
日光温室CO2加富对番茄叶片光合特性的影响
2018-05-10 08:11袁宏霞张振花郑金英邢国明
江苏农业科学 2018年7期
袁宏霞, 孙 胜, 张振花, 刘 洋, 李 靖, 郑金英, 邢国明
(山西农业大学园艺学院,山西晋中 030801)
番茄(LycopersiconesculentumMill.)为茄科草本植物,喜温喜光且种类繁多,因其营养丰富而备受广大消费者青睐。随着蔬菜产业不断发展,温室番茄栽培面积日益增大,但由于温室密闭结构妨碍了与外界气体交换,造成温室内CO2亏缺严重,进而影响了番茄品质与产量。光合作用是植物生长发育的基础,它不仅与自身因素(叶绿素含量、叶龄等)密切相关,CO2作为光合作用的唯一碳源,对植物光合作用有着重要的影响[1]。有研究表明,CO2浓度增加在引起光合速率增加的同时,会使气孔导度、蒸腾速率降低[2]。而张仟雨等的研究结果表明,大豆鼓粒期气孔导度对CO2浓度增加没有明显反应[3]。邵在胜等在CO2浓度对水稻光合作用影响的研究中也发现,CO2处理对不同生长时期叶片的气孔导度和蒸腾速率的影响不同,拔节期和抽穗期显著降低,而灌浆期的气孔导度和蒸腾速率无明显影响[4]。近年来我国在小麦、大豆、水稻、玉米、甜瓜及黄瓜等众多作物中都开展了CO2加富的研究,但有关高CO2浓度不同叶龄对番茄光合作用影响的研究鲜有报道。植物长期生活在高CO2浓度下会导致光合能力下降的现象称为对CO2的光合适应现象[5]。关于长期高CO2浓度处理对植物光合作用的影响,前人所获得的结果不尽一致。有研究表明,长期高CO2浓度条件下,植物光合速率的促进作用会消失,有的甚至还会降低[6-7]。但也有研究表明,植物长期处在高CO2浓度条件下会促进其光合作用[8]。叶龄是影响植物光合作用的内在因素,不同叶龄对CO2的利用率不同,因此它是反映植物连续生长发育过程的重要指标[9]。本研究通过研究番茄不同叶龄的光合能力对高CO2浓度的响应,进而明确高CO2浓度对植物生长发育的影响,为日后CO2精准施肥提供理论依据。
1 材料与方法
1.1 试验材料与试验设计
本试验在山西农业大学园艺站日光温室中进行。以“兴海12号”番茄为试材。CO2释放采用GMm220传感器自动控制系统(芬兰VAISALA公司和邯郸冀南新区盛炎电子科技有限公司),通过管道和循流风机均匀施气,自动检测温室内CO2浓度、温湿度。温室分为4个隔间,分别通入不同浓度的CO2,以CO2浓度(400±25) μmol/mol为对照(CK),其他处理依次为(600±25)μmol/mol(T1)、(800±25)μmol/mol(T2)和(1 000±25) μmol/mol(T3)。通施时间为每天 08:00—10:30,施肥期间温室密闭,阴雨天不补气。除施用的CO2浓度不同外每个隔间其他条件基本一致。
于2017年3月17日将4叶1心的番茄苗移栽到长7 m、宽0.8 m用黑色地膜覆盖的垄上,双行栽培,株距为0.3 m,行距0.8 m,每个隔间栽216株,缓苗10 d后进行CO2施肥。每个隔间选生长健康、长势基本一致的番茄5株,并做好标记。2017年4月7日,选择每株从下往上第5片真叶进行光合参数及叶绿素测定,之后每隔10 d测定1次,每次均选相同部位叶片,共记录处理50 d的数据。
1.2 方法
1.2.1 番茄光合参数的测定 选择晴天09:00—10:30,用Li-6400(Li-Cor,NE,USA)光合仪测定净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)。并计算水分利用效率(WUE,WUE=Pn/Tr)。测定时使用红蓝光源,设光合有效辐射(PAR)为1 100 μmol/(m2·s),叶温为28 ℃,CO2浓度用小钢瓶控制。
1.2.2 叶绿素含量测定 采用80%的丙酮提取,比色法[10]进行测定。
1.3 数据处理
数据用Excel 2007、SAS软件进行分析。
2 结果与分析
2.1 不同CO2浓度下净光合速率比较
由图1可知,不同CO2浓度下,番茄叶片光合速率先增后减。处理20 d时光合速率最大,T1、T2、T3分别比对照高出23.07%、29.08%、29.36%;20 d后光合速率开始下降,但T2浓度下光合速率高于对照及其他处理。可见一定范围内提高CO2浓度有利于植物光合作用,但不同叶龄对CO2的利用能力有所不同,整个生长阶段以800 μmol/mol浓度更有利于番茄进行光合作用。
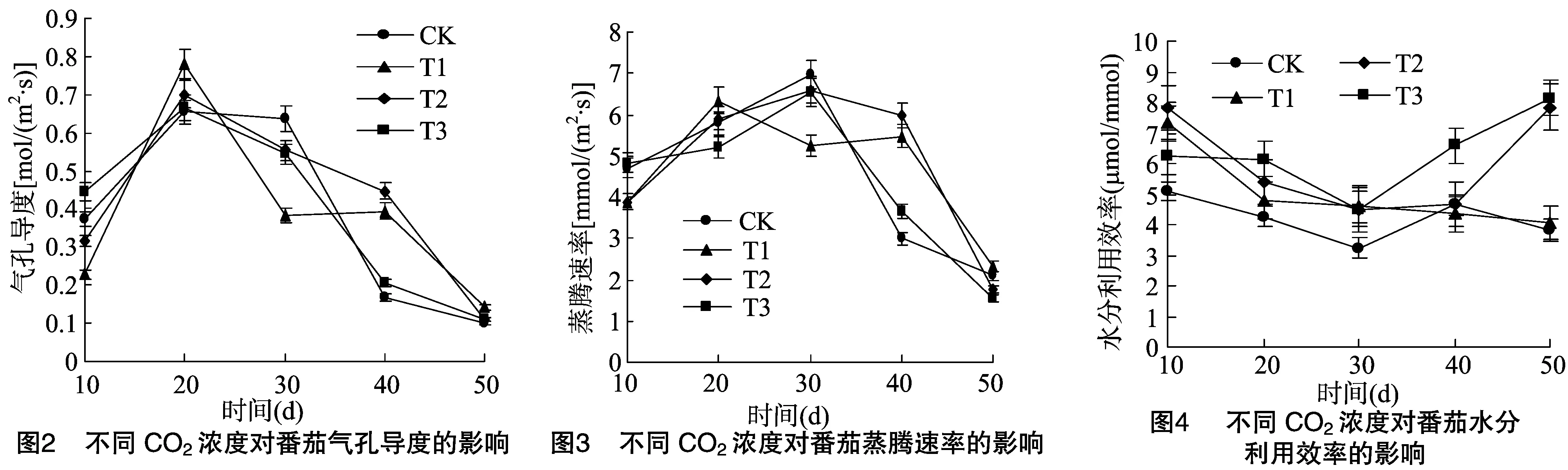
2.2 不同CO2浓度下气孔导度比较
由图2可见,处理10 d时T3条件下气孔导度高于对照,其他处理均低于对照。处理20 d时气孔导度达到最大,各处理均高于对照。随后开始下降,高CO2浓度下降低明显。30 d 后对照及T3条件下气孔导度迅速下降,T1、T2下降缓慢,到 40 d 时气孔导度均高于对照。50 d时各处理及对照气孔导度差异不大。

2.3 不同CO2浓度下蒸腾速率比较
由图3可见,蒸腾速率的大小与气孔导度表现出一致性,即气孔导度大的蒸腾速率也大。但蒸腾速率最大值出现在处理30 d时。这可能与叶面积有关,处理20 d时虽然气孔导度最大但叶面积较小,气孔数目及大小均小于30 d时,导致蒸腾速率较小。
2.4 不同CO2浓度下水分利用效率(WUE)比较
图4表明,水分利用效率随处理天数的增加表现出先降后升的趋势,高浓度下水分利用效率高于对照。除处理10 d时T2条件下最大外,其他时期均以T3条件下最大。30 d时水分利用效率降到最小,且各处理之间几乎无差异。30 d后水分利用效率开始增大,对照和T3处理下增加明显。40~50 d 时T2、T3处理下水分利用效率继续增大且50 d时大小基本相等,而CK和T1条件下水分利用效率略有下降。
2.5 不同CO2浓度下叶绿素含量比较
叶绿素在光合作用光吸收中起着重要的作用。在一定范围内,叶绿素含量与光合能力呈正相关[11]。由表1可知:叶绿素a、叶绿素b及类胡萝卜素含量随着处理天数先增大后减小,处理40 d时最大。叶绿素a在处理10 d和30 d时各处理之间及与对照无明显差异;处理20 d时T2叶绿素a含量显著高于CK和T3,较对照高出13.82%。处理40 d时T1含量达到最大与对照差异显著。可见一定生长时期提高CO2浓度有利于叶绿素a的形成;处理50 d时T3处理下叶绿素a含量较对照低25.24%,且显著低于其他处理。可能是由于高浓度CO2促进了叶绿素a的分解。叶绿素b在前40 d内各处理及对照之间差异不明显,处理50 d后T1含量最低,与T3相比出现极显著差异,说明CO2浓度对叶绿素b含量影响不大,但在叶片生长后期高浓度CO2可能会加快叶绿素b的分解。类胡萝卜素在处理30 d内与对照比较无差异,处理 40 d T1、T2高于对照且存在显著差异;处理50 d T3含量最低,与对照相比差异明显。叶绿素a、叶绿素b总量及叶绿素a/叶绿素b均在处理40 d和50 d后表现出一定的差异。综上,不同CO2浓度在不同叶龄阶段表现出不同程度的影响,较高浓度的CO2有利于嫩叶叶绿素a的积累,但会促进老叶中叶绿素a、叶绿素b及类胡萝卜素的分解,可能与高浓度CO2加快叶片衰老有关。
3 结论与讨论
在植物生长发育过程中,光合作用起到至关重要的作用。本试验对不同CO2浓度下不同叶龄番茄光合作用参数及叶绿素含量进行测定,结果显示,高CO2浓度条件下番茄光合速率均高于对照,这与前人的研究结果[12-13]一致。气孔是植物进行CO2交换的通道,植物主要通过调节气孔开闭来协调对CO2的吸收和水分的消耗,气孔导度作为影响光合作用的重要参数,前人已做了大量研究。王建林等、欧英娟等分别对北方粳稻、龙血树、春羽进行研究,结果表明随着CO2浓度的升高,气孔导度和蒸腾速率表现出降低的趋势[14-15]。张明等的研究结果表明,高CO2浓度条件下大豆光合作用和气孔导度均有增加[16]。胡晓雪等对野生大豆的研究结果显示,气孔导度对CO2浓度升高无显著变化[17]。而梁建萍等对高CO2条件下丁香的研究表明,一定的光合有效辐射,不同的CO2浓度范围内气孔导度变化不一[18]。本研究结果显示,在不同CO2浓度条件下,不同叶龄CO2浓度对气孔导度的影响不同。处理10 d时气孔导度在CO2浓度1 000 μmol/mol下最大且明显高于对照,其他处理下均低于对照。处理20、40 d时各处理气孔导度均高于对照。可能原因是高CO2浓度促进光合酶活性,从而提高植物光合能力,对CO2的需求增加,使气孔通透性增大。处理30 d时各处理气孔导度均低于对照,可能由于CO2浓度过高条件下,植物很容易吸收外界CO2,为了维持胞间CO2分压低于外界,植物会自动调节使气孔关闭,气孔导度降低。蒸腾速率与气孔导度的变化基本相似,随着处理天数的增加,蒸腾速率先增大后减小,在处理 30 d 时达到最大。水分利用效率在处理 30 d 时最小,各处理的水分利用效率均高于对照,可见一定时期提高CO2浓度有利于降低植物蒸腾增大水分利用效率,这与杨克彬等对红掌的研究[19]相符。
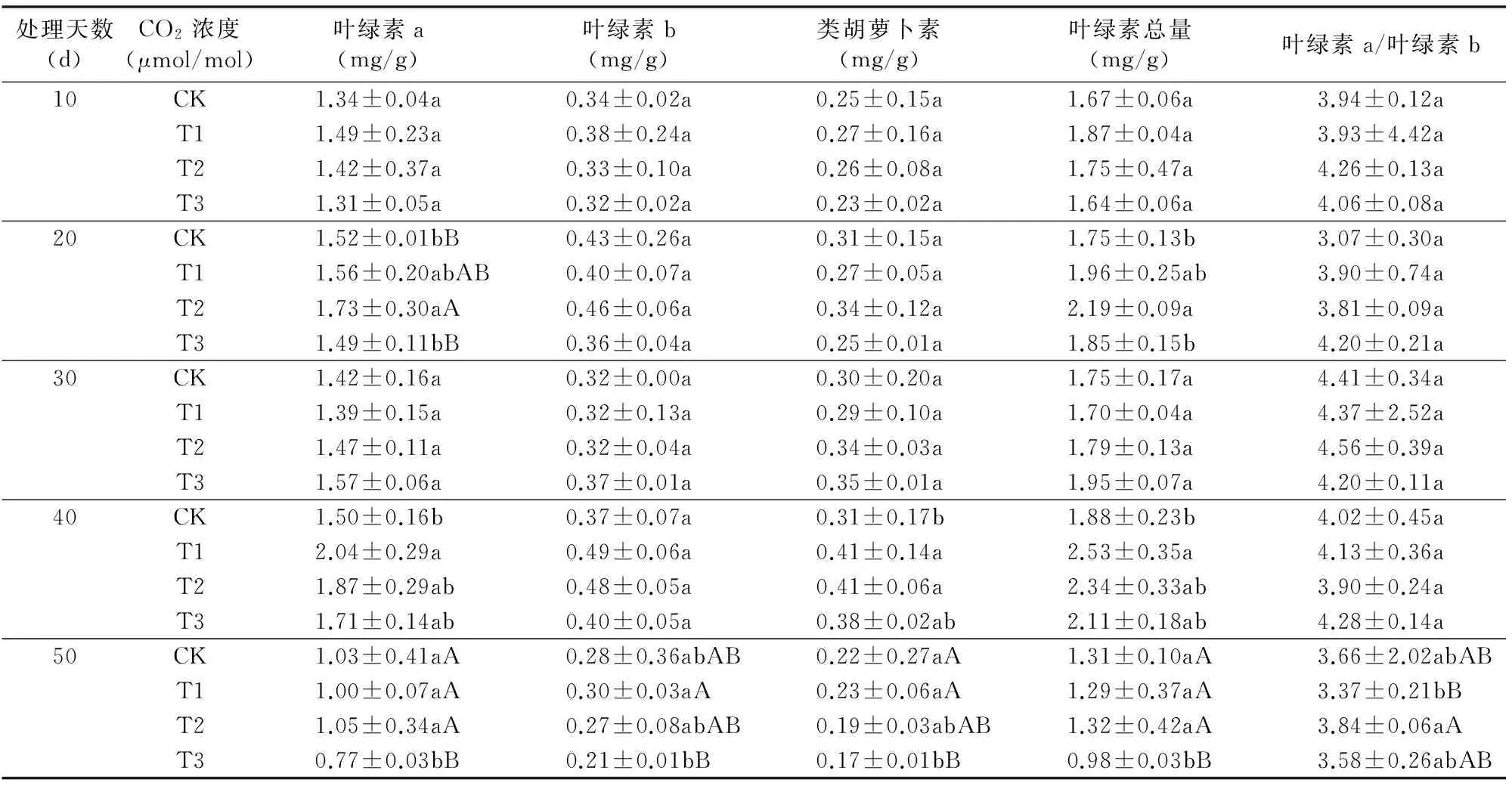
表1不同CO2浓度处理对番茄叶片叶绿素含量的影响
注:同列数据后不同大写字母表示不同处理间在0.01水平上的差异显著,不同小写字母表示在0.05水平上的差异显著。
叶绿素在电子吸收、传递及转化过程中起着重要作用,直接影响植物的光合能力。赵天宏等对大豆的研究发现CO2浓度增大能够提高草本植物叶片叶绿素含量[20];赵甍等对蒙古栎的研究表明,CO2浓度增加在一定程度上促进了叶绿素a的合成,而对叶绿素b及总叶绿素的形成基本无影响[21];赵国锦对番茄的研究表明,高浓度CO2处理,番茄叶片中叶绿素a和叶绿素b含量降低[22]。前人的研究大多只针对部分生长时期,本试验在此基础上进一步研究了不同CO2浓度处理下,番茄叶片从幼叶到老叶整个生长阶段的叶绿素变化,进一步完善前人研究结果。结果表明在一定的生长阶段,适当增加CO2浓度有利于番茄叶片叶绿素a含量的提高;对叶绿素b和类胡萝卜素的影响主要出现在叶片衰老阶段,高浓度CO2条件下二者含量明显低于对照,可见高浓度CO2会促进叶绿素降解,加速叶片衰老。
综上,CO2作为植物光合作用的唯一碳源,不同叶龄对CO2的需求不同,在自然条件无法满足植物光合所需CO2的情况下,提高CO2浓度可以促进番茄光合作用。本试验结果显示CO2浓度(800±25) μmol/mol时番茄光合速率高,叶绿素积累多,水分利用效率大,适宜番茄生长发育。但关于提高CO2促进叶片光合、加速叶片衰老的机理尚不明确,需要进一步深入研究。
参考文献:
[1]叶子飘,于 强. 光合作用光响应模型的比较[J]. 植物生态学报,2008,32(6):1356-1361.
[2]Mamatha H,Rao N K S,Laxman R H,et al. Impact of elevated CO2on growth,physiology,yield,and quality of tomato(LycopersiconesculentumMill) cv. Arka Ashish[J]. Photosynthetica,2014,52(4):519-528.
[3]张仟雨,宗毓铮,董 琦,等. 大气CO2浓度升高对大豆光合生理的影响[J]. 山西农业科学,2016,44(11):1675-1679.
[4]邵在胜,赵轶鹏,宋琪玲,等. 大气CO2和O3浓度升高对水稻‘汕优63’叶片光合作用的影响[J]. 中国生态农业学报,2014,22(4):422-429.
[5]Gunderson C A,Wullschleger S D. Photosynthetic acclimation in trees to rising atmospheric CO2:a broader perspective[J]. Photosynthesis research,1994,39(3):369-388.
[6]韩文军,廖飞勇,何 平. 大气二氧化碳浓度倍增对闽楠光合性状的影响[J]. 中南林学院学报,2003,23(2):62-65.
[7]Chen G Y,Yong Z H,Liao Y,et al. Photosynthetic acclimation in rice leaves to free-air CO2enrichment related to both ribulose-1,5-bisphosphate carboxylation limitation and ribulose-1,5-bisphosphate regeneration limitation[J]. Plant and Cell Physiology,2005,46(7):1036-1045.
[8]周玉梅,韩士杰,张军辉,等. CO2含量升高对水曲柳幼苗净光合与水分利用的影响[J]. 东北林业大学学报,2001,29(6):29-31.
[9]Kikuzawa K,Lechowicz M J. Ecology of leaf longevity[M]. Springer Tokyo,2011,23-30.
[10]Arnon D I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase inBetavulgaris[J]. Plant Physiology,1949,24(1):1.
[11]石 冰,王开运,邹春静,等. 大气CO2浓度和温度升高对草本植物生长的影响[J]. 现代农业科技,2008,(15):15-16.
[12]Cruz J L,AlvesA A C,LeCain D R,et al. Effect of elevated CO2concentration and nitrate:ammonium ratios on gas exchange and growth of cassava (ManihotesculentaCrantz)[J]. Plant and soil,2014,374(1-2):33-43.
[13]王晨光,郝兴宇,李红英,等. CO2浓度升高对大豆光合作用和叶绿素荧光的影响[J]. 核农学报,2015,29(8):1583-1588.
[14]王建林,于贵瑞,王伯伦,等. 北方粳稻光合速率、气孔导度对光强和CO2浓度的响应[J]. 植物生态学报,2005,29(1):16-25.
[15]欧英娟,彭晓春,董家华,等. CO2浓度升高对龙血树和春羽生长及光合生理的影响[J]. 西北植物学报,2013,33(11):2265-2272.
[16]张 朋,张文会,苗秀莲,等. CO2浓度倍增对大豆生长及光合作用的影响[J]. 大豆科学,2010,29(1):64-67.
[17]胡晓雪,杜维俊,杨珍平. 大气CO2浓度和气温升高对野生大豆光合作用的影响[J]. 山西农业科学,2015,43(7):798-801,853.
[18]梁建萍,刘咏梅,牛 远. CO2浓度升高对丁香光合生理特性的影响[J]. 山西农业科学,2007,35(7):10-12.
[19]杨克彬,孟凡志,郭先锋,等. 日光温室冬季增施CO2对切花红掌光合作用及生长发育的影响[J]. 应用生态学报,2017,28(6):1941-1947.
[20]赵天宏,史 奕,王春乙,等. CO2浓度和O3浓度倍增及其复合作用对大豆叶绿素含量的影响[J]. 生态学杂志,2003,22(6):117-120.
[21]赵 甍,王秀伟,毛子军. 不同氮素浓度下CO2浓度、温度对蒙古栎幼苗叶绿素含量的影响[J]. 植物研究,2006,26(3):337-341.
[22]赵国锦. 高浓度CO2对温室番茄光合特性的影响[J]. 北方园艺,2007(9):79-81.
猜你喜欢
生态学报(2024年1期)2024-01-25
科学(2022年4期)2022-10-25
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
中国农业科技导报(2020年12期)2020-03-15
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
