
糯玉米自交系SSR标记遗传多样性及群体遗传结构分析
2019-01-17 03:22艾为大王义发李宏杨瞿玉玑沈雪芳
作物学报 2019年2期
卢 媛 艾为大 韩 晴 王义发 李宏杨 瞿玉玑 施 标,* 沈雪芳,*
糯玉米自交系SSR标记遗传多样性及群体遗传结构分析
卢 媛1,2艾为大3韩 晴1,2王义发1李宏杨4瞿玉玑5施 标1,2,*沈雪芳1,2,*
1上海市农业科学院作物育种栽培研究所, 上海 201403;2CIMMYT: 中国特用玉米研究中心, 上海 201403;3安顺学院农学院, 贵州安顺 561000;4三亚市南繁科学技术研究院, 海南三亚 572000;5上海市青浦区朱家角镇农业综合服务中心, 上海 201713
为了解糯玉米种质的遗传基础, 利用29对SSR标记对87份糯玉米自交系进行遗传多样性分析, 共检测出180个等位变异, 平均每个位点6个等位变异, 多态性信息含量变幅为0.308~0.915, 平均为0.572。材料间遗传相似系数为0.49~0.93, 平均为0.66。通过聚类分析UPGMA (unweighted pair-group method with arithmetic means)方法在遗传相似系数0.64处将87份糯玉米自交系划分为4个类群, 分别包含9、66、10和2份材料。此外, 利用Structure群体遗传结构分析也将87份糯玉米自交系分为4个类群, 分别包含24、25、19和19份材料; 进一步分析表明, 供试群体中大部分糯玉米自交系的遗传变异较单一。本研究为糯玉米新品种选育和遗传进化分析提供了种质基础和理论依据。
糯玉米; 遗传多样性; 群体遗传结构
玉米(L.)是全球三大重要作物之一, 糯玉米(L.var.Kulesh)是玉米属的一个亚种。糯玉米富含支链淀粉、营养丰富、适口性良好等优良特性赋予其宝贵的价值和广泛的用途, 现已成为深受欢迎的食品和工业原料, 具有较高的经济价值。
中国是公认的糯玉米起源中心[1]。曾孟潜(1987)论证认为早在1760年前, 中国已种植糯玉米[2]。目前的研究普遍认为玉米起源于美洲, 野生玉米被驯化为普通玉米后传入中国, 其第9染色体短臂上的基因发生隐性突变, 形成了一种籽粒干燥后胚乳呈角质不透明、无光泽的蜡质状突变体, 蒸煮后呈糯性, 因此, 被我国喜糯性食物的人们经人工选择保留下来, 称为糯玉米[3]。
虽然, 糯玉米起源于中国, 但是, 关于糯玉米种质资源方面的研究起步较晚, 对糯玉米种质资源的聚类分析主要集中在其遗传多样性。随着生物技术的快速发展, 糯玉米遗传多样性分析方法已突破了形态学、细胞学、同工酶标记方法的局限, 进入了分子生物学研究阶段。目前, 分子标记技术(RFLP、RAPD、SSR、SNP等)因其多态性高、稳定性好、操作简单等特点, 已被广泛应用于糯玉米的遗传多样性分析。刘丽君等[4]利用93对SSR标记研究了55份糯玉米自交系的遗传多样性, 基于UPGMA方法供试自交系分为4类, 划分结果基本符合品系的来源情况, 发现江苏沿江地区糯玉米种质资源主要由通系5群、衡白522群以及突变体材料组成。吴斌等[5]利用SSR标记基于UPGMA方法将36份西双版纳的小糯玉米地方品种聚为6个类群, 地方品种独立成群。Zheng等[6]利用SSR标记基于UPGMA方法将165份糯玉米材料聚类为3个类群。目前, 基于UPGMA方法对糯玉米种质的遗传多样性研究较多, 但是, 对基于模型的糯玉米群体遗传结构的分析还未见报道。本研究基于SSR标记对目前在育种中使用的87份糯玉米自交系进行了遗传多样性和群体遗传结构分析, 确定了糯玉米种质间的遗传背景差异, 为糯玉米优良品种选育和杂种优势模式构建提供了研究基础和理论依据。
1 材料与方法
1.1 试验材料
87份糯玉米自交系(N1~N87)和4份糯玉米杂交种(N88~N91)分别于2016年春、2016年秋和2017年春种植于上海市农业科学院庄行综合试验站(附表1), 行长4.0 m, 行距0.6 m, 株距0.25 m, 田间管理同一般生产田, 以上材料由上海市农业科学院玉米遗传育种课题组提供。
1.2 玉米基因组DNA提取
苗期于田间采集以上91份糯玉米材料幼嫩叶片,用液氮研磨后, 利用植物基因组DNA提取试剂盒以改进后的CTAB法提取DNA[7]。试剂盒购于北京庄盟国际生物基因科技有限公司。
1.3 SSR引物筛选与PCR扩增
根据中华人民共和国农业行业标准(NY/T1432- 2014玉米品种鉴定技术规程SSR标记法)提供的40对核心引物序列和PCR扩增程序对供试的91份材料进行基因型鉴定[8-9]。此外, 根据玉米基因组数据库(Maize Genome Database, https://maizegdb.org/)公布的玉米SSR引物信息, 在玉米第1~第10染色体上取定位于不同染色体bin的SSR标记各10个鉴定基因型, 采用Lu等[10]的PCR扩增和产物鉴定程序。
1.4 数据统计与分析

2 结果与分析
2.1 SSR标记分布及多态性分析
从140对SSR标记中共筛选出29对扩增稳定、条带差异明显的引物用于对87份糯玉米自交系的基因型分析, 共检测到180个等位基因, 变幅为4~20, 每个标记平均检测到6个等位基因(表1)。87份糯玉米自交系的平均基因多样性为0.619, 变幅为0.323~ 0.921。29对标记的PIC变幅为0.308~0.915, 平均为0.572; 其中, 20对标记的PIC ≥0.50 (69%), 属于高度多态性位点; 其余9对标记0.250 < PIC < 0.50 (31%), 属于中度多态性位点。综上结果说明这29对SSR标记多态性较高, 能较好地满足遗传多样性分析需求。

表1 29对SSR标记的多态性参数
2.2 遗传多样性分析
糯玉米单交种“沪紫黑糯2号”与亲本申W93间的GS为0.80; “五彩甜糯2号”与亲本申W93和wh-2间的GS分别为0.67和0.64; 16HW-LY5与其亲本16HW-31间的GS为0.63; 16HW-LY8与其亲本16HW-46间的GS为0.68。综上结果可见, 供试单交种与其亲本间的GS变幅为0.63~0.80, 亲缘关系较近, GS结果与系谱一致, 说明本研究对糯玉米种质间遗传多样性分析结果合理可靠。
16SW-292和16AW-363之间的GS最小(0.49), 16SW-466和16SW-468之间的GS最大(0.93)。根据系谱信息16SW-292和16AW-363选育于不同亲本, 亲缘关系较远; 16SW-466和16SW-468选自于相同亲本的不同组合, 亲缘关系很近(表1)。87份糯玉米自交系间两两计算共获得3741个GS, 变幅为0.49~0.93, 平均为0.66。由图1可见, 87份糯玉米自交系间的GS呈单峰分布, 85.40%的自交系间GS在0.60~0.80之间, 91.29%的自交系间GS ≤0.75, 说明本研究供试糯玉米自交系间遗传基础较丰富, 适于进行遗传多样性分析。
2.3 聚类分析
聚类树状图表明87份糯玉米自交系在GS为0.64处可划分为四大类(图2)。
类群I包括wh-2、京W♂和W97等来自北京和上海的共9份糯玉米自交系, 自交系间GS为0.59~0.87, 平均为0.69。根据系谱信息发现, 自交系京W♂、以及从以京W♂为亲本组配的杂交种中选育的二环系均被聚类于类群I, 京W♂与类群I内其他糯玉米自交系间的平均GS为0.69, 推测类群I材料与京W♂亲缘关系较近。
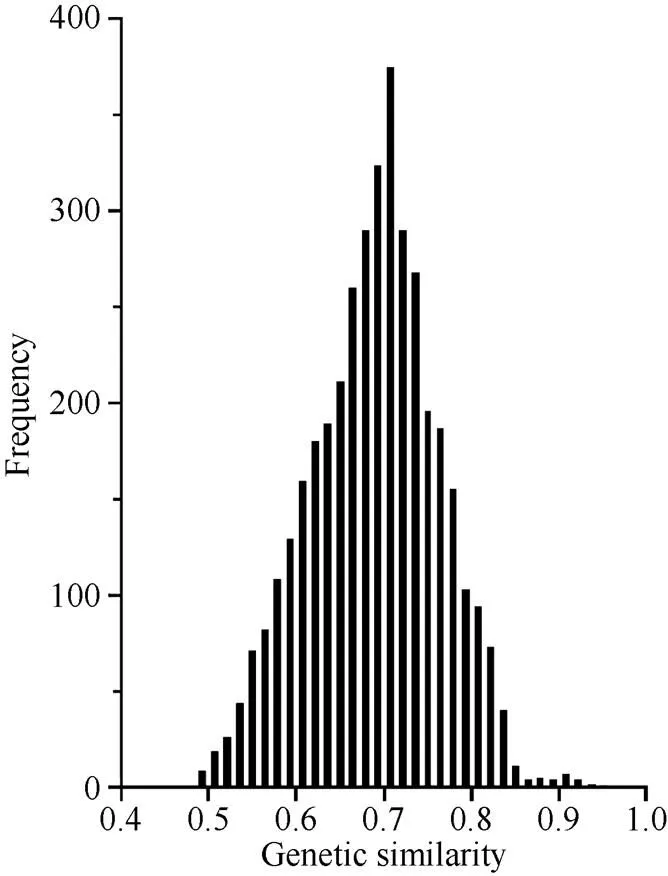
图1 糯玉米自交系间GS频次分布图
类群Ⅱ包括通系5、W158、09N-5M和16SW-365等66份糯玉米自交系, 材料间的GS为0.49~0.93, 平均为0.68。其中, 16SW-451和16SW-468间的GS最小, 为0.49; 16SW-466和16SW-468间的GS最大, 为0.93。此外, 类群II在GS为0.67处又可划分为5个亚类。亚群II-1包含通系5、W148和16SW-437等24份自交系, 它们之间的GS平均为0.72。其中, 包括糯玉米自交系通系5、以及以通系5为亲本杂交后选育的16SW-456、16SW-438、16SW-444等6份自交系, 通系5与亚群II-1内其他糯玉米自交系间的平均GS为0.69, 推测亚群II-1自交系与通系5亲缘关系较近。亚群II-2包括W151、W158、16SW-311等12份糯玉米自交系, 它们之间的GS平均为0.71; 其中除了来自河南的自交系16AW-1005外, 其余自交系均来源于上海和江苏。亚群II-3包括16SW-466、W-107、09N-5M等14份糯玉米自交系, 它们之间的GS平均为0.73; 其中除了来自河南的自交系16AW-1006外, 其余自交系均来源于上海和江苏。亚群II-4包括来自甘肃和上海的W116、16SW-322、16SW-46、16SW-333、16SW-427共5份糯玉米自交系, 它们之间的GS平均为0.65; 亚群II-4包括全部来自甘肃的自交系16SW-322、16SW-333和16HW-46, 它们与II-4亚群内来自上海的自交系W116和16SW-427间的平均GS分别为0.64、0.65和0.64, 推测W116和16SW-427与甘肃地区种质亲缘关系较近。亚群II-5包括黄W-♀、W153、16SW-365、16SW-373等来自广西、上海和广东的共11份糯玉米自交系, 它们之间的GS平均为0.71; 其中来自广西的16SW-327、16SW-365、16SW-373和16SW-374共4份糯玉米自交系, 它们与亚群II-5内其他自交系间的GS分别为0.67、0.73、0.74和0.72, 推测亚群II-5的自交系与广西糯玉米种质亲缘关系较近。
类群III包括09N-6M、W75、W152等10份糯玉米自交系, 它们之间的GS为0.57~0.85, 平均为0.70。类群III包括全部来自山东的自交系16SW-424、16SW-485和16HW-31, 它们与类群III其他自交系间的GS分别为0.75、0.75和0.71, 推测类群III自交系与山东地区种质亲缘关系较近。

图2 87份糯玉米自交系基于SSR标记的聚类分析图
来源于上海的糯玉米自交系16SW-321和申W93与其他材料间的GS平均分别为0.63和0.64, 遗传差异较大, 因此, 16SW-321和申W93被单独聚类为类群IV。
2.4 群体遗传结构分析
利用Structure软件分析发现, 在1~10范围内随着值的增大ln()值也随之增大(附图1)。因此, 依据Δ的方法进一步确定类群数, 发现在= 4时Δ值出现显著的峰值(图3-A), 据此推断87份糯玉米自交系可划分为4个类群, 分别包含24、25、19和19份糯玉米自交系(图3-B)。
利用自交系Q值可以分析不同类群在各自交系中所占成分比例。根据最大Q值分布, 87份糯玉米自交系中有15份Q值< 0.6, 占自交系总数的17.24%, 然而Q > 0.8和0.9的自交系分别占64.37%和47.13%, 说明该群体大部分材料的血缘关系较单一。

图3 87份糯玉米自交系的群体遗传分析图
A: Δ随值的变化趋势; B:= 4时, 以Structure运算获得的87份糯玉米自交系的群体遗传结构。
A: the line chart of Δchanging with-value; B: the population structure of 87 waxy maize inbred lines estimated by Structure (= 4).
3 讨论
3.1 糯玉米种质遗传多样性分析
一直以来, 搜集种质资源、拓宽遗传基础、对种质资源开展科学的鉴定与评价是选育优良品种的基础, 在品种改良中始终占据重要地位[14-15]。本研究利用29对SSR标记对87份目前育种中常用的糯玉米自交系进行遗传多样性分析, GS取值范围为0.49~0.93, 平均为0.66, 在GS为0.64处将供试群体划分为四大类。类群I包含9份糯玉米自交系, 其中包括京W♂、以及所有以京W♂为亲本选育的4份自交系。京W♂与类群I内其他糯玉米自交系间的平均GS为0.69, 亲缘关系较近, 推测16SW-122、W97、wh-2和16SW-292可能是北京种质京W♂在上海的改良自交系。
类群Ⅱ包含66份自交系, 包括“苏玉糯1号”的二环系16SW-437, 以“江南花糯”二环系为亲本选育的自交系16SW-296、16SW-324、16SW-363、16SW-369、16SW-438、16SW-452、16SW-459、16SW-465、16SW-466、16SW-468、16SW-471、16SW-476、16SW-487、16AW-1005、16AW-1008共15份材料, “美玉8号”的二环系16SW-365, “田白糯1号”的二环系16SW-458。“美玉8号”的母本是“苏玉糯1号”的二环系, 父本是衡白522与广西桂林农家种白糯玉米杂交选育的二环系[16]; “田白糯1号”母本是“苏玉糯2号”与“中糯1号”杂交选育的二环系, 父本选自广西河池农家种[17]; “苏玉糯2号”与“苏玉糯1号”的双亲都含有通系5和衡白522, 因此亲缘关系较近[18]。根据系谱信息, “美玉8号”、“田白糯1号”和“苏玉糯1号”的亲缘关系较近, 可见本研究结果与系谱信息一致。“苏玉糯1号”是由通系5与衡白522杂交育成的国审糯玉米品种, 具有高产、稳产性、品质较好、多抗的特点, 用其选育的二环系已在育种中大量使用, 组配出了“郑白糯4号”、“南农紫玉糯1号”、“山农202”等一批优质糯玉米品种[19]。“江南花糯”是由“苏玉糯1号”的二环系与农家种组配的杂交种, 具有产量高、肉质厚、糯性强、香味浓、外观美的特点[20]。类群II中16SW-460、16AW-1019和16AW-1005是以“郑白糯6号”的二环系为亲本选育的自交系。国审糯玉米品种“郑白糯6号”表现中矮秆、抗倒、抗病、产量高、结实性好、穗匀、品质好[21]。此外, 通系5和以通系5为亲本选育的12份自交系也被聚类于类群II。通系5农艺性状优良、糯性好、配合力高, 通系5或衍生品种已被国内多个育种单位作为种质资源广泛应用, 育成了28个优良的鲜食糯玉米品种。根据系谱信息可见, 类群II的自交系系谱来源品质优异, 推测类群II的自交系可作为骨干亲本用于今后的优质糯玉米选育。
类群IV只包含16SW-321和申W93两份材料, 其中申W93是“荆黑糯2号”的二环系。“荆黑糯2号”来源于湖北,据此推测类群Ⅳ种质与湖北糯玉米种质亲缘关系较近。
我国是公认的糯玉米起源中心, 据统计我国79.55%的糯玉米种质分布于西南地区(云南、广西、贵州、四川)[22]。本研究供试群体中包括来自北京、甘肃、河南、山东、江苏、上海、广西、广东和云南的糯玉米种质, 结合系谱信息和聚类结果可见, 大部分材料的亲缘关系与其地理来源相关; 而少部分材料推测是由于不同区域品种间进行了频繁的基因交流和重组, 遗传基础组成丰富, 失去了明显的区域特征, 导致聚类结果与地理来源信息不符。
综上结果说明目前我国糯玉米育种工作仍面临着亲本遗传基础狭窄的困境, 亟须加强对不同地区糯玉米种质资源的引进和创制, 为选育具有突破性的优质糯玉米新品种提供物质基础。
3.2 遗传相似系数聚类和Structure模型聚类结果比较
目前, 生产中应用的糯玉米自交系多来源于优质杂交种或中间群体, 很多材料已无系谱可查或遗传基础复杂, 无法利用系谱追踪法研究它们的亲缘关系。分子标记技术不受环境和季节限制、标记覆盖全基因组、多态性高、成本较低, 已成为遗传多样性分析最常用的技术。种质聚类方法有多种, 包括遗传相似系数法、遗传距离法、模型聚类法, 选择不同的参数会导致分析结果较大差异[23]。本研究通过GS将87份糯玉米自交系划分为4个类群, 分别包括9、66、10和2份材料; 而基于Structure模型的群体遗传结构分析也将这些材料分为4个居群, 分别包括24、25、19和19份材料。可见利用UPGMA方法和Structure模型聚类法划分的类群数一致, 说明本研究对群体类群的划分结果合理可靠。
基于遗传相似系数的聚类分析是根据材料间亲缘关系的远近, 类群划分时选择的GS水平受人为主观判定影响; 而群体遗传结构分析以亚群是否达到Hardy-Weinberg平衡建立的数学模型, 通过计算材料相应的Q值分析其组成成分来进行聚类划分。刘秀云等[24]通过UPGMA聚类分析将255个枣品种分为15个亚类; 同时, 利用群体结构分析也将这些材料划分为15个群体, 与聚类分析结果基本一致。王晋等[25]认为Structure软件的群体遗传结构分析可以避免人为因素对类群划分的影响, 进而对供试材料的群体结构进行校正。赵旭等[26]认为Structure的模型分析根据Δ峰值确定亚群间遗传分化不明显的复杂群体的类群数[13,27], 模型聚类方法可以将遗传距离聚类中系谱关系不明确的自交系划分到相应类群, Structure的模型聚类法更有利于研究玉米自交系的遗传关系, 将类群划分更细致; 而结合遗传相似系数和Structure模型聚类法能更好地确定自交系间的亲缘关系。本研究中虽然利用遗传相似系数聚类和Structure模型聚类划分的类群数相同, 但是各类群的种质组成差异较大, 推测这也是由于所利用的遗传相似系数聚类和Structure模型聚类计算方法不同导致的。因此, 进行种质聚类分析时, 根据种质已知信息, 选择合适的参数和算法对于准确评价群体间遗传多样性及种质的精细聚类分析具有重要意义。
4 结论
基于遗传相似系数, 通过UPGMA方法将87份糯玉米自交系划分为4个类群, 利用Structure软件进行群体遗传结构分析也将该群体聚类为4个居群, 但各类群包含的种质不同。本研究认为目前我们育种中使用的糯玉米自交系遗传变异较单一, 亟须引进和创制遗传基础丰富的种质资源。
[1] 丁照华, 孟昭东, 张发军, 汪黎明, 孙琪, 张庆伟. 我国糯玉米育种现状及发展对策. 玉米科学, 2006, 14(3): 46–48.Ding Z H, Meng Z D, Zhang F J, Wang L M, Sun Q, Zhang Q W. General situation and development priority of waxy corn breeding in China., 2006, 14(3): 46–48 (in Chinese with English abstract).
[2] 李玉军, 刘婷婷, 张泽志. 糯玉米起源、研究及发展概况. 耕作与栽培, 2010, (3): 52–53. Li Y J, Liu T T, Zhang Z Z. The origin, research and development of waxy maize., 2010, (3): 52–53 (in Chinese).
[3] 鲍坚东. 中国糯玉米起源与育种选择分子机制. 浙江大学博士学位论文, 浙江杭州, 2011. Bao J D. The Origin and Selective Breeding of Chinese Waxy Maize. PhD Dissertation of Zhejiang University, Hangzhou, Zhejiang, China, 2011 (in Chinese with English abstract).
[4] 刘丽君, 张丹, 薛林, 李建, 徐辰武. 基于SSR标记的江苏沿江地区糯玉米种质资源遗传多样性研究. 江苏农业学报, 2011, 27: 723–729.Liu L J, Zhang D, Xue L, Li J, Xu C W. Genetic diversity of waxy corn germplasm resources in Jiangsu coastal district by SSR markers., 2011, 27: 723–729 (in Chinese with English abstract).
[5] 吴斌, 李建, 柯洁, 熊志文, 胡美玲, 胡坤, 刘坚. 西双版纳糯玉米地方品种遗传多样性分析. 玉米科学, 2013, 21(2): 17–23.Wu B, Li J, Ke J, Xiong Z W, Hu M L, Hu K, Liu J. Genetic diversity of waxy maize landraces in Xishuangbanna., 2013, 21(2): 17–23 (in Chinese with English abstract).
[6] Zheng H J, Wang H, Yang H, Wu J H, Shi B, Cai R, Xu Y B, Wu A Z, Luo L J. Genetic diversity and molecular evolution of Chinese waxy maize germplasm, 2013, 8: e66606.
[7] Murray M G, Thompson W F. Rapid isolation of high molecular weight plant DNA., 1980, 8: 4321–4326.
[8] Wang F G, Tian H L, Zhao J R, Yi H M, Wang L, Song W. Development and characterization of a core set of SSR markers for fingerprinting analysis of Chinese maize varieties.2011, 56: 7–17.
[9] 王凤格. NY/T1432—2014玉米品种鉴定技术规程 SSR标记法. 北京: 中国农业出版社, 2014. pp 2–10. Wang F G. NY/T1432—2014 Technical Specification of Maize Variety Identification by SSR Markers. Beijing: China Agriculture Press, 2014. pp 2–10 (in Chinese).
[10] Lu Y, Xing L P, Xing S J, Hu P, Cui C F, Zhang M Y, Xiao J, Wang H Y, Zhang R Q, Wang X E, Chen P D, Cao A Z. Characterization of a putative new semi-dominant reduced height gene,, in wheat (L.)., 2015, 42: 685–698.
[11] Nei M. Genetic distance between populations, 1972, 106: 283–292.
[12] Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets., 2016, 33: 1870–1874.
[13] Evanno G, Regnaut S, Goudet J. Detecting the number of clusters of individuals using the software structure: a simulation study., 2005, 14: 2611–2620.
[14] 李凤艳, 张兴华, 张仁和. 玉米优异地方种质资源的筛选与评价. 植物遗传资源学报, 2003, 4: 225–227. Li F Y, Zhang X H, Zhang R H. The selection and evaluation of elite maize landraces., 2003, 4: 225–227 (in Chinese with English abstract).
[15] 石云素. 国家库玉米种质资源的保护与利用. 植物遗传资源学报, 2011, 12(3): 8. Shi Y S. Protection and utilization of national maize germplasm resources., 2011, 12(3): 8 (in Chinese).
[16] 赫忠友, 赫晋. 美玉(加甜糯) 8号鲜食专用玉米杂交种的选育. 作物杂志, 2005, (5): 68–69. He Z Y, He J. Breeding of fresh-eating hybrid maize—Meiyu 8., 2005, (5): 68–69 (in Chinese).
[17] 梁耿文. 糯玉米新组合田白糯1号的选育. 广东农业科学, 2006, (12): 17–18. Liang G W. Breeding of a new waxy corn variety Tianbainuo1., 2006, (12): 17–18 (in Chinese).
[18] 陈国清, 陆虎华, 薛林, 黄小兰, 石明亮, 印志同.早熟优质糯玉米杂交种苏玉糯2号的选育及应用. 金陵科技学院学报, 2006, 22(3): 71–74. Chen G Q, Lu H H, Xue L, Huang X L, Shi M L, Yin Z T. Breeding and application of early-mature, good-quality and high-yield waxy hybrid Suyunuo 2., 2006, 22(3): 71–74 (in Chinese with English abstract).
[19] 黄小兰, 孙权星, 陈小晖, 彭长俊, 陆冬梅. 苏玉糯1号的特征特性及其二环系的应用. 现代农业科技, 2012, (12): 47–49.Huang X L, Sun Q X, Chen X H, Peng C J, Lu D M. Characteristics of Suyunuo1 and application of Suyunuo1’s second cycle inbred lines., 2012, (12): 47–49 (in Chinese).
[20] 袁建华, 颜伟, 陈艳萍, 张跃中. 糯玉米杂交新组合——江南花糯. 江苏农业学报, 2002, 18: 240–240. Yuan J H, Yan W, Chen Y P, Zhang Y Z. New hybrid waxy maize combination—Jiangnanhuanuo., 2002, 18: 240–240 (in Chinese).
[21] 胡学安, 周波, 魏良明. 优质糯玉米新品种郑白糯6号选育研究. 农业科技通讯, 2007, (4): 17. Hu X A, Zhou B, Wei L M. Breeding of a new high quality waxy corn variety Zhengbainuo 6., 2007, (4): 17 (in Chinese).
[22] 刘永建. 用SSR标记研究西南糯玉米种质资源的遗传多样性. 四川农业大学博士学位论文, 四川成都, 2002. Liu Y J. Genetic Diversity of Waxy Corn Germplasms in Southwestern China Revealed by Simple Sequence Repeats. PhD Dissertation of Sichuan Agricultural University, Chengdu, Sichuan, China, 2002 (in Chinese with English abstract).
[23] 陈红菊, 岳永生, 樊新忠, 张传生, 杜立新. 山东地方鸡种遗传距离与聚类分析方法比较研究畜牧兽医学报, 2004, 35: 33–36.Chen H J, Yue Y S, Fan X Z, Zhang C S, Du L X. A comparative study of genetic distance and clustering analysis among Shandong indigenous chicken breeds.2004, 35: 33–36 (in Chinese with English abstract).
[24] 刘秀云, 李慧, 刘志国, 赵锦, 刘孟军. 基于SSR标记的255个枣品种亲缘关系和群体遗传结构分析. 中国农业科学, 2016, 49: 2772–2791.Liu X Y, Li H, Liu Z G, Zhao J, Liu M J. Genetic diversity and structure of 255 cultivars ofMill., 2016, 49: 2772–2791 (in Chinese with English abstract).
[25] 王晋, 王世红, 赖勇, 孟亚雄, 李葆春, 马小乐, 尚勋武, 王化俊. 大麦SSR标记遗传多样性及群体遗传结构分析核农学报, 2014, 28: 177–185.Wang J, Wang S H, Lai Y, Meng Y X, Li B C, Ma X L, Shang X W, Wang H J. Genetic diversity and population structure analysis by using SSR markers in barley., 2014, 28: 177–185 (in Chinese with English abstract).
[26] 赵旭, 方永丰, 王汉宁. 玉米SSR标记杂优类群划分及群体遗传结构分析. 核农学报, 2013, 27: 1828–1838.Zhao X, Fang Y F, Wang H N. Analysis of population genetic structure of maize by SSR marker and heterotic grouping., 2013, 27: 1828–1838 (in Chinese with English abstract).
[27] Lia V V, Poggio L, Confalonieri V A. Microsatellite variation in maize landraces from Northwestern Argentina: genetic diversity, population structure and racial affiliations, 2009, 119: 1053–1067.
附表1 91份糯玉米种质名称及系谱来源
Supplementary table 1 List of 91 waxy maize germplasm resources and their pedigrees

编号No.名称Name系谱Pedigree来源Origin N116SW-122W97/玉美头601♀ W97/Yumeitou 601♀上海 Shanghai N2W97不详 Unknown上海 Shanghai N3W148不详 Unknown上海Shanghai N416SW-292W75/W95上海Shanghai N516SW-296(江南花糯二环系/白478)/W22(Second cycle lines from Jiangnanhuanuo/Bai 478) /W22江苏 Jiangsu N6通系5 Tongxi 5不详 Unknown江苏 Jiangsu N716SW-311申W22/H54 Shen W22/H54上海 Shanghai N816SW-312京W♂/W100 JingW♂/W100北京 Beijing N916SW-320W82/(W88/紫黑W1号) W82/(W88/Zihei W1)上海 Shanghai N1016SW-321申W22/(南汇黑/申W13) Shen W22/(Nanhuihei/Shen W13)上海 Shanghai N1116SW-322W96/张红 W96 / Zhanghong甘肃 Gansu N1216SW-324W75/[江南花糯二环系/(祝西/5003)]W75/[Second cycle lines from Jiangnanhuanuo/(Zhuxi/5003)]江苏 Jiangsu N1316SW-327W75/玉美头601♀ W75/Yumeitou 601♀广西 Guangxi N14京W♂ Jing W♂不详 Unknown北京 Beijing N1516SW-333申W48/张红 Shen W48/Zhanghong甘肃 Gansu N16W153不详 Unknown上海 Shanghai N17W152不详 Unknown上海 Shanghai N18W158不详 Unknown上海 Shanghai N1916SW-347W93/京W♂ W93/Jing W♂北京 Beijing N2016SW-349申W22/H88 Shen W22/H88上海 Shanghai N2116SW-350京W♂/H54 JingW♂/H54北京 Beijing N2216SW-363(江南花糯二环系/衡白522)/(江南花糯二环系/G478)(Second cycle lines from Jiangnanhuanuo/Hengbai 522)/(Second cycle lines from Jiangnanhuanuo/G478)江苏 Jiangsu N2316SW-365美玉8号二环系 Second cycle lines from Meiyu8广西 Guangxi N2416SW-367(通系5/昌7-2)/W88 (Tongxi 5/Chang 7-2)/W88不详 Unknown N2516SW-369[江南花糯二环系/(祝西/5003)]/(江南花糯二环系/衡白522)[Second cycle lines from Jiangnanhuanuo/(Zhuxi/5003)]/(Second cycle lines from Jiangnanhuanuo/Hengbai 522)江苏 Jiangsu N2616SW-37307东N-1/玉美头601♀ 07DongN-1/Yumeitou 601♀广西 Guangxi N2716SW-374W95/玉美头601♀ W95/Yumeitou 601♀广西 Guangxi N2816SW-424W93/西星黑糯1号二环系W93/Second cycle lines from Xixingheinuo 1山东 Shandong N29W116不详 Unknown上海 Shanghai N3016SW-426W88/申W22 W88/Shen W22上海 Shanghai N3116SW-427W75/[(W01/申W48)/W01] W75/[(W01/Shen W48)/W01]上海 Shanghai N3216SW-429W01/中糯1号OP W01/Zhongnuo 1 OP北京Beijing N3316SW-437苏玉糯1号二环系 Second cycle lines from Suyunuo1江苏 Jiangsu N3416SW-438[通系5/(515/小黄糯)]/江南花糯二环系[Tongxi 5/(515/Xiaohuangnuo)]/ Second cycle lines from Jiangnanhuanuo云南 Yunnan N3516SW-439[通系5/(515/小黄糯)]/(申W13/通系5)[Tongxi 5/(515/ Xiaohuangnuo)]/(Shen W13/Tongxi 5)云南 Yunnan
(附续表1)
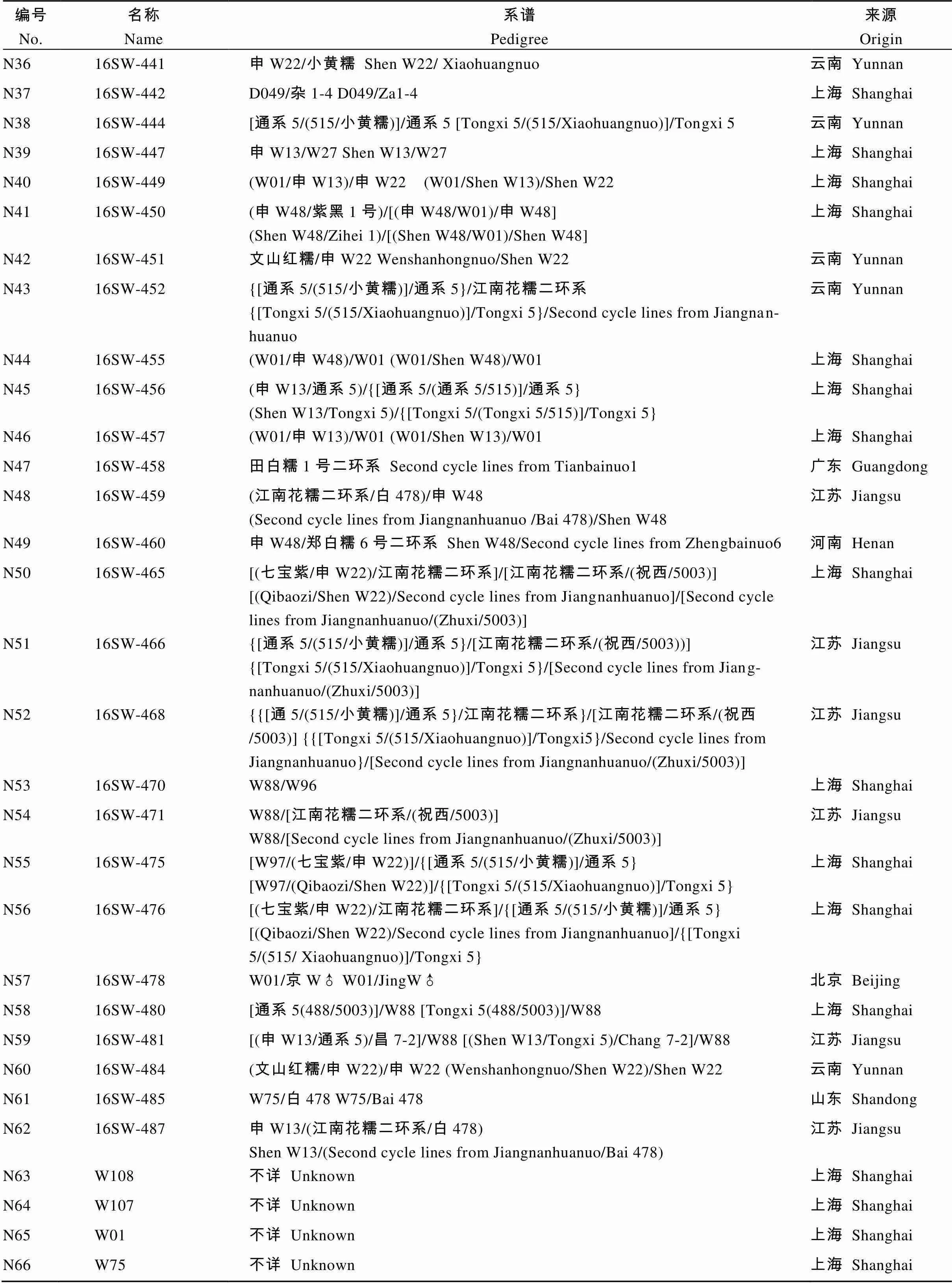
编号No.名称Name系谱Pedigree来源Origin N3616SW-441申W22/小黄糯 Shen W22/ Xiaohuangnuo云南 Yunnan N3716SW-442D049/杂1-4 D049/Za1-4上海 Shanghai N3816SW-444[通系5/(515/小黄糯)]/通系5 [Tongxi 5/(515/Xiaohuangnuo)]/Tongxi 5云南 Yunnan N3916SW-447申W13/W27 Shen W13/W27上海 Shanghai N4016SW-449(W01/申W13)/申W22 (W01/Shen W13)/Shen W22上海 Shanghai N4116SW-450(申W48/紫黑1号)/[(申W48/W01)/申W48](Shen W48/Zihei 1)/[(Shen W48/W01)/Shen W48]上海 Shanghai N4216SW-451文山红糯/申W22 Wenshanhongnuo/Shen W22云南 Yunnan N4316SW-452{[通系5/(515/小黄糯)]/通系5}/江南花糯二环系{[Tongxi 5/(515/Xiaohuangnuo)]/Tongxi 5}/Second cycle lines from Jiangnanhuanuo云南 Yunnan N4416SW-455(W01/申W48)/W01 (W01/Shen W48)/W01上海 Shanghai N4516SW-456(申W13/通系5)/{[通系5/(通系5/515)]/通系5} (Shen W13/Tongxi 5)/{[Tongxi 5/(Tongxi 5/515)]/Tongxi 5}上海 Shanghai N4616SW-457(W01/申W13)/W01 (W01/Shen W13)/W01上海 Shanghai N4716SW-458田白糯1号二环系 Second cycle lines from Tianbainuo1广东 Guangdong N4816SW-459(江南花糯二环系/白478)/申W48(Second cycle lines from Jiangnanhuanuo /Bai 478)/Shen W48江苏 Jiangsu N4916SW-460申W48/郑白糯6号二环系 Shen W48/Second cycle lines from Zhengbainuo6河南 Henan N5016SW-465[(七宝紫/申W22)/江南花糯二环系]/[江南花糯二环系/(祝西/5003)][(Qibaozi/Shen W22)/Second cycle lines from Jiangnanhuanuo]/[Second cycle lines from Jiangnanhuanuo/(Zhuxi/5003)]上海 Shanghai N5116SW-466{[通系5/(515/小黄糯)]/通系5}/[江南花糯二环系/(祝西/5003))]{[Tongxi 5/(515/Xiaohuangnuo)]/Tongxi 5}/[Second cycle lines from Jiangnanhuanuo/(Zhuxi/5003)]江苏 Jiangsu N5216SW-468{{[通5/(515/小黄糯)]/通系5}/江南花糯二环系}/[江南花糯二环系/(祝西/5003)] {{[Tongxi 5/(515/Xiaohuangnuo)]/Tongxi5}/Second cycle lines from Jiangnanhuanuo}/[Second cycle lines from Jiangnanhuanuo/(Zhuxi/5003)]江苏 Jiangsu N5316SW-470W88/W96上海 Shanghai N5416SW-471W88/[江南花糯二环系/(祝西/5003)]W88/[Second cycle lines from Jiangnanhuanuo/(Zhuxi/5003)]江苏 Jiangsu N5516SW-475[W97/(七宝紫/申W22)]/{[通系5/(515/小黄糯)]/通系5}[W97/(Qibaozi/Shen W22)]/{[Tongxi 5/(515/Xiaohuangnuo)]/Tongxi 5}上海 Shanghai N5616SW-476[(七宝紫/申W22)/江南花糯二环系]/{[通系5/(515/小黄糯)]/通系5}[(Qibaozi/Shen W22)/Second cycle lines from Jiangnanhuanuo]/{[Tongxi 5/(515/ Xiaohuangnuo)]/Tongxi 5}上海 Shanghai N5716SW-478W01/京W♂ W01/JingW♂北京 Beijing N5816SW-480[通系5(488/5003)]/W88 [Tongxi 5(488/5003)]/W88上海 Shanghai N5916SW-481[(申W13/通系5)/昌7-2]/W88 [(Shen W13/Tongxi 5)/Chang 7-2]/W88江苏 Jiangsu N6016SW-484(文山红糯/申W22)/申W22 (Wenshanhongnuo/Shen W22)/Shen W22云南 Yunnan N6116SW-485W75/白478 W75/Bai 478山东 Shandong N6216SW-487申W13/(江南花糯二环系/白478)Shen W13/(Second cycle lines from Jiangnanhuanuo/Bai 478)江苏 Jiangsu N63W108不详 Unknown上海 Shanghai N64W107不详 Unknown上海 Shanghai N65W01不详 Unknown上海 Shanghai N66W75不详 Unknown上海 Shanghai
(附续表1)

编号No.名称Name系谱Pedigree来源Origin N67W151不详 Unknown上海 Shanghai N68W114不详 Unknown上海 Shanghai N69张改♀ Zhanggai ♀不详 Unknown上海 Shanghai N70wh-2不详 Unknown上海 Shanghai N7116HW-31(江南花糯二环系/白478)/W95(Second cycle lines from Jiangnanhuanuo/Bai 478)/W95山东 Shandong N7216HW-46(W96/张红)/申W22 (W96/Zhanghong)/Shen W22甘肃 Gansu N7316AW-1001引种大/W95 Yinzhongda/W95上海 Shanghai N7416AW-1003申W48/[(申W48/紫黑1号)/申W48] Shen W48/[(Shen W48/Zihei 1)/Shen W48]上海 Shanghai N7516AW-1004[(申W48/W01)/申W48]/[(申W48/紫黑1号)/申W48] [(Shen W48/W01)/Shen W48]/[(Shen W48/Zihei 1)/Shen W48]上海 Shanghai N7616AW-1005[(江南花糯二环系/白478)/郑白糯6号二环系]/京W♂[(Second cycle lines from Jiangnanhuanuo/Bai 478)/Second cycle lines from Zhengbainuo6]/Jing W♂河南 Henan N7716AW-1006[(W48/5003)/W48]/中糯2号二环系[(W48/5003)/W48]/Second cycle lines from Zhongnuo 2北京 Beijing N78查12at-1385Cha 12at-1385不详 Unknown上海 Shanghai N7916AW-1008张红/[江南花糯二环系/(祝西/5003)]Zhanghong/[Second cycle lines from Jiangnanhuanuo/(Zhuxi/5003)]江苏 Jiangsu N8009N-5M不详 Unknown上海 Shanghai N8109N-6M不详 Unknown上海 Shanghai N8209N-S150P不详 Unknown上海 Shanghai N8316AW-1015[(申W48/紫黑1号)/(申W48/W01)]/申W22 [(Shen W48/Zihei 1)/(Shen W48/W01)]/Shen W22上海 Shanghai N8416AW-1019申W48/郑白糯6号二环系 Shen W48/Second cycle lines from Zhengbainuo 6河南 Henan N85W-109不详 Unknown上海 Shanghai N86申W93 Shen W93荆黑糯2号二环系 Second cycle lines from Jingheinuo 2上海 Shanghai N87黄W-♀ Huang W- ♀不详 Unknown上海 Shanghai N8816HW-LY516HW-31/申W48 16HW-31/Shen W48上海 Shanghai N8916HW-LY816HW-4/16HW-46上海 Shanghai N90沪紫黑糯2号Huziheinuo 2申W74/申W93 Shen W74/Shen W93上海 Shanghai N91五彩甜糯2号Wucaitiannuo 2申W93/wh-2 Shen W93/wh-2上海 Shanghai
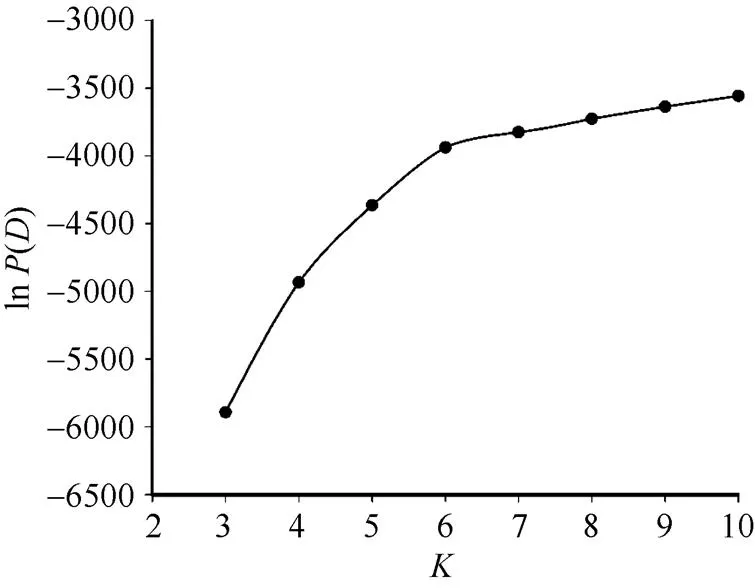
附图1 ln P(D)随K值的变化
Geneticdiversity and population structure analysis by SSR markers in waxy maize
LU Yuan1,2, AI Wei-Da3, HAN Qing1,2, WANG Yi-Fa1, LI Hong-Yang4, QU Yu-Ji5, SHI Biao1,2,*, and SHEN Xue-Fang1,2,*
1Crop Breeding and Cultivation Research Institute, Shanghai Academy of Agricultural Sciences, Shanghai 201403, China;2CIMMYT: China Specialty Maize Research Center, Shanghai 201403, China;3College of Agriculture, Anshun University, Anshun 561000, Guizhou, China;4Sanya Sci-Tech Academy of Hainan National Breeding and Multiplication, Sanya 572000, Hainan, China;5Agricultural Service Center of Qingpu Town in Zhujiajiao District of Shanghai, Shanghai 201713, China
To understand the genetic basis of waxy maize (L.var.Kulesh), 87 waxy maize inbred lines were screened by 29 pairs of SSR markers for genetic diversity analysis. Totally, 180 alleles were detected in this population, with an average of 6 alleles per locus. Polymorphic information content (PIC) ranged from 0.308 to 0.915, with an average of 0.572. The genetic similarity between materials ranged from 0.49 to 0.93 with an average of 0.66. Based on the UPGMA (unweighted pair-group method with arithmetic means) cluster analysis, divided 87 waxy maize inbred lines into four groups with genetic similarity of 0.64, including 9, 66, 10, and 2 materials. In addition, 87 waxy maize inbred lines were also divided into four genetic subpopulations, including 24, 25, 19, and 19 materials, by the population genetic structure analysis. Further study showed that genetic variation among most of waxy maize inbred lines in this population was relatively simple. This study provides a germplasm base and theoretical basis for new variety breeding and evolutionary genetics analysis of waxy maize.
waxy maize; genetic diversity; population structure
2018-01-15;
2018-10-08;
2018-11-03.
10.3724/SP.J.1006.2019.83008
施标, E-mail: 18918162109@163.com; 沈雪芳, E-mail: shenxuefang68@163.com
本研究由上海市种业发展项目[沪农科种字(2016)第1-3号], 上海市科学技术委员会科研计划项目(153919N1300, 17391900100, 17391902702), 上海市农业科学院卓越团队项目(农科创2017B-08)资助, 上海市市级农口系统青年人才成长计划项目[沪农青字(2015)第1-4号], 上海市市级农口系统青年人才成长计划项目[沪农青字(2018)第1-6号]和2019上海市农业科学院青年科技人员“助跑”计划项目资助。
This study was supported by the Shanghai Agriculture Applied Technology Development Program, China (20160103), the Research Project of Shanghai Science and Technology Commission, China (153919N1300, 17391900100, 17391902702), and the Shanghai Academy of Agricultural Sciences Excellence Team Project, China (2017B-08), the Youth Talent Development Plan of Shanghai Municipal Agricultural System, China (20150104), the Youth Talent Development Plan of Shanghai Municipal Agricultural System, China (20180106), and the Youth Science and Technology Personnel of Shanghai Academy of Agricultural Sciences.
URL:http://kns.cnki.net/kcms/detail/11.1809.S.20181101.1001.002.html
猜你喜欢
中国种业(2022年8期)2022-08-19
农业技术与装备(2022年4期)2022-06-29
种子(2021年8期)2021-09-23
农业科技与信息(2020年14期)2020-12-18
农村实用技术(2020年4期)2020-12-15
北京农学院学报(2020年1期)2020-03-09
农民致富之友(2020年2期)2020-02-12
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
陕西农业科学(2019年6期)2019-07-19
