
耦合叶肉导度的陆面过程模型最大叶肉导度参数的敏感性分析
2024-01-25 07:05:16陈美玲袁国富陈报章
生态学报 2024年1期
陈美玲,袁国富,陈报章
1 中国科学院地理科学与资源研究所生态网络观测与模拟重点实验室,北京 100101 2 中国科学院地理科学与资源研究所资源与环境信息系统国家重点实验室,北京 100101 3 中国科学院大学资源与环境学院,北京 100049 4 气象防灾减灾湖南省重点实验室,长沙 410118
陆面过程模型一般采用FvCB模型生物化学光合模型[1-3](Farquhar-von Caemmerer-Berry Model)和气孔导度模型模拟碳同化过程,通常考虑从叶片表面到胞间的气孔导度[4]。已有研究表明,在陆面过程模型光合模块中添加从胞间到羧化位点的叶肉导度,能有效改善陆面过程模型模拟总初级生产力(GPP,Gross Primary Production)对大气CO2增加的响应(CO2施肥效应)的精度[5-7]。当前陆面过程模型中添加叶肉导度的方法最常用的是连乘模式[5-6,8-10],由影响叶肉导度的不同环境因子影响函数的乘积再乘以最大叶肉导度参数计算获得。除不同环境因子影响外,最大叶肉导度参数的取值也会显著影响叶肉导度模拟结果,从而影响陆面过程模型模拟CO2施肥效应的精度。目前的陆面过程模型叶肉导度模拟相关研究还较少探讨最大叶肉导度参数取值变化对叶肉导度和CO2施肥效应模拟结果的影响。
最大叶肉导度代表了基因或者遗传对叶肉导度的影响[5-6,11],是叶肉导度计算方法中的重要参数。一般将25 ℃、饱和光条件下完全展开的成熟叶片中叶肉导度最大值定义为植物叶片最大叶肉导度(gmmax25,Maximum Mesophyll Conductance)。为了对模型中gmmax25进行参数化,不同研究人员对gmmax25的取值有不同的选择。部分研究认为gmmax25与叶片结构有关,是植被功能型特征之一,gmmax25的极限由叶片结构决定[12],决定gmmax25最典型的叶片结构为比叶面积(叶片面积/叶片干重,SLA),Sun等[5]将其用于CLM4.5模型中gmmax25的计算。然而有研究认为SLA并不能完全解释gmmax25的重要变化[13-14],因为SLA集成了可能对gm产生影响的几个不同叶子特性[11,15]。Suits等[10]使用最大羧化速率(Maximum Carboxylation Rate,Vcmax)来表现gmmax25的参数特征,Knauer等[6]利用叶水平测量来对JSBATCH模型中gmmax25进行参数化,相对来说避开了使用植被特征参数的不合理性。Knauer等[7]认为gmmax25不太可能由单一植被特征来解释其变化,目前并未有一个公认且合理的对gmmax25参数化的方法。对于决定gmmax25取值的影响因素也存在争议,叶水平的测量结果表明,gmmax25不仅在各植被功能型之间存在较大差异,在同一种植被功能型中也存在较大误差[6],甚至在同一种植物中叶肉导度存在差异如阴叶的最大gmmax25低于阳叶。gmmax25值会因为测量方法的不同产生不确定性[12],导致在实际应用中同一植被功能型的取值存在较大差异[5-6]。由于gmmax25取值的不确定性,对陆面过程模型叶肉导度模块的gmmax25进行参数优化讨论很有必要。
本文以国际上重要陆面过程模型之一--EALCO模型为例,通过人为设定不同gmmax25值,模拟叶肉导度和GPP,分析和揭示叶肉导度和GPP模拟对gmmax25的敏感性。并与已有研究相关结果比较,以期探讨陆面过程模型中gmmax25参数优化途径,为陆面过程模型改进叶肉导度模拟提供重要参考。
1 材料与方法
1.1 观测和驱动数据
本研究以美国哈佛森林生态监测站(Harvard Forest Environmental Monitoring Site,US-Ha1站)(42.538°N,72.171°W)典型温带落叶阔叶林为研究对象,位于美国麻省(Massachusetts)哈佛森林地区,海拔340 m。植被类型以红橡树(Quercusrubra)和红枫树(Acerpalmatum‘Atropurpureum’)为优势种的落叶阔叶林为主[16],冠层高度30 m,树龄80-110年[17]。土壤类型以多石的沙质土壤为主。2002-2012年1月平均气温为-4.31 ℃,7月平均气温为19.80 ℃。
EALCO模型驱动数据和验证数据来自FLUXNET2015小时步长数据,线性插值成半小时。土壤数据来自国家青藏高原科学数据中心全球土壤数据[18](GSDE,The global soil dataset for earth system modeling),共8个垂直分层,最深为2.3 m,原始数据为0.1°×0.1°空间分辨率(约10 km),提取信息包括沙土、陶土、土壤有机质含量、土壤容重、15 cm土壤碳含量、土壤总碳含量。坡度数据来自地理空间数据云[19],ASTER GDEM 30 m分辨率数字高程数据,通过ArcGIS软件工具进行坡度提取。LAI数据来源于国家地球系统科学数据中心[20],2011-2012年GLASS产品叶面积指数LAI MODIS,数据集分辨率为1 km。地下水位数据来自USGS国际水系统站点实测数据[21]。根分布数据来自Jackson等[22],根分6层,随土壤深度变化。叶片碳氮比来自Zhou等[23]。以上模型参数本地化数据见表1。
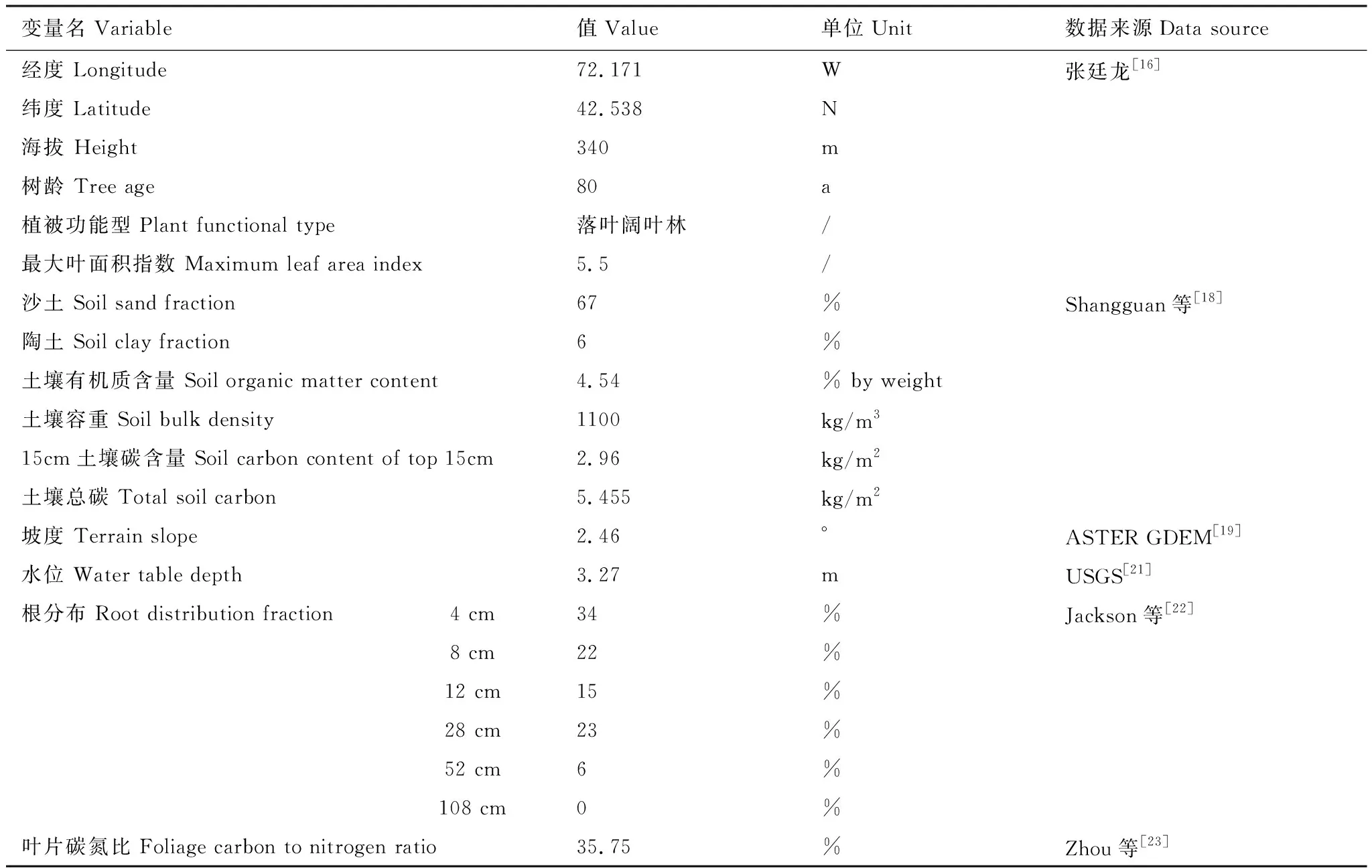
表1 模型参数本地化数据Table 1 Model parameter localization data
1.2 EALCO模型介绍
EALCO模型,全名为Ecological Assimilation of Land and Climate Observations,由加拿大遥感中心Shusen Wang开发[24],其前身是加拿大气候中心Diana L Verseghy开发的CLASS (Canadian Land Surface Scheme)模型[25-26]。该模型包含5种地表覆盖类型:(1)有植被,地表无雪覆盖;(2)有植被,地表有雪覆盖;(3)无植被,地表无雪覆盖;(4)无植被,地表有雪覆盖;(5)水体[27]。模型全面模拟了地表水碳循环的各个过程,包括蒸散发(冠层蒸腾、冠层蒸发和地表蒸发)、冠层雨雪的截留、水分在土壤以及雪层中的传输过程(包括冻融过程)、地表径流、雪覆盖动态变化、水分在根-茎-叶-大气中的传输过程以及土壤水分下渗与地下水的补给水循环过程[27];在碳循环过程中计算了植物光合作用、自养呼吸、根系氮吸收、凋落物、植物碳氮分配和生长、土壤异养呼吸等,模拟植物、土壤和生态系统CO2交换。在碳分配过程中,将碳分配给叶、茎杆粗根和细根,分为基质碳和结构碳;在植被模块,实现胞间CO2浓度、大气CO2浓度以及叶片表面CO2浓度动态平衡模拟。
1.3 EALCO模型叶肉导度耦合方法
1.3.1叶肉导度模型
本文中采用的叶肉导度算法主要参考Sun等[5]和Knauer等[6],组合四个影响因子进入EALCO模型中,分别为:氮分布影响函数f1(N)、温度响应函数f2(Tl)、水分胁迫函数f3(φc)和光照影响函数f4(Qa)。其公式如下所示:
gm=gmmax25f1(N)f2(Tl)f3(φc)f4(Qa)
(1)
式中,gm为叶肉导度(Mesophyll Conductance)(mol m-2s-1);gmmax25为最大叶肉导度(mol m-2s-1),是叶肉导度模拟的主要参数之一。氮分布影响函数f1(N)选自Knauer等[6]中的冠层氮分布廓线,分为阴叶f1(NB)和阳叶f1(ND)。公式如下:
f1(ND)=exp(-0.11*sunLAI)
(2)
f1(NB)=exp(-0.11*shaLAI)
(3)
式中,sunLAI表示为阳叶的叶面积指数(m2/m2);shaLAI表示为阴叶的叶面积指数(m2/m2)。
温度响应函数f2(Tl)和水分胁迫函数f3(φc)均取自EALCO模型[28]:
(4)
式中,Tl是叶片温度(K);A为经过参数本地化后的值,无量纲量;R是理想气体常数(J mol-1K-1);S是熵变(J mol-1K-1);Ha是活化能(J/mol);Hdl是低温下的失活能(J/mol);Hdh是高温下的失活能(J/mol)。
(5)
式中,φc是植被叶水势(MPa);φc,min和φc,max代表光合作用冠层水势的零点和阈值点。方程(4)和(5)中参数取值参见文献[29]。
光照影响因素f4(Qa)取自Knauer等[6]:
f4(Qa)=1-(1-fmin)e-0.003Qa
(6)
式中,Qa是光量子通量(W/m2),需要对光合有效辐射(PAR,Photosynthetically Active Radiation)(W/m2)进行转换[30]得到。fmin是经过大量数据拟合获得的参数,取值为0.15[31-33]。
1.3.2顾及叶肉导度的光合作用模型
EALCO模型中光合作用模拟采用FvCB模型,仅考虑气孔导度,未考虑叶肉导度[24]。FvCB模型考虑叶肉导度计算光合速率一般须将光合模型中的胞间CO2浓度(Ci,Intercellular Carbon Dioxide Concentration)替换成羧化位点CO2浓度(Cc,Carbon Dioxide Concentration at the site of Carbonxylation),在此基础上对光合模型参数进行重新优化调整。
加入叶肉导度后的暗反应光合速率计算公式更改成为[6]:
Vd,c=Vcc,max(Cc-τc)/[Cc+kc,c(1+Oc,c/ko,c)]
(7)
式中,Vd,c为考虑叶肉导度的暗反应阶段光合速率(μmol m-2s-1);Cc为羧化位点CO2浓度(Pa);其余符号为考虑叶肉导度后重新调整的光合参数,其中Vcc,max为羧化速率(μmol m-2s-1);τc为光合补偿点(Pa);kc,c为1,5二磷酸核酮糖羧化酶(Rubisco,Ribusco-1,5-Bisphosphate Carboxylase/Oxygenase)羧化的米氏常数(Pa);ko,c为Rubisco氧化的米氏常数(kPa);Oc,c为叶绿体部位O2浓度或分压(kPa)。
加入叶肉导度后的光反应光合速率计算公式更改为[6]:
Vl,c=J(Cc-τc)/(εCc+10.5τc)
(8)
式中,Vl,c为考虑叶肉导度的光反应阶段光合速率(μmol m-2s-1),J和ε分别是电子传输速率和CO2固定所需的电子,取值依照EALCO模型[29]。
1.3.3基于Cc的光合模型参数优化方法
添加叶肉导度后的光合模型参数优化调整,采用Sun的方法[34],利用净光合速率(An,Net Photosynthetic Rate)/胞间CO2浓度曲线(An/Ci曲线)进行拟合估计。具体方法是选取一组米氏常数(Kc、Ko)值,采用最小二乘法对原模型与添加叶肉导度后新模型进行拟合,通过演化算法[35]求得Vcmax和最大电子传输速率(Jcmax,Maximum Potential rate of Electron Transport)的全局最优解。包括三步:第一步,根据原FvCB模型模拟一条在适宜温度(25 ℃)、饱和光(1200 μmol m-2s-1)、无胁迫下An/Ci曲线,Ci取值范围为1-150 pa。第二步,将gmmax25取一固定值,Ci和An取自第一步中的结果,根据Fick第一定律Cc=Ci-An/gm,将Cc代入FvCB模型中,求新模型的光合速率,拟合出一条新的在适宜温度(25 ℃)、饱和光(1200 μmol m-2s-1)、无胁迫下An/Ci曲线(即An/Cc曲线)。第三步,将第一步和第二步生成的两条光合响应曲线进行拟合,手动调整Kc和Ko,进行非线性规划(最小二乘法)最优求解,获得新的考虑叶肉导度的光合模型参数Vcmax和Jmax。
1.3.4EALCO模型中叶肉导度的模拟实现
EALCO模型中叶肉导度的模拟实现过程见图1所示,分为三步:第一步,给Cc设定一个初值,Cc=Ci,采用优化后的光合参数(Kc、Ko、Vcmax和Jmax)参与FvCB光合模型过程。第二步,计算气孔导度和叶肉导度(公式(1))。第三步,将计算好的gm,与前两步计算的光合速率、气孔导度带入Fick第一定律中Cc=Ci-Angm,求得羧化位点CO2浓度。通过循环控制条件[29]:

图1 EALCO模型中叶肉导度的嵌入过程Fig.1 Embedding process of mesophyll conductance in EALCO model Cc:羧化位点CO2浓度;Ac:EALCO模型中的光合作用速率;gc:气孔导度;gm:叶肉导度;gmmax25:最大叶肉导度;Ci:胞间CO2浓度;An:净光合作用速率;RCO2:呼吸作用速率;Ca:大气中CO2浓度;εC:CO2的循环控制条件判断项,约为10-2An;ra:边界层阻力;rl:气孔阻力(1/gc)
(9)
根据判断函数εc<εC是否应跳出循环。
1.4 敏感性分析方法
本研究主要通过在模型中人为设置不同的gmmax25值,分析模型输出叶肉导度和GPP的变化,以模拟试验的方法,探讨模型叶肉导度和GPP模拟对最大叶肉导度取值的敏感性。由于gmmax25、叶肉导度的实际观测值大多集中在实验室条件,自然状况下的参考数据较少,而模型模拟以自然环境为驱动,模拟结果很难通过对应的实际观测数据加以检验和分析。因此本研究与现有相关研究结果进行比较,从以下四个方面对模型模拟叶肉导度和GPP对gmmax25取值变化的敏感性加以分析:
首先,与实际观测GPP比较,分析实际观测值与添加叶肉导度后的模拟值之间拟合统计参数决定系数(R2,The Coefficient of Determination)和均方根误差(RMSE,Root Mean Squared Error)的变化,揭示不同gmmax25取值下模型模拟GPP精度变化,并基于此探讨最大叶肉导度可能取值范围;
其次,研究表明叶肉导度与气孔导度之间存在一定的线性比例关系[36-38],通过拟合不同gmmax25取值下叶肉导度与气孔导度的线性关系,分析线性斜率变化特征,并与相关研究结果比较,探讨最大叶肉导度的可能取值范围;
再次,本研究分析不同gmmax25取值下模型模拟的CO2施肥效应,与相关研究比较,探讨最大叶肉导度的可能取值范围;
最后,根据公式(1)可知,当环境要素相对稳定时,叶肉导度随gmmax25取值线性增加,斜率意味着叶肉导度模拟对最大叶肉导度取值的敏感性。本研究分析了月尺度上的gmmax25与模型模拟叶肉导度的线性关系,从模型分析的角度揭示最大叶肉导度取值的时间(季节)敏感性。
研究的最终目的是希望为模型最大叶肉导度参数取值提供优化路径和思路,为陆面过程模型添加叶肉导度提供重要参考。
2 结果
2.1 最大叶肉导度不同取值对GPP模拟精度的影响
本研究参考了Knuaer等[6]文中gmmax25在0.04-1.21 mol m-2s-1取值和Sun等人[5]文中gmmax25在0.1-0.247 mol m-2s-1取值,针对US-Ha1站点落叶阔叶林,考虑最终模拟结果特征,最大范围的选取了gmmax25值,分别为0.175、0.35、0.5、0.75、0.8、1、3.5 mol m-2s-1。
模型模拟的默认条件是未进行基于Cc的光合参数优化方法这一步骤,以2011年观测数据为例加以分析。对半小时GPP模拟值与观测值进行线性拟合,图2显示了随gmmax25取值不同,添加叶肉导度后的半小时输出GPP模拟结果与通量观测值之间的决定系数R2和均方根误差RMSE的变化,可以看出gmmax25从0.175、0.35、0.5、0.75、0.8、1.0到3.5变化,R2逐渐增大趋近1,RMSE逐渐减小。总体上,当gmmax25小于0.5 mol m-2s-1时,相关性较差,RMSE较大,且决定系数变化率较大;当gmmax25从1.0 mol m-2s-1到3.5 mol m-2s-1时,相关性较好,RMSE较小,且RMSE和相关性变化率较小。这个结果表明gmmax25取值越大,GPP模拟整体精度越高,但gmmax25达到一定值后,模拟精度的改善空间有限;同时gmmax25值越低,GPP模拟整体精度会快速降低。

图2 EALCO模型中默认条件下添加叶肉导度后GPP模拟与通量估算拟合优度Fig.2 Goodness of fit metrics between GPP simulation in EALCO under default condition with mesophyll conductance and flux estimationR2:决定系数;RMSE:均方根误差
植物叶片gmmax25反映了植物的生理和结构特征,存在合理的取值范围,不能为提高GPP模拟整体精度随意提高gmmax25值。综合考虑本研究结果体现出的模拟精度需要,以及gmmax25可能的合理取值范围,在后续处理gmmax25取值时,本研究将gmmax25取值分别设置为0.5、 0.57、 0.7、0.8、1.0、1.2 mol m-2s-1进行分析。
2.2 不同最大叶肉导度取值下叶肉导度与气孔导度关系
以2011年全年数据为例,模拟了阳叶叶肉导度与气孔导度,并分析了二者之间的关系。图3是gmmax25分别取值0.5、0.57、 0.7、 0.8、1.0、1.2 mol m-2s-1时,EALCO模型模拟的半小时阳叶叶肉导度和阳叶气孔导度的散点图,线性拟合关系及相应的统计数据。
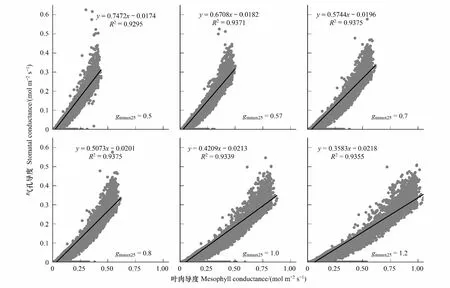
图3 不同gmmax25取值下模拟叶肉导度与气孔导度的线性关系Fig.3 The linear relationship between simulated mesophyll conductance and stomatal conductance under different gmmax25 values
模拟结果显示如下特征:(1)气孔导度模拟值的变化范围不受gmmax25值变化影响,基本维持在0-0.5 mol m-2s-1范围内,叶肉导度模拟值随gmmax25值增大而扩大,从0-0.45 mol m-2s-1(对应gmmax25取值0.5 mol m-2s-1)到0-0.9 mol m-2s-1(对应gmmax25取值1.2 mol m-2s-1)。这个结果说明EALCO模型模拟气孔导度基本独立于叶肉导度过程,不受叶肉导度变化的影响。(2)气孔导度与叶肉导度呈现很好的线性关系(R2均大于0.9)。考虑到气孔导度模拟与叶肉导度模拟可以近似看成两个独立过程,这种拟合的线性关系印证了二者之间的线性关系。由于拟合出的线性函数的截距值均较小(在0.01-0.02之间),线性关系曲线的斜率表明气孔导度与叶肉导度之间存在固定的比例关系。这一模拟结果与前人文献中实验观测结果相似[36,39-40],即植物叶片的叶肉导度和气孔导度之间整体存在固定比例关系。(3)气孔导度与叶肉导度的固定比例关系随gmmax25取值变化而变化。gmmax25分别取值0.5、0.57、0.7、0.8、1.0 、1.2 mol m-2s-1时,对应的气孔导度与叶肉导度的比值(即拟合曲线的斜率)分别是0.7472,0.6708,0.5744,0.5073,0.4209和0.4177。gmmax25取值越高,气孔导度与叶肉导度比值越小。比值一直小于1也表明气孔导度值小于叶肉导度值。
2.3 不同gmmax25取值下模拟GPP对CO2浓度升高的响应
本研究分别模拟了大气CO2初始浓度为300、350、400、450 μmol/mol时,升高100 μmol/mol后,模型模拟GPP增长率的差异。其中GPP模拟以US-Ha1站点落叶阔叶林树种2002-2012年共11年数据进行模拟获得多年平均GPP,GPP增长率通过大气CO2增加100 μmol/mol后的多年平均GPP减去初始大气CO2浓度下的多年平均GPP再除以初始大气CO2浓度下的多年平均GPP获得。
表2展示了gmmax25分别取值0.5、0.57、0.7、0.8、1.0、1.2 mol m-2s-1时的GPP增长率变化与未添加叶肉导度下GPP增长率变化的比较。
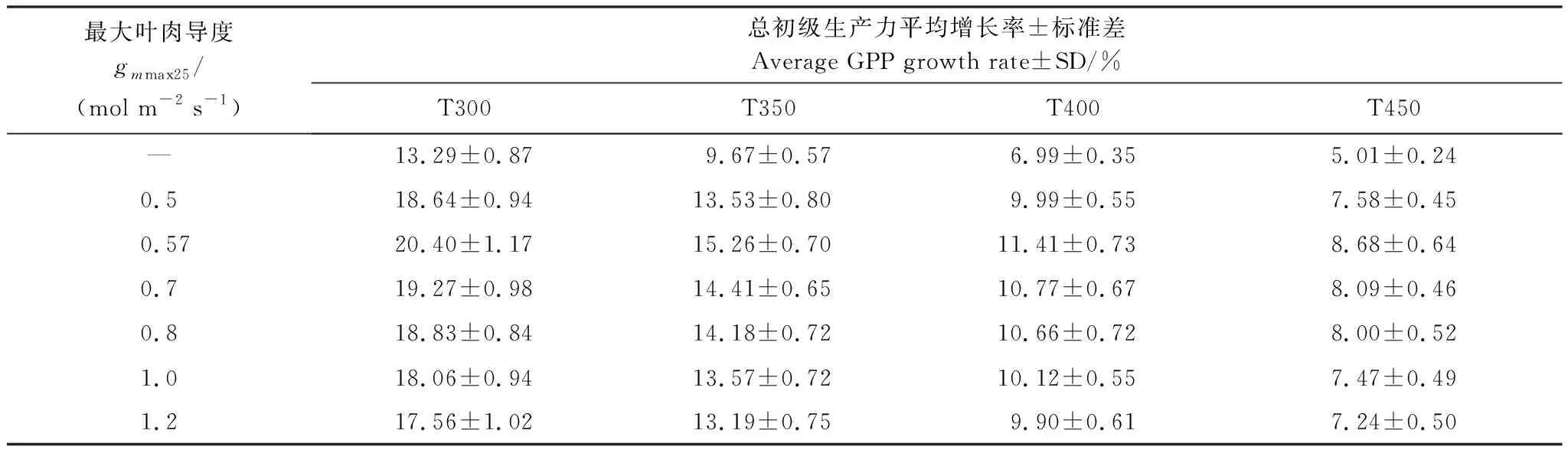
表2 不同gmmax25取值下EALCO模型模拟不同CO2浓度增幅范围的GPP平均增长率(2002-2012年平均值)和标准差Table 2 Average GPP growth rate and Errors in EALCO model with different gmmax25 values from 2002 to 2012 under different ranges of CO2 concentration increment
结果显示模型添加叶肉导度后,模拟GPP对大气CO2浓度上升的响应呈现如下特征:(1)同未添加叶肉导度相比,添加叶肉导度后模型模拟GPP对大气CO2升高的增长率普遍增加;(2)随着初始CO2浓度升高,GPP随CO2增加的增长率下降,这一趋势不管是否添加叶肉导度均存在;(3)随着gmmax25值的增加,GPP增长率没有呈现单一的变化趋势。当gmmax25取值为0.57 mol m-2s-1时GPP增长率普遍高于其他最大叶肉导度取值时的GPP增长率;gmmax25取值高于0.57 mol m-2s-1后,随gmmax25取值增加,GPP增长率呈下降趋势;(4)不同gmmax25取值下GPP增长率的变化幅度差异(最大GPP增长率与最小GPP增长率之差)不大。在T300范围时,最大变化幅度差异为2.34%;T350范围时,最大变幅差为1.73%;T400范围时,最大变幅差为1.42%;T450范围时,最大变幅差为1.21%。上述变幅差仅比GPP增长率的标准差略高,显示gmmax25的不同取值对GPP增长率的模拟结果影响有限。
2.4 叶肉导度模拟对gmmax25取值的季节敏感性
在月尺度上,环境因子的影响相对稳定,叶肉导度对gmmax25取值更为敏感。本文计算了生长季月平均叶肉导度,以2011年生长季(3-11月)为例(图4),可以发现如下特征:(1)在月尺度上叶肉导度随最大叶肉导度的增加而线性增加,与gmmax25显著线性相关(R2>0.98),这个现象也表明在月尺度上光照、温度、水分和氮含量等环境因子对叶肉导度的综合影响相对稳定。(2)考虑叶肉导度与gmmax25线性关系曲线方程的截距均较小,方程的斜率体现了gmmax25取值对叶肉导度模拟值的影响程度,斜率越大意味着gmmax25取值变化对叶肉导度模拟结果影响越大,或叶肉导度模拟对gmmax25取值越敏感。可以发现不同月份的曲线斜率不同,表明叶肉导度模拟对gmmax25取值敏感度在不同月份存在差异。叶肉导度与gmmax25线性关系的曲线方程斜率变化特征是:从3月份的0.0742逐月增加到7月份,最大为0.4479,之后逐月下降,至11月为0.1859,生长季内逐月变化趋势呈单峰状。其中7、8月份的斜率大于0.4,5、6、9月份的斜率在0.3-0.4之间,4、10月份的斜率在0.2-0.3之间,11月份斜率在0.1-0.2之间,3月份斜率小于0.1。这个结果表明生长盛期的7、8月份gmmax25取值对叶肉导度模拟结果影响最大,或者说生长盛期叶肉导度模拟对gmmax25取值最为敏感,其次是5、6、9月份等生长次盛期,其他月份的影响较小。

图4 月平均叶肉导度随gmmax25的变化特征(以2011年为例)Fig.4 Characteristics of monthly mean mesophyll conductance changed by gmmax25 (such as in 2011)
3 讨论
3.1 陆面过程模型添加叶肉导度模拟的权衡
目前的陆面过程模型模拟GPP对大气CO2浓度升高的响应明显偏低,大部分模型模拟GPP对CO2浓度每升高100 μmol/mol,GPP增长率一般在5%左右,远远低于FACE(Free-Air CO2Enrichment)实验观测的15.5%的增长率[41]。在EALCO模型中添加叶肉导度后,GPP对CO2升高100 μmol/mol的响应与实验观测结果更接近,证实了模型添加叶肉导度能提高CO2施肥效应的模拟精度。
另一方面,图2的结果表明叶肉导度越大(或叶肉阻力越小),模型模拟GPP整体精度越高,意味着模型只要考虑叶肉导度就会影响GPP模拟整体精度。在不改变模型其他参数的情形下,在陆面过程模型中添加叶肉导度对GPP模拟的影响是相反的,一方面添加叶肉导度会降低GPP模拟的整体精度。另一方面添加叶肉导度会提高模型模拟CO2施肥效应的精度。由于本研究向EALCO模型中添加叶肉导度时除调整了光合模块的参数外,并未进一步考虑模型其他模块参数的调整。模型的碳、氮、水循环过程模拟是相互耦合的,在调整了光合参数后其他参数如果不加以调整,势必会影响GPP模拟的结果和精度。从本研究的模拟结果看,的确降低了GPP模拟整体精度。叶肉导度的添加意味着叶片碳交换过程增加了叶肉阻力项,导致GPP模拟比未添加叶肉导度时低是可以预期的。尽管添加叶肉导度提高了模型模拟GPP应对CO2升高的响应精度,但GPP绝对值模拟结果的降低仍然是不可接受的。我们认为,陆面过程模型添加叶肉导度模拟在调整光合模型参数的同时,应该调整和优化其他模拟过程的参数值,以尽量维持GPP模拟的整体精度。如何同时调整和优化模型其他参数是陆面过程模型考虑叶肉导度模拟需进一步关注的环节。
3.2 本案例中最大叶肉导度的合理取值范围
不同植被功能型的最大叶肉导度作为模拟叶肉导度的关键参数,它的合理取值没有明确的实验数据支撑,目前的相关应用中对gmmax25的取值也存在差异[5,6],但陆面过程模型中不同植被功能型gmmax25的合理取值对于叶肉导度模拟至关重要。在本研究中,分别分析了不同gmmax25取值对叶肉导度、GPP模拟结果的影响,以及在不同gmmax25取值下模拟的叶肉导度与气孔导度之间的关系、GPP对大气CO2浓度升高的响应差异等。将这些结果与过往的相关研究或实验观测结果对比,可以大致推测出本案例(即针对US-Ha1台站落叶阔叶林在EALCO模型)中的gmmax25合理取值范围。
李勇等[38]归纳的有关叶肉导度观测实验数据表明叶肉导度值整体在0-0.6 mol m-2s-1范围内变化。从图4可以看出,当gmmax25取值达到0.8以后,部分叶肉导度模拟值超出了0-0.6 mol m-2s-1这一范围。依据李勇等[38]的归纳结果,为确保叶肉导度值在0-0.6 mol m-2s-1之内,gmmax25取值小于0.8 mol m-2s-1较为合理。
气孔导度与叶肉导度的关系多与内部水分利用效率存在联系[37,39],大量研究表明气孔导度与叶肉导度之间存在明显的线性关系。有研究认为气孔导度与叶肉导度保持在0.79的稳定比例[39],也有统计研究表明,气孔导度与叶肉导度之间存在1:1的比例[38]。本试验模拟结果证实了气孔导度与叶肉导度之间的线性关系,其比值受最大叶肉导度的影响,随gmmax25取值增加而下降。当gmmax25取 0.5-0.7 mol m-2s-1之间时,气孔导度与阳叶叶肉导度的比例大于0.57,超过0.8以后比值已低于0.5,与已有研究总结的比值相差较大。
当gmmax25取值越低时,模拟GPP精度显著下降(图2),暗示gmmax25取值的最低值不能过低。综上所述,在本案例中,当gmmax25取值范围在0.5-0.7 mol m-2s-1之间时,既能获得较合理的GPP模拟精度,又能使得模拟的叶肉导度值范围在合理范围之内,使得气孔导度与叶肉导度之间的比值更为合理。我们据此推断0.5-0.7 mol m-2s-1是本案例gmmax25参数的合理取值范围。
3.3 陆面过程模型最大叶肉导度参数优化途径
鉴于最大叶肉导度取值对陆面过程模型叶肉导度和GPP模拟的显著影响,陆面过程模型中添加叶肉导度需要对该参数进行优化,以确保更合理的模拟结果。根据本研究的分析,提出陆面过程模型最大叶肉导度参数优化的途径:
首先分析和判断模型模拟获得的叶肉导度值是否在合理范围之内。目前已经有大量的实验观测数据给出了不同植被功能型叶肉导度的变化范围,当所选gmmax25值模拟获得的叶肉导度变化范围超出实验观测叶肉导度值时,那么可认为选择的gmmax25值是不合理的。
其次可将模拟的气孔导度与叶肉导度比值与既有研究进行比较。目前的文献中气孔导度与叶肉导度比值在0.7-1.0之间,当二者比值小于这一范围时,需谨慎调整gmmax25值大小。
另外还应分析GPP模拟精度对gmmax25取值的敏感性,gmmax25取值范围应降低gmmax25对GPP模拟精度的影响。
4 结论
(1)gmmax25直接影响叶肉导度模拟值,在月尺度上呈显著线性关系,gmmax25值越大,叶肉导度模拟值越大。这种影响在生长季不同生长期不一样,在7、8月份的生长盛期,叶肉导度模拟对gmmax25值最敏感,其次是5、6和9月的次生长盛期,4、10月的敏感度继续降低,在3月和11月叶肉导度模拟对gmmax25值的敏感度最低。
(2)模拟试验结果证实:陆面过程模型添加叶肉导度有助于改善CO2施肥效应精度,但gmmax25的不同取值对施肥效应模拟精度并不随取值差异一致性变化。在本案例中,gmmax25取值在0.5-0.57之间时,施肥效应响应敏感度(GPP增长率)随gmmax25值增加而增加,但大于0.57后,敏感度随gmmax25值增加而下降。这一规律是否在其他植被功能型存在,或者在其他陆面过程模型中存在,还需要进一步研究,其内在机理也需要进一步探讨。
(3)通过将模型模拟的叶肉导度的变化范围与实验观测到的叶肉导度变化范围进行比较,将模型模拟的气孔导度与叶肉导度比值与实际观测比较,可以初步判断最大叶肉导度合理取值范围,是陆面过程模型最大叶肉导度参数优化的重要途径。
致谢:感谢加拿大遥感中心高级研究员王树森老师和福建师范大学副教授陈耀亮老师在EALCO模型研究中给予的帮助。
猜你喜欢
农业工程学报(2022年3期)2022-04-16 03:19:44
蔬菜(2022年12期)2022-01-01 11:32:10
星星·散文诗(2021年34期)2021-12-21 10:39:25
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
节水灌溉(2020年9期)2020-09-16 00:32:12
发明与创新·大科技(2020年1期)2020-04-17 09:01:39
北京大学学报(自然科学版)(2020年2期)2020-03-26 05:12:12
农村百事通(2019年1期)2019-01-30 13:36:34
水利水电快报(2018年10期)2018-11-05 08:48:30
