性化.经DNA 产物回收试剂盒收集的线性化质粒与毕赤酵母GS115 感受态细胞混匀,置于预冷的2 mm 电转杯中,以电压1 500 V、电容25 μF、电阻200 Ω 的条件进行电击,最后将电转化产物涂布于含有Zeocin(终质量浓度为200 μg/mL)的YPD 平板上,30 ℃培养3~4 d,观察转化子的生长.
利用菌落PCR 验证的方法[15]进行阳性转化子的筛选.挑取不同酵母转化子于0.2% SDS 溶液中,混匀,沸水浴 10 min,使菌体细胞膜破裂释放出基因组DNA,12 000 r/min 离心10 min,取上清液作为实验组的PCR 模板;用未转入质粒的毕赤酵母GS115 单克隆作阴性对照,同样上述处理;用重组质粒pPICZαA-Hydramacin-1 作阳性对照.利用软件(Primer premier 5.0)设计的特异性引物 P1(5'-GAAGCTGTCATCGGTTACTCA-3')和 P2(5'-TCC GCACAAACGAAGGTC-3')进行PCR 验证.PCR 反应结束后,取5 μL PCR 产物进行0.8%琼脂糖凝胶电泳,检测抗菌肽基因是否成功插入到毕赤酵母基因组中.
挑取不同阳性转化子于5 mL BMGY 培养基中,28 ℃、220 r/min 培养18~24 h 后,按20%的比例转接于25 mL BMGY 培养基中培养;待A600达8.0~10.0 时,收集菌体,离心弃上清液,用甲醇体积分数为1.0%的25 mL BMMY 培养基重悬菌体,进行诱导表达;每隔24 h 补加甲醇,96 h 后,离心收集上清液进行Tricine-SDS-PAGE 凝胶电泳[16],检测目的蛋白是否表达.
1.2.3 表达条件的优化
将扩大培养后的重组酵母转化子离心收集,重悬于甲醇体积分数分别为 0.5%、1%、1.5%、2%的BMMY 培养基中培养,每隔24 h 取样,并补加甲醇至各自的终浓度,144 h 后终止培养,最后通过BCA Protein Assay Kit 测定发酵上清液中的总蛋白含量,同时进行Tricine-SDS-PAGE 凝胶电泳和银染实验检测目的蛋白,并结合ImageJ 1.8.0 软件对电泳结果进行灰度分析,比较不同发酵时间、不同甲醇体积分数对目的蛋白表达量的影响.
1.2.4 重组抗菌肽Hydramacin-1 的纯化
通过离子交换层析的方法,使用1 mL HiTrap SP柱(GE Healthcare,USA)纯化抗菌肽 Hydramacin-1.首先,将发酵上清液在4 ℃、8 000 g 的条件下离心10 min,去除菌体,并用0.22 μm 的水系滤器过滤发酵上清液;然后用3 个柱体积的50 mmol/L 磷酸盐缓冲液(pH 7.0)对发酵上清液透析置换.HiTrap SP 柱先用5 倍柱体积的含有1 mol/L NaCl 的50 mmol/L磷酸盐缓冲液(pH 7.0)洗涤,再用5 倍柱体积的50 mmol/L 磷酸盐缓冲液(pH 7.0)平衡.用注射器将样品以1 mL/min 的速度注入柱中,用5 倍柱体积的50 mmol/L 磷酸盐缓冲液(pH 7.0)漂洗;最后用5 倍柱体积的含有150 mmol/L NaCl 的50 mmol/L 磷酸盐缓冲液(pH 7.0)洗脱目的蛋白,通过Tricine-SDSPAGE 凝胶电泳和银染实验,结合ImageJ 1.8.0 软件进行灰度分析,检测纯化后的抗菌肽纯度.将纯化后的抗菌肽真空冷冻干燥,保存.
1.2.5 重组抗菌肽Hydramacin-1 的抑菌活力测定
通过测定抗菌肽质量浓度分别为30、60、80、100 μg/mL 时对各指示菌的抑制率,进而评价其抗菌活性的高低.挑取指示菌于1 mL LB 培养基中,培养至对数生长期,再用LB 培养基稀释菌体至1×105mL-1左右,待用.用无菌的50 mmol/L 磷酸盐缓冲液(pH 7.0)将抗菌肽溶解至所需浓度,将20 μL 的各浓度抗菌肽加入96 孔板中,再向每个孔中加入100 μL 稀释完毕的菌液,每个样品3 个平行,37 ℃过夜振荡培养16 h,通过酶标仪在600 nm 下测定样品的吸光度.实验组 A600值记为 A,20 μL 无菌的50 mmol/L 磷酸盐缓冲液(pH 7.0)和100 μL 的新鲜LB 培养基混合液吸光度为 A1,20 μL 无菌的50 mmol/L 磷酸盐缓冲液(pH 7.0)与100 μL 的受试菌混合液吸光度为A0,按照式(1)计算抑制率.

2 结果与分析
2.1 Hydramacin-1重组表达载体的构建
抗菌肽 Hydramacin-1 的 DNA 序列全长为180 bp,具有60 个氨基酸,根据毕赤酵母密码子的偏好性(http://www.kazusa.or.jp/codon/)对其基因序列进行优化设计(图1).
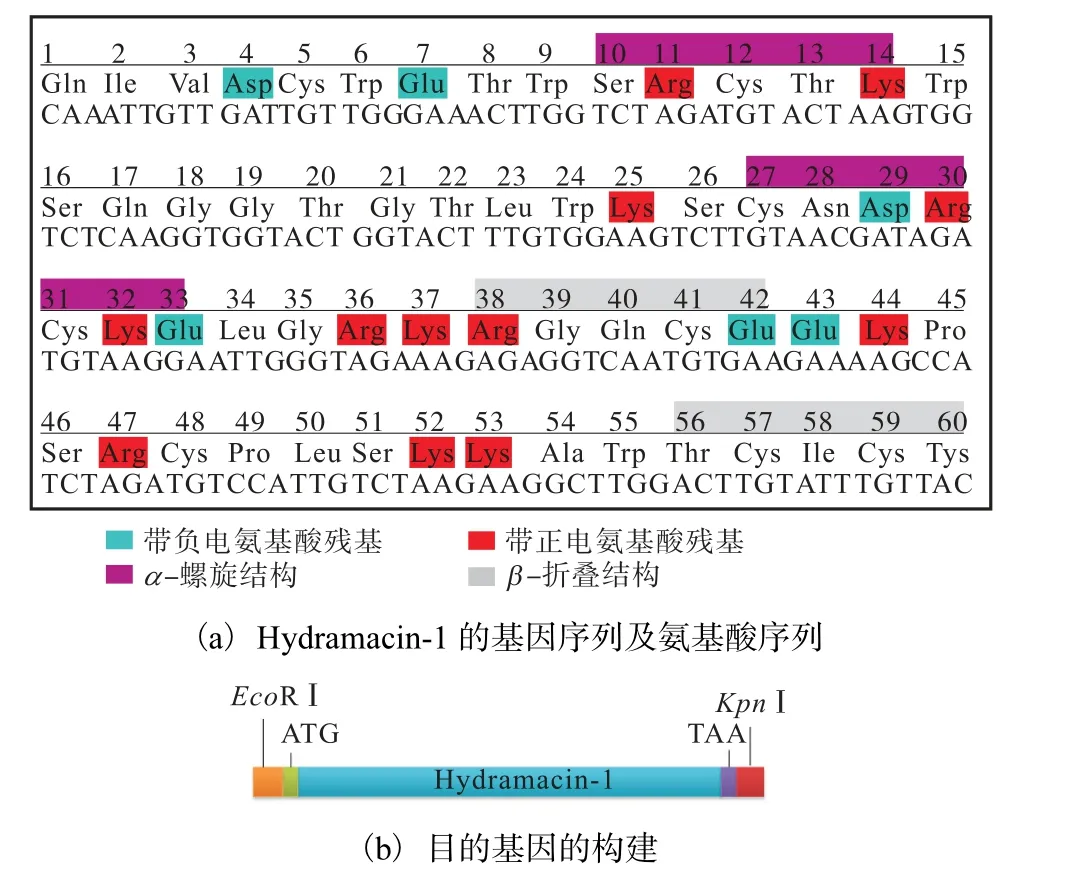

图1 pPICZαA-Hydramacin-1重组表达载体的构建Fig.1 Construction of P.pastoris expression plasmid pPICZαA-Hydramacin-1
优化后的DNA 序列如图1(a)所示;在优化后的DNA 序列前加上EcoRⅠ酶切位点和ATG 起始密码子,序列后加上TAA 终止密码子和KpnⅠ酶切位点,序列长度为198 bp,如图1(b)所示;再将其连接至毕赤酵母外泌型表达载体pPICZαA 中,如图1(c)所示.
用限制性核酸内切酶EcoRⅠ和KpnⅠ对获得的重组质粒进行双酶切,得到两条大小分别约为200 bp和3 500 bp 的条带,与理论上设计的载体长度一致(图2).此后将质粒送往北京奥科公司测序,测序结果同样证明重组表达质粒pPICZαA-Hydramacin-1 构建成功.

图2 pPICZαA-Hydramacin-1的双酶切验证Fig.2 Restriction identification of pPICZαA-Hydramacin-1
2.2 高表达量转化子的获得
采用菌落PCR 的方法进行阳性转化子的验证,结果如图3 所示.泳道1 为阴性对照,无条带出现,证明实验的可靠性;泳道2 为阳性对照,重组质粒作模板,扩增出750 bp 的单一目的条带;泳道3—9 为验证的7 个不同酵母转化子,其所出现的条带与阳性对照条带相一致,说明Hydramacin-1 基因已整合到毕赤酵母基因组中,它们均为阳性转化子.
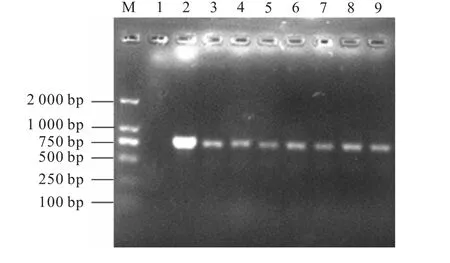
图3 菌落PCR筛选阳性转化子Fig.3 Positive transformant screening by colony-PCR
由于不同转化子中插入的目的基因片段数目不相同,导致拷贝数不同;拷贝数越高,表达量越高,因此进一步筛选获得的阳性转化子.取等体积的不同阳性转化子的发酵上清液进行Tricine-SDS-PAGE 凝胶电泳检测,并对不同转化子的目的蛋白条带进行灰度分析比较,结果如图4 所示.7 个阳性转化子表达的目的蛋白大小均在7.0×103左右,证明其得到了准确表达;通过对目的条带进行灰度分析比较,最终选取了蛋白表达量最高的6 号转化子进行后续实验研究.

图4 重组抗菌肽Hydramacin-1在毕赤酵母中的诱导表达Fig.4 Expressing recombinant Hydramacin-1 in P.pastoris
2.3 发酵时间和甲醇体积分数对蛋白表达的影响
2.3.1 诱导表达条件对总蛋白表达量的影响
由于实验中所用载体为外泌型表达载体,蛋白主要富集在发酵上清液之中,并且GS115 菌株分泌的自身蛋白量很低[17],所以通过BCA 法测定上清液中总蛋白的表达量情况,也可在一定程度上反映目的蛋白的表达量情况.诱导表达条件对总蛋白表达量的影响如图5 所示.由图5 可知:转化空白质粒的毕赤酵母在不同体积分数甲醇的诱导下,总蛋白表达水平在72 h 后趋于稳定,最高表达量小于100 μg/mL;而在4 个不同体积分数甲醇的诱导下,转化重组表达载体的阳性转化子的总蛋白表达水平均呈上升趋势,并在144 h 时达到最高,在甲醇体积分数为1.5%时,阳性转化子分泌的总蛋白量已达200 μg/mL,是转化空质粒毕赤酵母蛋白表达量的2 倍.

图5 诱导表达条件对总蛋白表达量的影响Fig.5 Effect of expression conditions on total protein expression
2.3.2 诱导表达条件对目的蛋白表达量的影响
通过Tricine SDS-PAGE 凝胶电泳结合灰度分析,在甲醇体积分数为1.5%时,目的蛋白表达量随诱导时间的延长而增加,在诱导时长为120 h 时,灰度分析值最大,目的蛋白表达量达到最高(图6).

图6 诱导表达时间对目的蛋白表达量的影响Fig.6 Expression of recombinant Hydramacin-1 at different induction time
在诱导表达时间为120 h 的条件下,目的蛋白表达量随甲醇体积分数的提高而增加,在甲醇体积分数为1.5%时,目的蛋白表达量达到最高(图7).
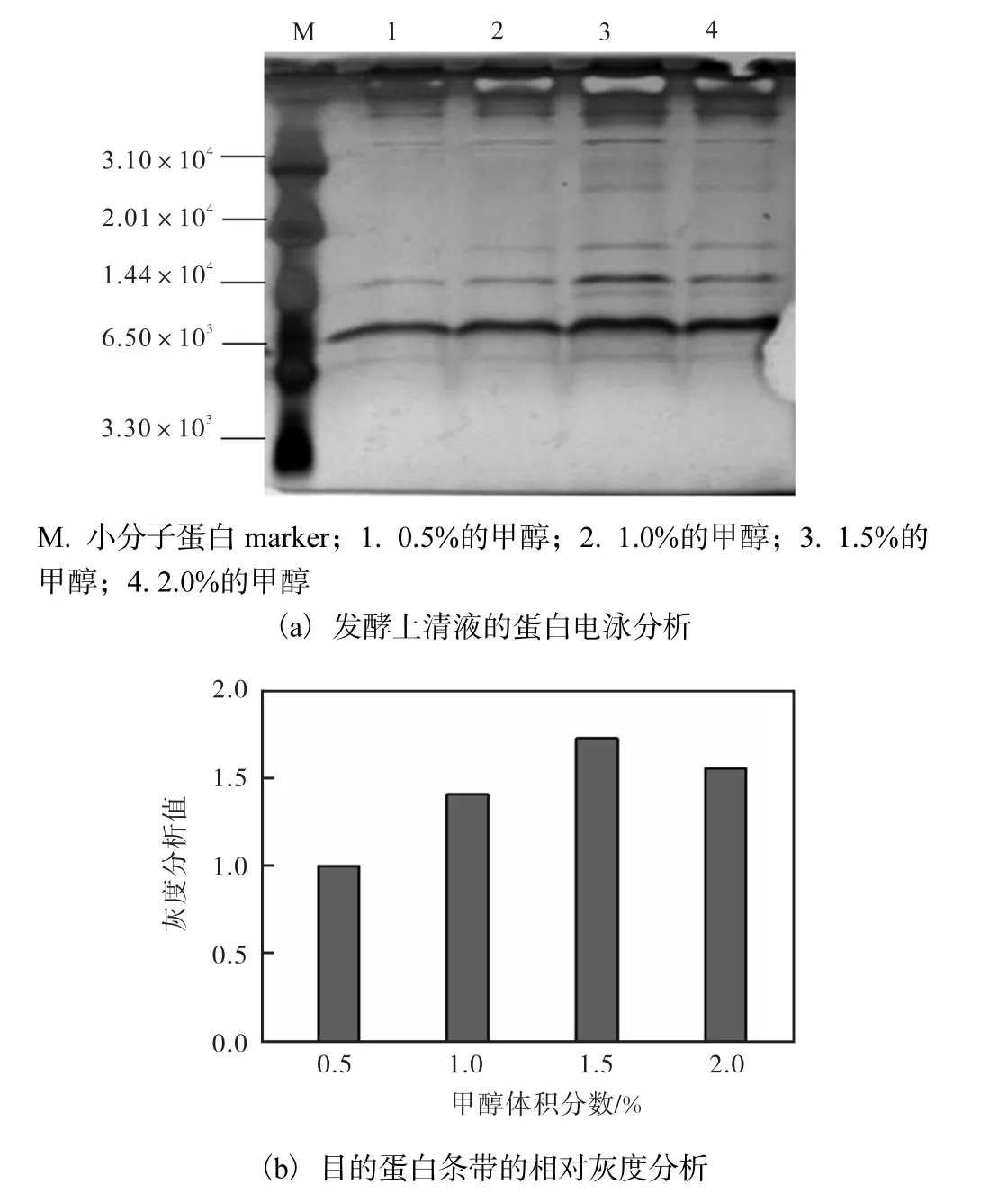
图7 甲醇体积分数对目的蛋白表达量的影响Fig.7 Expression of recombinant Hydramacin-1 induced with different methanol concentrations
因此,甲醇体积分数为1.5%,发酵时间为120 h为发酵最优条件,总蛋白表达量可达166 μg/mL.由灰度分析得知,目的蛋白表达量占总蛋白量的50%以上,由此可计算出目的蛋白的产量可达83 mg/L.
2.4 重组抗菌肽Hydramacin-1的纯化
通过HiTrap SP 柱对发酵上清液进行纯化,得到了纯度较高的目的蛋白,如图8 所示.

图8 阳离子交换柱纯化重组抗菌肽Hydramacin-1Fig.8 Cation exchange column purified recombinant Hydramacin-1
泳道1 为流穿液,杂蛋白流出,目的蛋白仅有微小损失;泳道2 为漂洗液,未见目的蛋白被洗下来,可见漂洗液的离子强度与pH 条件选择正确;泳道3—5 为洗脱液对目的蛋白进行洗脱的结果,得到单一目的条带,经灰度分析,蛋白纯度达90%以上.
2.5 重组抗菌肽Hydramacin-1的抑菌活力测定
使用纯化后的目的蛋白进行抑菌活力测定,结果如图9 所示.由图9 可知:重组抗菌肽Hydramacin-1具有广谱抗菌特性,8 0 μ g/m L 的重组抗菌肽Hydramacin-1 可将大肠杆菌O157 ATCC35150 全部抑制,抑制率达100%;100 μg/mL 的重组抗菌肽Hydramacin-1 对大肠埃希氏菌ATCC10305 的抑制率为100%,而对革兰氏阳性菌单增李斯特氏菌ATCC 21633、金黄色葡萄球菌ATCC25923 和枯草芽胞杆菌LZZ-133 的抑制率仅分别为38%、72%和64%.由此可见,相比于革兰氏阳性菌,重组抗菌肽Hydramacin-1 对革兰氏阴性菌具有更好的抑制作用.

图9 不同蛋白质量浓度下的重组抗菌肽Hydramacin-1对指示菌的抑制率Fig.9 Inhibition rate of the purified recombinant Hydramacin-1 with different concentration against the tested bacterial strains
3 讨 论
抗菌肽来源广泛,稳定性好,并且不易产生耐药性,可应用于医药、食品、饲料等不同领域,因此抗菌肽成为了当前研究的热点.例如,由于Nisin[18]可以很好地抑制单增李斯特菌和梭状肉毒杆菌的生长,进而有效保证食品安全,延长货架期,很多国家如美国、中国等均允许其应用到食品防腐保鲜上,尤其是应用于易被单增李斯特菌污染的乳制品上.本研究中的抗菌肽Hydramacin-1 若直接从生物体Hydra magnipapillata 中分离制备,存在着成本高、效率低、分离难度大等诸多问题.为解决这个问题,本实验选用毕赤酵母表达系统,制备了可高效表达重组抗菌肽Hydramacin-1 的基因工程菌株.通过多次实验以及对总蛋白表达量和目的蛋白表达量进行分析,120 h为最佳诱导表达时间.超过120 h,目的蛋白表达量降低,分析原因可能为发酵时间过长,目的蛋白发生降解,杂蛋白增多所致.此外,诱导所使用的甲醇体积分数也不宜过高,这是因为甲醇体积分数过高会对细胞造成毒害作用.本实验显示重组抗菌肽Hydramacin-1 在毕赤酵母中的最优表达条件是:培养温度为28 ℃,甲醇体积分数为1.5%,发酵时间为120 h.在此条件下,目的蛋白表达量可达83 mg/L.许多其他类型的抗菌肽,如 Mytichitin-A[19]、Scygonadin[20]和VpDef[21]也已在毕赤酵母中成功表达,并且在最佳培养条件下,这些重组肽的产量分别可达到45.5、70、60 mg/L.以上结果均可表明,毕赤酵母表达系统有利于重组抗菌肽Hydramacin-1 的高效表达.后续可通过发酵及进一步优化发酵条件的方式提高其产量.
重组抗菌肽Hydramacin-1 的高抗菌活力非常重要.本文测定了不同浓度下的重组抗菌肽Hydramacin-1 对5 株受试菌的抑制率,可知其具有广谱抗菌活性,对革兰氏阴性菌的抑菌效果明显优于革兰氏阳性菌,大肠杆菌O157 对其最为敏感.这一特点与大肠杆菌体系所表达的Hydramacin-1[22]的抗菌特点相一致.本研究中的重组抗菌肽Hydramacin-1 相比于Defensing-TK[23](对金黄色葡萄球菌的 MIC 为3 μg/mL)和rMP1106[24](对金黄色葡萄球菌的MIC为0.06 μg/mL)等抗菌肽的抗菌活性,还需进一步提高.目前主要是通过分子设计与结构改造的方法来提高肽的生物活性[25-26],例如氨基酸的替换,将一个或多个氨基酸经定点突变技术替换为精氨酸、赖氨酸等带正电的氨基酸,增加抗菌肽的带正电性,进而提高抗菌肽的生物活性.目前本实验室也正在对抗菌肽进行定向改造,以期获得活性更高的抗菌肽.
综上所述,本研究首次成功制备了具有高效表达能力和较好生物活性的重组抗菌肽Hydramacin-1 的毕赤酵母表达菌株,为后续抗菌肽Hydramacin-1 更深入的科学研究和应用奠定了基础.
猜你喜欢
现代食品(2022年5期)2022-12-13
江西农业大学学报(2022年4期)2022-10-08
农林经济管理学报(2022年4期)2022-10-08
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
江西农业学报(2021年4期)2021-04-20
渔业致富指南(2020年21期)2020-12-19
三农资讯半月报(2020年11期)2020-06-21
中国畜牧业(2019年22期)2019-12-30
中国酿造(2019年9期)2019-10-08
