
商陆属2种植物花器官发生过程观察
2018-06-23 06:59:00郑宏春拓向辉马春兰王菲菲张一开郭巧慧
西北植物学报 2018年5期
郑宏春,拓向辉,马春兰,王菲菲,张一开,郭巧慧
(陕西学前师范学院 生命科学与食品工程学院,西安 710100)
商陆科由于商陆属植物所具有的独特特征,使商陆科的地位备受关注。商陆属花被1轮,5枚,雄蕊6~33枚,1轮或多轮,心皮5~16枚,排成1轮,基部合生上部分离,或除花柱柱头外全合生[1-3]。在近些年的研究中关于商陆科的地位出现了不同的观点,这些观点主要是因为分子生物学的研究结果和形态学观察结果不同所产生的。
许多研究者[4-10]的研究结果都不再支持原来认为该科为石竹亚纲(纲)的基部类群[11-14]的观点,这些研究者都是依据分子进化的研究结果所得出。Ronse De Craene等[15]对商陆属的花器官发生做了比较深入的研究,尤其通过对其雄蕊发生式样的讨论,认为商陆属植物花是从3基数花演化来的。陈丹等[16]通过对商陆属的PhytolaccadodecandraLHérit花器官发生的观察,支持Ronse De Craene 等[15]的观点,认为商陆属植物的花是起源于3基数花。郑宏春等[17-18]通过对商陆属几种具有不同花构成植物的花器官发生和发育的观察,认为商陆属植物花器官发生中,雄蕊的发生是以环状分生组织为基础,并不表现为3基数的发生,并根据这一结果,认为商陆科依然是石竹亚纲的基部类群。
Ronse De Craene等[15]总结了商陆属花和雄蕊之间关系的3个主要观点:1) 一个古老的观点,直到后来的一些作者仍支持的是,商陆属的雄蕊基本是外轮对萼雄蕊(具有10枚雄蕊);2) 早期的器官发生报告显示雄蕊多数是离心的轮式发育;3) Ronse De Creane等[15]建议石竹目的5基数花可能是从3基数花的2轮花被简化来的,随后在2个花萼的融合点的雄蕊丢失[19]。郑宏春等[17-18]在研究了3种商陆属植物的花器官发生后,认为商陆属植物的雄蕊的发生过程不能说明其雄蕊的发生具有3基数的特点或迹象,认为商陆属的雄蕊是在环状分生组织上发生的,它们的发生没有很清楚的对应规律。因此,商陆属花发生的模式对于确定商陆科的系统地位,有着重要的意义,它是商陆科能不能作为石竹亚纲的基部类群的重要依据,也是研究石竹亚纲植物起源的重要问题。
商陆属植物的花器官发生虽已有6种植物被详细地进行了研究,但是得出了2种完全不同的结论,也没有对商陆属的花发生模式进行过探讨。本研究对产于中国的红蕊商陆(Phytolaccaesculentavon Houtte)和浙江商陆(PhytolaccazhejiangensisW. T. Fan)进行了扫描电镜观察,并结合已经有的观察对商陆属植物花器官发生的模式进行讨论,以确定商陆属植物花发生的模式和商陆属植物花的基数,以及雄蕊和雌蕊是否具有叶性器官发生的特点。
1 材料和方法
红蕊商陆(Ph.esculenta)采自广西药用植物园,浙江商陆(Ph.zhejiangensis)采自浙江天目山自然保护区,凭证标本保存于中国科学院植物研究所植物标本馆(PE)。植物的花序经FAA固定,解剖镜下将花发育的不同阶段外部成熟的结构剥去,露出正在发生的器官原基和器官发育的早期阶段,经过下列程序给材料脱水,并通过醋酸异戊酯替换材料中的酒精。70%酒精→85%酒精→95%酒精→纯酒精→3/4纯酒精+1/4醋酸异戊酯→1/2纯酒精+1/2醋酸异戊酯→3/4纯酒精+1/4醋酸异戊酯→纯醋酸异戊酯,每步骤处理时间均为15 min。然后经HCP-2 CO2临界点干燥仪干燥。用导电双面胶将材料粘在铝合金台子上,SPI-MODULE离子溅射仪镀金膜。Hitachi-800扫描电镜观察照相。
2 观察结果
2.1 花被的发生
扫描电镜照片显示,红蕊商陆和浙江商陆植物的花发育均在花原基发生后(图版Ⅰ,1~4;图版版Ⅲ,1~3),小苞片以2/5圆周相继发生,小苞片B1和B2按顺时针方向相隔2/5圆周发生(图版Ⅰ,3),但2个小苞片不是对生(图版Ⅰ,3、4; 图版Ⅲ,2、3)。第1枚花被的发生位置决定于2枚苞片的发生顺序,2种植物中小苞片发生的位置,影响第1枚花被发生的位置,如果2个小苞片的发生沿近轴侧开始,同时小苞片按顺时针方向发生(图版Ⅰ,5、6;图版Ⅲ,3~5),则花被也是顺时针发生;若小苞片按逆时针方向发生,则花被发生也是逆时针的(图版Ⅰ,5、6;图版Ⅲ,6)。
花被片的发生是紧接小苞片的发生进行的(图版Ⅰ,5;图版Ⅲ,3),第1枚花被发生在距第2枚小苞片2/5圆周的位置,和苞片发生的方向相同。紧接着第2枚花被同样在距第1枚花被片2/5圆周的位置发生,同样也是在距离第2枚小苞片2/5圆周的位置。紧接着是第3枚花被在距第2枚2/5圆周的位置发生,接着是第4枚、第5枚以同样的距离和方向发生。可见花被的发生有顺时针(图版Ⅰ,8;图版Ⅲ,4~6)和逆时针(图版Ⅰ,6、7、9;图版Ⅲ,6)方向。
2.2 雄蕊的发生
红蕊商陆和浙江商陆植物花被发生结束时,顶端分生组织呈五边形,2种植物雄蕊在开始发生前,顶端会发生一个短暂的生长过程(图版Ⅱ,1~3;图版Ⅲ,9;图版Ⅳ,1),顶端分生组织长成一个近似圆形的形态(图版Ⅱ,1;图版Ⅲ,7、8)。雄蕊发生时,雄蕊原基没有明显的先后顺序,几乎是同时发生的(图版Ⅱ,1、2;图版Ⅲ,9;图版Ⅳ,1),所有雄蕊原基形成时,没有太大的大小差别(图版Ⅱ,1、2;图版Ⅳ,1),稍大一点也不与紧接其后的雄蕊原基有位置上的规律(图版Ⅱ,3;图版Ⅳ,1)。有时2个相当大小的雄蕊原基会紧挨在一起(图版Ⅱ,3),偶尔也会看到雄蕊和花被位置似乎有确定关系(图版Ⅱ,4~6),其外轮雄蕊有少数似与花被互生,但由于外轮雄蕊数多于花被数,不会每个雄蕊都与花被互生,也没有规律性。
雄蕊的数目在不同的花中有变化(图版Ⅳ),也会造成这种位置关系的不确定,如在红蕊商陆中就观察到7枚雄蕊(图Ⅳ,9)。雄蕊几乎是同时发生于花原基的顶端周围,这个区域被称为环状分生组织(图版Ⅲ,8中的RM所示)。雄蕊发生之初大小在不同的花中没有与花被对应的规律。从红蕊商陆和浙江商陆观察发现,内轮雄蕊先发生,外轮雄蕊发生较晚(图版Ⅱ,4、5)。
2.3 雌蕊的发生
从图版Ⅱ,5和图版Ⅳ,2可以看出,红蕊商陆和浙江商陆的心皮都是在雄蕊发生后,紧接着雌蕊开始发生,形成与雄蕊互生的突起,这些突起起初是分离的(图版Ⅱ,5~7;图版Ⅳ,2~4),随后其基部组织参加雌蕊的形成,而将雌蕊的每一个心皮在基部连成一个整体(图版Ⅱ,8、9;图版Ⅳ,5~9)。雌蕊基部发育成为整个雌蕊的子房,上部分离部分生长,最后成为花柱和柱头部分。观察发现,子房在2个种早期都是联合的。在浙江商陆,这个联合的部分生长较多,始终表现为合生心皮(图版Ⅱ,8、9),因此形态学上看作合生心皮。在红蕊商陆(Ph.esculenta)则不同,子房上部离生部分在花柱生长时,同时有较多的生长,而基部合生的部分生长较少,从而使子房的大部分表现为分离(图版Ⅳ,8、9),到成熟时,分离的部分占很大的比例,因此形态学上认为是离生心皮。
3 讨 论
3.1 花被的发生
最近对植物花基数发生的研究认为,5基数花在花被发生时花被都是以2/5圆周方式发生的,6基数(3基数)的是以1/3圆周方式发生的[20-21]。由于商陆属植物的花被为5,偶尔可见6枚。而且它的发生方式是2/5圆周,所以不能认为它是3基数而不是5基数。本研究观察发现,红蕊商陆和浙江商陆在花器官的发生中,在花的3个苞片中最下面一个苞片是最先从花原基基部发生的,然后在两侧先后发生2个小苞片,接着就顺序发生5个花被。在小苞片和花被之间没有什么明显的时间界限和形态的区别,只是在进一步的发育中,最先发生的2枚小苞片,逐渐地到了花柄的中下部,形态也不像花被,而成为小苞片,这显然是由于花柄上部的居间生长造成。因此可以认为商陆属植物的花被形成是对叶子在茎上的形成方式的一种继承或继续。这可以更好地解释商陆属植物中偶尔出现的6枚花被,和在商陆科中演化出具有4枚花被的植物。
3.2 商陆属植物雄蕊的发生
商陆属植物雄蕊的形成是提出商陆属植物起源于3基数花的重要证据来源[15-16]。我们认为商陆属植物的雄蕊发生与花被发生之间存在一个明显的时间间隔,在这一段时间里顶端的分生组织由原来五边形生长成近圆形。这个过程虽然短暂,这说明雄蕊发生可能需要进行与花被发生不同的准备,这种准备是植物形成雄蕊所必需的。因此可以说,商陆属植物雄蕊是与叶不同源的器官,至少它的形成所需要的内部条件与花被形成的内部条件可能有较大区别。植物在开始雄蕊发生前进行这种准备需要一定的时间,在形态上会发生一定的变化。商陆属植物雄蕊的发生次序实际上是没有固定规律的,无论用多小的时间间隔,找不出雄蕊发生顺序的连续照片。前面提到的两位作者所给出的图片上显示,在不同的位置的雄蕊有大小差别,当然在一般情况下这种差别可以作为雄蕊发生时间顺序的依据,但是在对多个花观察之后,我们注意到,雄蕊的大小并没有一致规律,因此我们不认为商陆属植物雄蕊的发生具有3基数花的特点。商陆属植物雄蕊的发生是在顶端的环状分生组织上形成的[17-18],环状分生组织上形成多少雄蕊是和环状分生组织的大小及每个雄蕊原基大小有关,而不和一定的基数有关。这样我们可以容易地说明,在商陆属中的雄蕊和心皮的数目的变化,也能简单地说明为什么商陆属植物雄蕊的变化总是不遵守通常按轮排列植物所遵守的规律。同时也说明所谓外轮雄蕊对萼的情况是不存在的,从发生上看少于10枚的雄蕊,都在1轮上发生。多于10枚雄蕊的都会有2轮。有许多研究者支持商陆属植物为外轮雄蕊对萼[22-27],本研究的结果不支持这样的观点。从花器官发生来看,当雄蕊少于10枚时,雄蕊只有1轮,并没有形成2轮,少于10枚雄蕊时不存在外轮雄蕊对萼的问题。当雄蕊多于10枚时雄蕊排成2轮,外轮雄蕊的发生晚于内轮雄蕊,与内轮雄蕊也没有明确的位置关系,加之雄蕊数目变化多,外轮雄蕊也不是确定地对萼。本研究的结果支持雄蕊多数时,雄蕊呈离心式发育的观点[28-29],从发生和形态看,雄蕊都不具有叶性器官的特点。
3.3 心皮的发生
心皮的发生是一个较少争论的问题,因为心皮的发生基本上是同时等大的,所以不会显示出一定顺序来,它也不与花被成规律对应排列。但是心皮与内轮雄蕊相间排列成互生状是一个明显的特征。心皮的发生和雄蕊的发生之间是完全连续的,没有任何可以观察到的间隔,这一点说明,雄蕊和心皮发生所需要的内部条件可能是一致的,至少是可以迅速过渡的,它们在起源的性质上应该是一致的。从心皮的发生上也可以看出商陆属是比较原始的,我们仍然支持商陆科是石竹目中的基部类群,不支持它是石竹目中比较进化的类群的观点。
3.4 斐波那契(Fibonacci)数列与花器官发生
斐波那契数是近些年来经常提到的关于植物花器官发生的一个重要的概念[20, 30]。在商陆属的花器官发生中同样存在斐氏数,首先2/5圆周的发生本身就有2个斐氏数,2、5都是斐氏数,2/5圆周所转过的角度也正好是一个斐氏数144°。虽然目前还没有关于斐氏数在生物中大量出现原因的研究报道,但是这可能是生物起源与进化中所遵守的一个非常重要的规律。因此商陆属花的基数5,以及心皮出现的8可能是该属起源的2个原始的基数,因为8也是一个斐氏数,而具有10心皮的种可能是由具有5心皮的植物经过器官的加倍所产生的。而8是一个自然基数,但是由于8这个基数本身所存在的问题并没有得到太多的发展,它要求较大的空间,因此这个基数的植物仅有商陆1个属。研究斐氏数在植物的形态构成和演化中的原理可能是揭开植物系统演化的一个重要途径。
3.5 商陆属植物花器官的发生模式
花形态发生和发育提供了重要的关于器官进化,形态发生和系统关系的信息,进一步使我们理解被子植物的系统发生[31]。商陆属植物的花由于它的特殊性,长期以来没有能够合理地说明它的起源历史。它所存在的问题是,花被是5枚,而雄蕊和雌蕊的变化使得用于其他类群的花器官起源的解释都显得有些不足。也就是说我们看不出它是否就是遵守5基数花的起源规律,也看不出它是否遵守其它基数花的起源规律。Ronse De Craene等[15]认为商陆花为3基数花的根据是不充分的。本观察结果也说明商陆属植物的花被是5基数的,并没有3基数的任何痕迹。加之雄蕊是发生于环状分生组织上,所以没有表现出一定基数的发生特点。
综上所述,商陆属植物的花器官的发生可以分成两个部分:花被的发生是苞片发生的继续,为2/5圆周,是5基数,是一个斐波那契(Fibonacci)数;雄蕊和雌蕊的发生是产生于环状分生组织上的,几乎是同时发生,没有发生顺序,不遵守基数特征,而发育空间对花部数目有较大的影响。雄蕊不具有明显的叶性器官发生的特点,2轮时离心的轮式发生,无对萼外轮雄蕊。因此,商陆科仍是石竹目的原始类群。
参考文献:
[1] WALTER H. Phytolaccaceae[M]. Wilhelm Engelmann: Leipzig, 1909.
[2] HEIMERL A. Phytolaccaceae[M]// in Die natürlich Pflanzenfamilien, A. Engler and K. Prantl, Editor. Wilhelm Engelmann: Leipzig. 1934: 1-14.
[3] ROHWER J G. Phytolaccaceae[M]// in The Families and Genera of Vascular Plants II, K. Kubitzki, J.G. Rohwer and V. Bitrich, Editors. Springer-Verlag: Berlin. 1993: 506-515.
[4] MANHART J R, RETTIG J H. Gene sequence data[M]// in Caryophyllales: Evolution and Systematics, H-D Behnke and T J Mabry, Editors. Springer-Verlag: Berlin, Heidelberg. 1994: 236-245.
[5] RETTIG J H, WILSON H D, MANHART J R. Phylogeny of Caryophyllaces-gene sequence data[J].Taxon, 1992,41: 201-209.
[6] DOWNIE S R, PALMER J D. A chloroplast DNA phylogeny of Caryophyllales based on structure and inverted repeat restriction site variation[J].SystematicBotany, 1994,19: 236-252.
[7] APG. An ordinal classification for the families of flowering plants[J].AnnalsoftheMissouriBotanicalGarden, 1998,85: 531-553.
[8] APG. An update of the angiosperm phylogeny group classification for orders and families of flowering plants: APG II[J].BotanicalJournaloftheLinneanSociety, 2003,141: 399-436.
[9] APG. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III[J].BotanicalJournaloftheLinneanSociety, 2009,161: 105-121.
[10] APG. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV[J].BotanicalJournaloftheLinneanSociety, 2016,181(1): 1-20.
[11] TAKHTAJAN A. Flowering Plants[M]. Berlin: Springer. 2009: 90-104.
[12] DAHLGREN R. A revised system of classification of the angiosperms[J].BotanicalJournaloftheLinneanSociety, 1980.80: 91-124.
[13] CRONQUIST A. An Integrated system of Classification of Flowering Plants[M]. New York: Columbia University, 1981.
[14] WU Z Y, LU A M, TANG Y C. Synopsis of a new “polyphyletic-polychronic-polytopic” system of the angiosperm[J].ActaPhytotaxonomicaSinica, 2002,40: 289-322.
[15] RONSE DE CRAENE L, VANVICKENROYE P, SMETS E F. A study of floral morphological diversity in Phytolacca (Phytolacaceae) based on early floral ontogeny[J].InternationalJournalofPlantScience, 1997,158(1): 57-72.
[16] CHEN D, WANG Q. Pentamerous flowers in the genus Phytolacca have been derived from trimerous flowers-New evidence from the floral organogenesis of Phytolacca dodecandra[J].AcatPhytotaxonomicaSinica, 2004,42(4): 345-351.
[17] ZHENG H C, LU A M, HU Z H. Floral Organogenesis in Phytolacca (Phytolaccaceae)[J].AcatPhytotaxonomicaSinica, 2004,42: 352-364.
[18] ZHENG H C, LU A M, HU Z H. Floral Organogenesis and Ring Meristem in Phytolacca[J].AmericanJournalofPlantSciences, 2015,6: 445-455.
[19] RONSE DE CRAENE, L. SMETS E.F. Merosity in flowers: Definition, origin, and taxonomic significance[J].PlantSystematicsandEvolution, 1994.191(1): p. 83-104.
[20] RONSE DE CRAENE L. Meristic changes in flowering plants: How flowers play with numbers[J].Flora, 2016, 221: 22-37.
[21] DOS SANTOS P AND L P RONSE DE CRAENE. Floral development of Lewisia (Montiaceae): Investigating patterns of perianth and stamen diversity[J].Flora, 2016, 221: 4-13.
[22] EICHLER A W.BlüthendiagrammeII[M]. Leipzig: Wilhelm Engelmann,1878.
[23] HEIMERL A. Phytolacaceae[M]// in Die Natürlich Pflanzenfamilien. A. Engler and K. Prantl, Editors. Wilhelm Engelmann: Leipzig. 1889.
[24] WALTER H. Die diagramme derPhytolaccaceen[J].Bot.Jahrb.Syst., 1906,37(suppl 85): 1-57.
[25] FRIEDRICH H-C. Studenüben die natürliche verwandtschaft der plambaginales und centrspermae[J].Phyton(Austria), 1956,6: 220-263.
[26] LEINS P AND C ERBAR. Putative Origin and Relationships of the Order from the Viewpoint of Developmental Flower Morphology[M]// in Caryophyllales:Evolutio and Systematics, H.-D. Behnke and T.J. Mabry, Editors. Springer-verlag: Berlin. 1994: 303-316.
[27] LEINS P, ERBAR C. Flowers in Magnoliidae and the origin of flowers in other subclasses of the angiosperms. II. The relationships between flowers of Magnoliidae, Dilleniidae, and Caryophyllidae, in Early Evolution of Flowers. Plant Systematics and Evolution Supplement 8[M], P K Endress and E M Friis, Editors. Springer Vienna: Vienna. 1994: 209-218.
[28] ECKDART T. Classical morphological features of Centrospermous families[J].PlantSystematicsandEvolution, 1976,126: 5-25.
[29] BUXBAUM F. Vorlaufige untersuchungen uber umfang systematische stellung und Gliderung der Caryophyllales (Centrospermae)[J].BeitrageBiologiePflanzen, 1961,36: 1-56.
[30] NISHIYAMA Y. Why is a flower five-petaled[J].JournalofScienceEducationandTechnology, 2004,13(1): 352-364.
[31] LIU X L, LI J H, YANG Y F, ZHU J Y. Floral development ofGymnospermiummicrorrhynchum(Berberidaceae) and its systematic significance in the Nandinoideae[J].Flora, 2017, 228: 10-16.
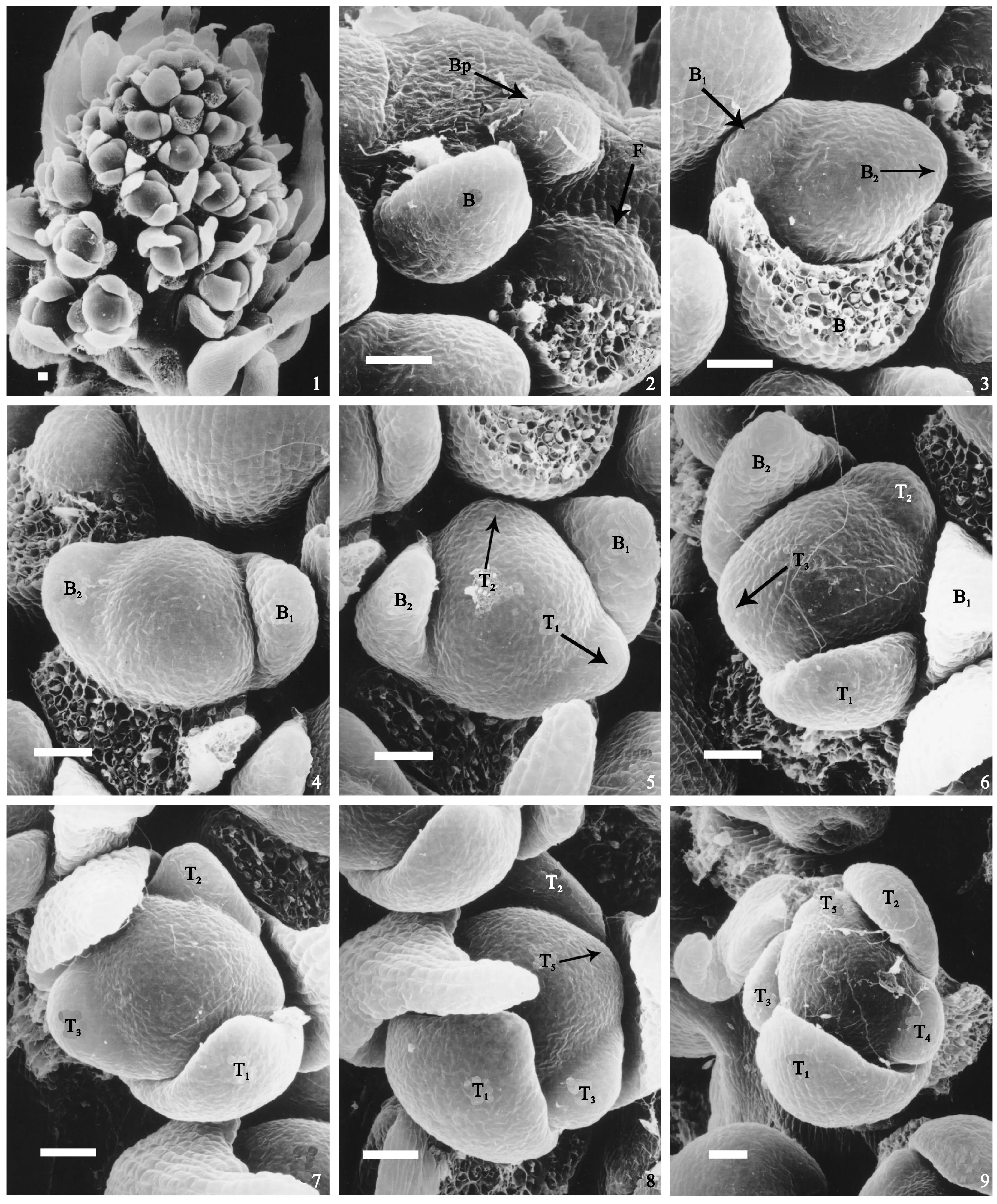
图版Ⅰ 浙江商陆(Phytolacca zhejiangensis)花发生发育过程B. 苞片;Bp. 苞片原基;T. 花被;B1、B2分别代表第1小苞片和第2小苞片;T1、T2、T3、T4、T5分别代表第1、第2、第3、第4、第5花被;标尺=50 μm1. 花序的发育; 2. 花发生(苞片原基首先发生,花原基在苞片的腋部发生);3. 小苞片在花原基两侧发生;4. 正在发育的小苞片,一大一小;5. 第1和第2枚花被发生;6. 第3枚花被发生;图7. 第4枚花被发生;8. 第5枚花被发生;9. 5枚花被都已经发生PlateⅠ Initiation of bracts and sepals in Ph. zhejiangensis.B represents bract; Bp represents bract primordium; T represents tepal; B1 and B2 represents first bractlet, second bractlet; T1, T2, T3, T4, T5 represents first sepal, ..., fifth sepal; Bar = 50 μmFig.1. Developing inflorescence; Fig.2. Initiating flower. The bract primordium initiated firstly, then the floral primordium initiated in the axilla of bract; Fig.3. The bractlets were initiating at the two sides of floral primordium; and the left one is bigger than right one; Fig.4. Developing bractlets, the right one is bigger than left one; Fig.5. The first and the second sepal was initiating; Fig.6. The third sepal is initiating; Fig.7. The fourth sepal was initiating; Fig.8. The fifth sepal was initiating; Fig.9. Five sepals have initiated

图版Ⅱ 浙江商陆(Ph. zhejiangensis)雄蕊和心皮的发生发育过程Sp. 雄蕊原基;S. 雄蕊;So. 外轮雄蕊;T1、T2、T3、T4、T5分别代表第1、第2、第3、第4、第5枚花被;Cp.心皮原基;C.心皮;标尺=50 μm1. 雄蕊在顶端分生组织周围发生;2. 8个雄蕊原基在顶端环状分生组织出现;3. 发育中的雄蕊原基;4. 雄蕊发育,心皮形成;5. 外围的雄蕊在发生发育,心皮原基在环状分生组织上形成;6、7.雄蕊发育中,心皮原基在发育;8、9.心皮生长发育,子房室在心皮内侧形成Sp represents androecium primordium; S represents stamen; So represents outer stamen; T1,T2,T3,T4,T5 represent first tepal, ...,fifth tepal; Cp represents carpel primordium; C represents carpel; Bar = 50 μmPlate Ⅱ. Initiation of androecium and capel of Ph. zhejiangensisFig.1. Androecia are initiating on the edge of apex meristem (ring meristem); Fig.2. Eight androecium primordia appeared on the ring meristem; Fig.3. Developing androecia primordium; Fig.4. Androecia were developing and carpels were forming; Fig.5. Outer stamens were developing and carpel primordia were forming on the ring meristem (edge of apex meristem); Fig.6 and 7. Stamens were developing and carpel primordia were forming and developing; Fig.8 and 9. Carpels were developing and the ovary loculus formed at inside of carpel

图版Ⅲ 红蕊商陆(Ph. esculenta)花被和雄蕊的发生过程察B. 苞片;B1,B2代表第1小苞片,第2小苞片;T. 花被;T1、T2、T3、T4、T5代表第1花被,……, 第五花被片;S. 雄蕊;RM. 环状分生组织;标尺=50 μm1. 发育中的花序;2. 具有2个苞片的花原基;3. 第1和第2花被发生;4. 花原基具有两个明显的花被片;5. 第3个花被原基发生;6. 第5个花被原基发生;7. 5个花被片形成,顶端分生组织呈五边形;图8. 顶端分生组织变成圆形;9. 雄蕊原基在顶端分生组织上形成(顶端分生组织的边缘)Plate Ⅲ Sepals initiated and androecium initiated in Ph. esculentaB represents bract; B1, B2 represents first and second bractlets; T represents tepal; T1, T2, T3, T4, T5 represents first tepal, ..., fifth tepal; S represents stamen; RM represents ring meristem; Bar = 50 μmFig.1. Developing inflorescence; Fig.2. Floral primordium with two bracts; Fig.3. The first and second sepal initiated; Fig.4. Floral primordium with two evident sepals; Fig.5. The third sepal initiated; Fig.6. The fifth sepal initiated; Fig.7. Five sepals were formed and the apex meristem was pentagon; Fig.8. Apex meristem changed to roundness; Fig.9. Androecium primordia were forming on ring meristem (the edge of apex meristem)

图版Ⅳ 红蕊商陆(Ph. esculenta)的雄蕊和心皮发生与发育C.心皮;Ov.胚珠;S.雄蕊;标尺=50 μm1. 雄蕊原基在环状分生组织上发生;2. 心皮原基在环状分生组织上发生;3.发育中的心皮;4. 胚珠在环状分生组织上发生; 5~9. 心皮发育中;7是图8的部分放大Plate Ⅳ Initiation and development of androecia and carpels in Ph. esculentaC represents carpel; Ov represents ovule; S represents stamen; Bar = 50 μmFig.1. Androecium primordia were initiating on ring meristem; Fig.2. Carpel primordial were initiating on ring meristem; Fig.3. Developing carpels; Fig.4. Ovule primordia were initiating on ring meristem; Fig.5-9. Carpel were developing; Fig.7 is a part of Fig.8.
猜你喜欢
山西农业大学学报(自然科学版)(2023年2期)2023-06-23 03:51:04
大众健康(2022年7期)2022-07-30 02:19:04
少年博览·小学高年级(2021年11期)2021-11-21 03:36:55
浙江林业(2021年8期)2021-09-22 21:01:30
知识就是力量(2021年6期)2021-07-09 08:33:12
石河子大学学报(自然科学版)(2021年6期)2021-03-03 01:50:22
新课程·下旬(2018年7期)2018-01-19 11:55:40
生物学教学(2017年1期)2017-02-18 09:11:43
首都食品与医药(2015年13期)2015-11-03 08:54:32
江苏中医药(2015年1期)2015-01-09 05:13:31
