
南方丘陵区油茶气孔导度模型修正
2022-04-16 03:19:44陈永忠岳伶俐吴友杰
农业工程学报 2022年3期
夏 雄,陈永忠,岳伶俐,马 彬,吴友杰※
(1. 湖南农业大学水利与土木工程学院,长沙 410128;2. 湖南省林业科学院,长沙 410004)
0 引 言
近年来全球气候不断变化,中国南方丘陵区旱灾较频发,严重制约了农业可持续发展。油茶是中国南方重要的农林经济作物,具有很高的营养价值和保健功能,能够缓解耕地资源压力,带动产业发展。叶片气孔导度是植物与外界环境进行水分和气体交换的主要通道,调节着植物体的光合作用与蒸腾作用,是油茶水分损失和能量平衡的重要指标,在复杂的气候环境条件下,气孔可通过膨压变化感知环境的改变,从而在时间和空间上有不同的开度。然而针对中国南方丘陵区油茶气孔导度的量化研究较少,定量研究气孔导度能够深入了解油茶对外界环境变化的反馈机制与抗旱性能。但直接利用现有的模型会因地区差异而产生较大误差,因此亟需在中国南方丘陵区展开系统深入研究。
目前,量化气孔导度主要有两种方法:一是直接测量法,例如用LI-6400、LCi-SD便携式光合仪等仪器在野外可以直接测量不同植物叶片的气孔导度,能适应于各种不同的环境,但对仪器要求较高,费时费力,有时会因气候因素导致测量不稳定,无法连续精确测量数据。二是模型法,国内外许多学者对气孔导度模型展开了大量的研究,其中最常见的是气孔导度与环境因子的多元非线性经验模型Jarvis模型和与净光合速率及环境因子的线性相关半经验模型Ball-Woodrow-Berry(BWB)模型。Leuning等对BWB模型进行了改进,提出了Ball-Berry-Leuning(BBL)模型。Jarvis模型形式简单,能直接利用环境因子建立多元非线性函数关系,BWB模型和BBL模型参数较少,对样本的需求量较低,具有一定生理意义。除此之外,还有基于脱落酸(Abscisic Acid,ABA)和保卫细胞膨压控制建立的气孔导度模型。模型法避免了直接测量法的缺点,能进行多尺度的气孔导度模拟,在对气孔的环境响应机理认识不足时,模型模拟是较为有效的分析工具。金欣欣等利用改进后的Tardieu-Davies气孔导度模型较好地模拟了水稻气孔导度的日变化过程;彭世彰等引入叶气温差代替了叶水势对气孔导度的影响,改进了Jarvis模型和BBL模型;Medlyn等通过最佳气孔行为理论与光合作用模型相结合,推导出了新的气孔导度模型。虽然前人已经进行了大量的野外试验和模型模拟优化,并建立诸多适用于本地区的气孔导度模型,但由于地理环境和气候影响条件以及作物种类的不同,这些模型参数的适用性存在差别,因此有必要在中国南方丘陵区建立合适的气孔导度模型。
7-8月是中亚热带红壤丘陵区季节性干旱高发期,此时油茶叶片通过气孔蒸腾耗水较大,为研究气孔的开闭情况,因此本研究在油茶果实生长的关键时期以及对土壤水分敏感的时期(6-9月),选择具有代表性、应用最广泛气孔导度模型(Jarvis模型、BWB模型、BBL模型)来模拟并进行修正,探究油茶气孔导度的时间变化和影响因子,进行多模型模拟精度对比分析,建立并筛选出适合本地区油茶树的气孔导度模型,旨在为深入理解南方丘陵区生态系统中油茶表面能量与水分交换提供理论依据,并为节水灌溉提供理论基础。
1 材料与方法
1.1 试验设计和方法
本试验在湖南省国家油茶工程技术研究中心进行(东经28°12',北纬113°06')。占地33.33 hm,海拔100 m左右,为低山丘陵地带,地表土壤以红壤为主,占土壤总面积的75%,田间持水量为20%~31%,属于亚热带季风性湿润气候,年平均气温17.2 ℃,年均日照时长1 500 h左右,年降水量1 361.6 mm,降水丰沛,雨热同期,植被以亚热带常绿阔叶林为主,主要林木为油茶。
试验于2020和2021年6—9月油茶树主要生长季内进行,供试材料为生长3~9 a的长林品种“秋霞”油茶树,树高1.2 m左右,胸径12 cm左右。测试时随机选取长势较好无病害的3棵油茶树,每棵树测定东南西北四个方位的叶片,每月选取晴天与阴天相结合观测2~6 d,采用便携式光合仪(LCi-SD,UK)在7:00—19:00每隔1 h进行监测,不同观测日期再根据天气、日出日落时间等变化情况做出相应调整,以获得气孔导度g(mol/(m·s))、光合速率A(mol/(m·s))、叶片温度T(℃)、胞间CO浓度C(mol/mol)和叶片CO浓度C(mol/mol)等指标。采用微型自动气象观测站(HOBOware,U30 Station,USA)同步记录大气温度T(℃)、相对湿度H(%)和光合有效辐射PAR(mol/(m·s))等,气象站每隔1 min进行采样工作,每隔15 min记录一次数据。利用ECHO (5TE,Decagon Devices Inc.,Pullman,USA)系统每隔10 min连续监测0~60 cm土层土壤含率,并用烘干法进行校正。2020年在试验周期内选择10 d进行日变化监测,获得样本数量81个,土壤含水率变化为12.3%~32.2%;2021年在试验周期内选择12 d进行日变化监测,获得样本数量136个,土壤含水量变化为14.2%~34.6%。模型通过2021年的58组数据根据spss25.0进行非线性回归分析拟合和最小二乘法求解各参数,再用2021年其余的78组数据和2020年的81组数据进行模型验证。
1.2 模型介绍
1.2.1 Jarvis经验阶乘气孔导度模型
目前,针对气孔导度模型的研究,主要以Jarvis建立的经验型阶乘模型和Ball建立的半经验模型的应用比较广泛,Jarvis模型是数个环境因子综合作用的产物,它是在环境因子对气孔导度的影响是相互独立的假设上建立的阶乘模型,模型具体表达式如下:

式中g为气孔导度,(PAR)、(VPD)、(T)、(C)、()分别为光合有效辐射、饱和水汽压差、大气温度、大气中的二氧化碳浓度和叶水势对气孔导度的影响函数,其中f(VPD)、f(T)、f(C)、f()的函数值在0~1之间。通常植物气孔对二氧化碳浓度的响应范围在0~250mol/mol内,而该研究区试验期内二氧化碳浓度平均值在400mol/mol左右,日变化不会超过30%,远大于气孔响应上限,因此二氧化碳浓度对气孔导度的影响可忽略不计,取f(C)=1。
在前人的研究中,对气孔导度的单一因子响应模型给出了不同的表达式。一般气孔导度对光合有效辐射的响应函数采用公式(2)~(4)来计算。
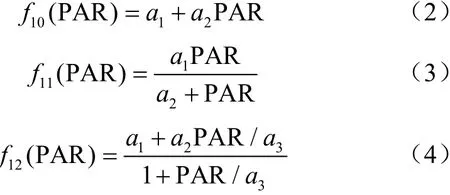
气孔导度对饱和水汽压差的响应函数可用公式(5)~(7)表示。
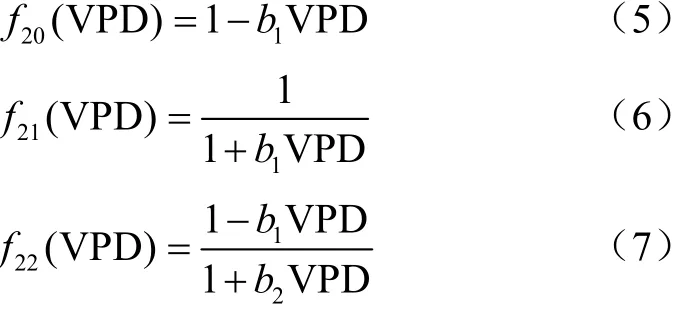
根据Hofstra等的研究,空气温度对叶片气孔导度的响应呈二次曲线关系。
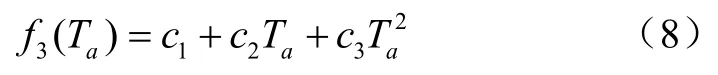
叶片的水分平衡主要取定于土壤水分状况和潜在蒸散,通常用土壤水分影响函数()来代替()。
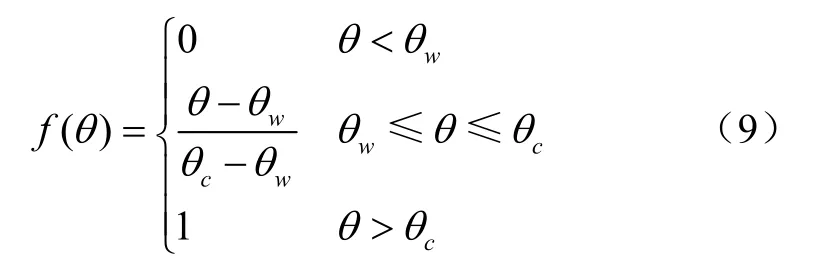
式中、、、、、、、均为待定参数;T为空气温度,℃;为根系吸水层土壤平均含水率,%;本研究选取0~60 cm土层的平均含水率;θ为凋萎含水率;θ为田间最大持水率。利用环刀法对试验区土壤进行试验,测得θ=0.09,θ=0.29,与试验地相同的周寅杰等低山丘陵地区油茶林地土壤水分特性分析中的数据相近。
1.2.2 Leuning-Ball 线性气孔导度模型
Ball等研究发现气孔导度与光合速率具有线性关系,提出了线性气孔导度Ball-Woodrow-Berry(BWB)模型,即
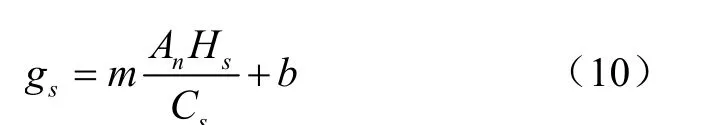
式中A为净光合速率,mol/(m·s);H为大气相对湿度,%;C为叶表面CO浓度,mol/mol;、为经验系数。由于该模型在低CO浓度时受到限制,Leuning等对方程进行了修正,得到了Ball-Berry-Leuning(BBL)模型,即
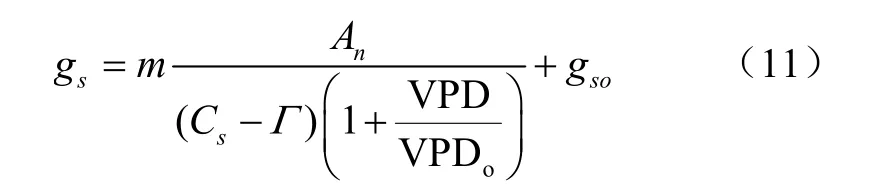
式中为二氧化碳补偿点,mol/mol;VPD为叶片与空气间的饱和水汽压差,kPa;g为光补偿点处的气孔导度值,mol/(m·s);VPD为经验常数。其中饱和水汽压差的计算公式如下:

C3植物的二氧化碳补偿点同叶面温度T(℃)成二次曲线关系,公式如下:
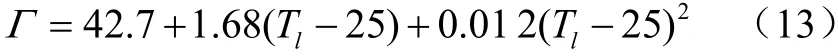
1.3 模型评价
试验观测数据采用spss25.0进行拟合,并给出模型拟合度()和显著性。以均方根误差(Root Mean Square Error,RMSE)、平均绝对误差(Mean Absolute Error,MAE)、模型斜率()评价各模型模拟效果。RMSE、MAE、和的计算公式如下:

式中O表示实测值;M表示模拟值;为样本数。越接近1,表明模型模拟效果越好;RMSE、MAE值越小,说明模拟效果越好;模型斜率反映了模型是否被高低估情况,当>1时,表示模型高估,<1时表示模型低估,接近1,模拟效果越好。
2 结果与分析
2.1 实测气孔导度日变化
在2020、2021年油茶生长最旺盛的6-9月,选取了该时段内典型的晴天和阴天进行分析。从图1可以看出,晴天,油茶气孔导度呈小“双峰”,整体变化趋势基本一致,8:00-11:00,油茶气孔导度随着太阳光合有效辐射和温度的增加而增大,并在11:00达到峰值。12:00-14:00,为防止叶片蒸腾速率过快和油茶体内水分大量散失,植物内部进行自我保护调节,保卫细胞体积缩小,气孔部分关闭,导致气孔导度在中午快速降低。15:00-17:00,随着蒸腾的减小,油茶土壤水分而得到一定程度的补给,导致气孔导度有所回升。阴天,油茶叶片气孔导度因受环境因素不稳定影响,呈现出颠荡起伏的趋势,一天内的波动幅度较晴天小,在10:00-16:00之间变化较为稳定。2021年油茶气孔导度值整体比2020年大,主要原因可能是2020年末油茶树经过了修剪,树冠与树体的光照条件得到改善,光合作用和生理代谢活动增强,促进了叶绿体中与光合作用相关酶的作用;也可能是因为2020年试验期内的平均光合有效辐射(274.6mol/(m·s))、气温(27.5 ℃)、土壤含水率(0.24)等因素与2021年(304.5mol/(m·s))、29.2 ℃、0.27)的差异较大所导致。
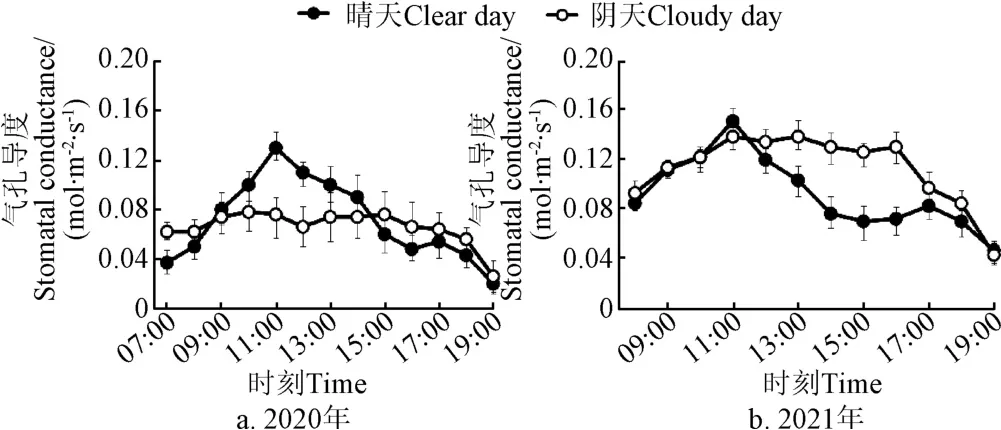
图1 不同年份油茶叶片气孔导度日变化过程Fig.1 Diurnal variation of stomatal conductance of Camellia oleifera leaves in different years
2.2 气孔导度模型确定与比较
将Jarvis模型各因子的表达式((PAR)~(PAR)、(VPD)~(VPD)、(T)、())进行组合,得到了9组Jarvis模型,分别为Jarvis0-~Jarvis8-。有研究引入()后Jarvis模型的精确度反而下降,因此本文把Jarvis0-~Jarvis8-模型去掉()与之比较,探究其是否会提高模型的精度,记为Jarvis-0~Jarvis-8。Yin等曾引入胞间CO浓度(C)来代替叶表面CO浓度,提高了模型的精度。本文探究对比了两种二氧化碳浓度在BBL模型和BWB模型中的适用情况,用叶面CO浓度计算的记为BWB和BBL模型;用胞间CO浓度计算的记为BWB-和BBL-模型。用2021年58组数据运行结果如表1。
分析可得,去掉()后,Jarvis模型模拟效果得到了提高,由于植物水与土壤水存在密切的水力联系,模型对水分响应模拟时,可只考虑其中一种,所以在南方丘陵区利用Jarvis模型时可以忽略这一因子的影响,因此说明Jarvis模型主要受PAR、T以及VPD的影响,土壤水分对Jarvis模型的影响不大。王秋玲等研究得出春玉米在Jarvis模型中加入()后,模型的下降18%,但李丽等引入土壤水分函数Jarvis模型模拟精度得到提高,这可能是低土壤含水率才是影响植物气孔导度的主导因子,而南方夏秋季节多雨,土壤高含水率相比北方较高,所以加入土壤含水量后模型模拟效果反而降低,Zhang等研究也表明在水源充足的情况下,气孔导度模型低估了9%。因此在本试验区可以忽略土壤水分对Jarvis模型的影响。在Jarvis模型中,气孔导度模型Jarvis-3~Jarvis-5整体模拟效果较差,模拟较好的是Jarvis-2和Jarvis-8模型,两者均在=0.01置信度水平上极显著相关,但Jarvis-8的值大于Jarvis-2,绝对值误差比较小,模型斜率也接近于1,模拟出的气孔导度值也比较符合实测值,于是选择Jarvis-8模型来进行油茶叶片气孔导度模拟效果的对比分析,其表达式为
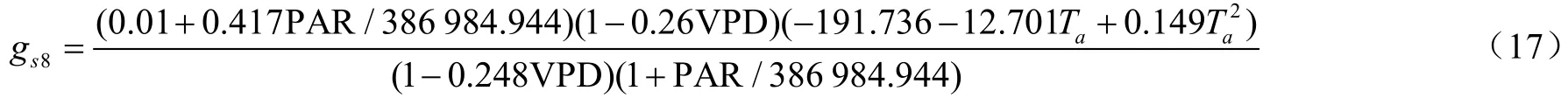
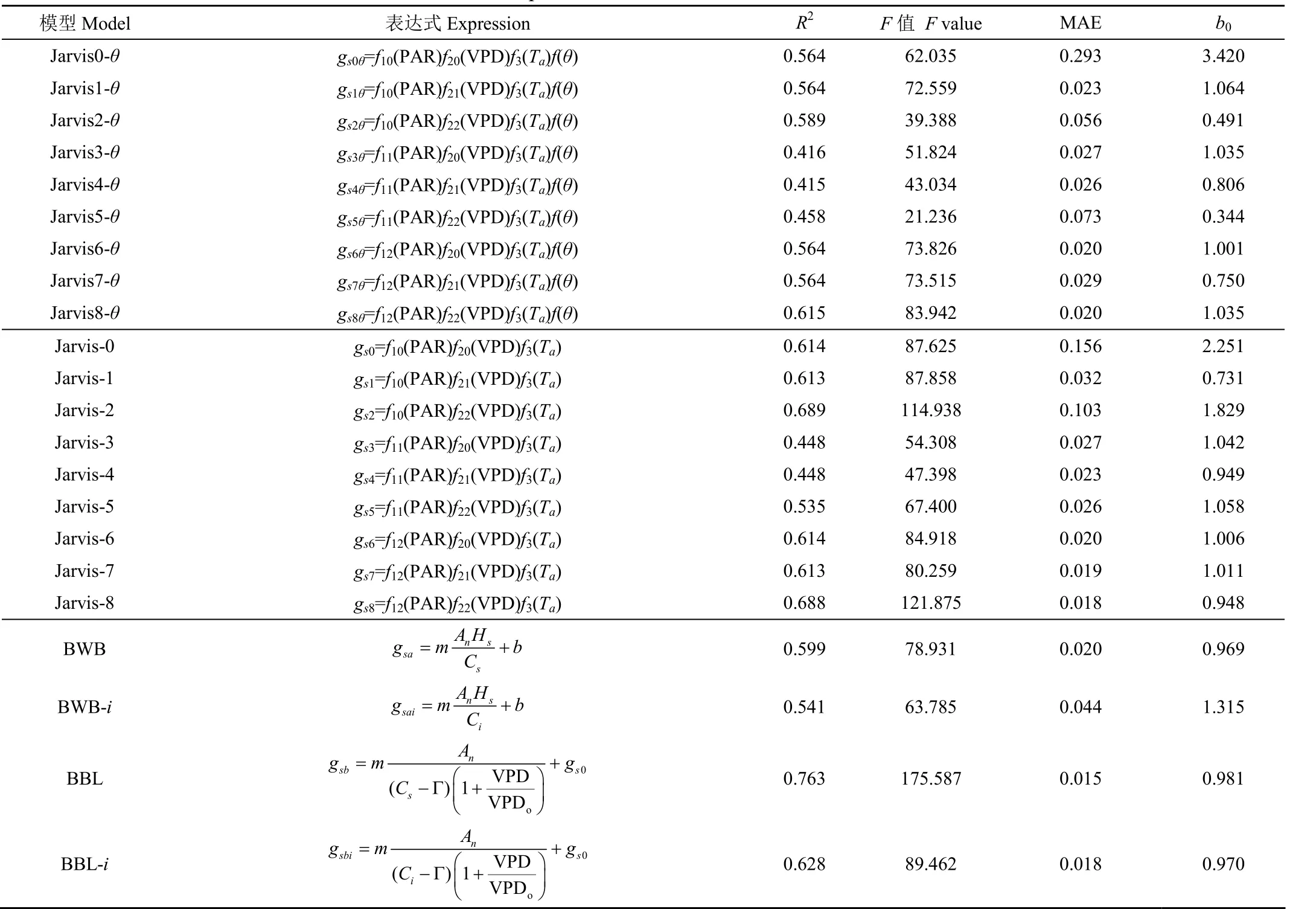
表1 不同气孔导度组合模型比较Table 1 Comparison of different combination models
在BBL和BWB模型中,用叶面CO浓度计算的模型明显优于用胞间CO浓度计算的模型,这与Yin等的结论存在差异,可能是因为其考虑了暗呼吸速率。通过计算,BWB和BBL模型的表达式如下:

将筛选出来的三种模型代入2021年58组数据分别计算气孔导度,结果如图2所示,三种模型模拟结果都比较均匀地分布在实测值的两侧,模拟值与实测值的相关性均通过了0.01水平的显著性检验,表明对油茶气孔导度的预测指示作用较好。整体上,BBL模型(0.76)的优于Jarvis-8模型(0.69)和BWB模型(0.60),平均绝对误差(MAE)BBL模型(0.015)最小,BWB模型最大(0.020),说明BBL模型效果较好,BWB模型效果较差,但三种模型的斜率<1,说明都有所低估。模拟效果从好到差排序依次为:BBL模型、Jarvis-8模型、BWB模型,但在气孔导度实测值较大时,模型模拟值都偏离在1:1线下方,这表明此时段内模型模拟值偏低,从数据分析得出此时段是一天内叶气温差较大、CO内外浓度差较大的时候,气孔导度受环境因子影响,气孔开闭处于不稳定阶段,仪器测定时气孔导度值呈现波动,因此导致这时段内存在一定误差。

图2 油茶气孔导度模拟值与实测值比较Fig.2 Comparison between simulated and measured stomatal conductance of Camellia oleifera
2.3 气孔导度模型修正与模拟
该研究区气孔内外CO浓度差(Δ=C-C)在0~250mol/mol范围内,可能对气孔导度存在一定影响。因Jarvis模型是环境因子的阶乘模型,于是在Jarvis模型中引入气孔内外CO浓度差(Δ),在BBL模型中用Δ来代替C-,探究是否能够增加模型的精确度,记为Jarvis-、BBL-模型。
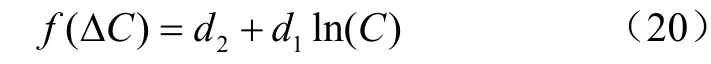
BBL模型其实质仍是BWB模型,因BBL模型拟合度更高,所以选择BBL模型进行修正。引入叶气温差后,分别记为Jarvis8-、和BBL-模型。叶气温差Δ与气孔导度有较强的联系。因此:
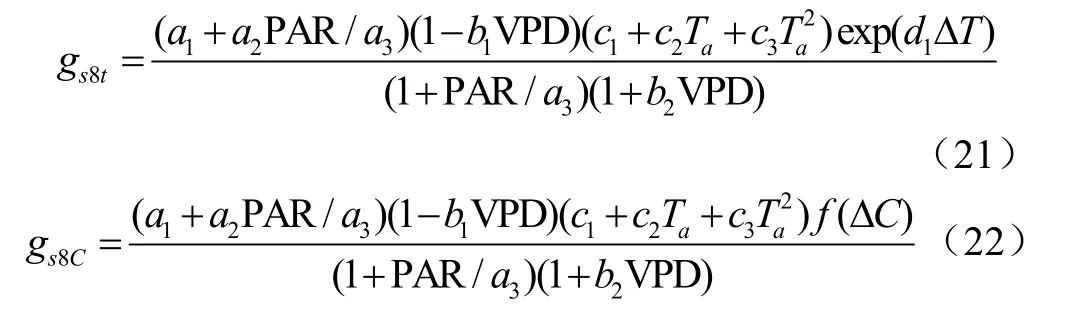
修正后的BBL模型:

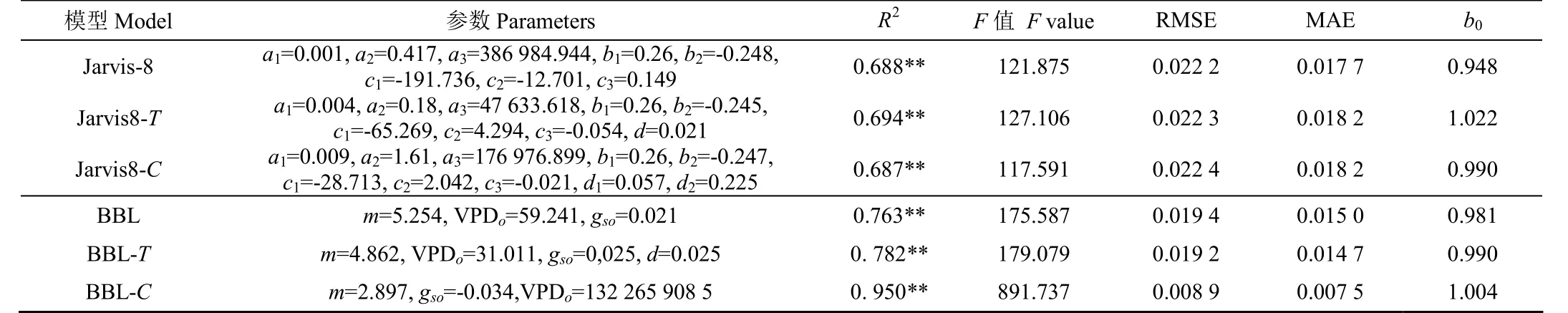
表2 改进的气孔导度模型拟合比较Table 2 Comparison of modified stomatal conductance model fitting
通过2021年58组数据对模型改进对比分析,如表2所示,引入叶气温差对两类模型都有所提高,但效果不显著,R提高非常小,但模型斜率有一定提高。引入二氧化碳浓度差,对Jarvis-8模型影响不大,而对BBL模型影响较大,改进后的BBL-C模型的决定系数从BBL模型的0.76提高到了0.95,提高近10%,模型斜率非常接近于1,使BBL模型不再低估。从整体预测结果上看,Jarvis模型模拟效果从好到差依次为:Jarvis-T、Jarvis-8、Jarvis-,BBL模型模拟效果从好到差依次为:BBL-、BBL-、BBL。
2.4 修正模型比较和验证
选择两类改进模型中最优模型与原模型进行对比,结果如图3a、图3b。可以看出,对于所有改进的Jarvis模型,并没有改变模拟值在实测气孔导度低时偏大、在实测气孔导度高时偏低的情况,因此引入这些因子对Jarvis模型的改进效果不是很显著。而改进的BBL-模型模拟值非常贴近于实测值,提高了模型模拟精度。就实测资料而言,模拟效果从好到差依次为:BBL-、BBL、Jarvis-、Jarvis-8。为更进一步验证BBL-模型的准确性,选取2020年的81组样本数据和2021年没有参加参数拟合的78组样本数据对BBL-模型进行验证,气孔导度模拟值与实测值的对比结果如图3c、图3d所示。模型模拟值和实测值之间拟合度非常高,2020年为0.92,2021年为0.95,2021年气孔导度预测值均匀分布在1:1线周围,偏离程度较小,2020年相对较离散,可能是人为因素和气候条件差异导致。整体上BBL-气孔导度模型对本试验区的气孔导度预测结果准确度较高,可用于模拟本地区的油茶叶片气孔导度值。因此建议改进BBL-气孔导模型作为南方丘陵区油茶叶片气孔导度响应模型。

图3 不同最优改进模型的比较与验证Fig.3 Comparison and verification of different optimal improved models
2.5 不同情况下气孔导度日变化模拟
夏季、秋季油茶叶片因光合速率变化不同,气孔导度呈现不同峰型的变化趋势;阴天油茶叶片气孔导度变化趋势复杂无规律,晴天则出现“M”型变化规律;土壤含水率的变化通过影响保卫细胞及其周围细胞的膨压,进而使气孔开度有不同的变化趋势,为了探究BBL-模型在各种条件下的适用性,选择试验期内油茶不同季节、不同天气、不同土壤含水率日变化3 d平均值进行模拟并与实测值进行比较,结果如图4所示,在不同情况下,模型日变化模拟值与实测值变化趋势趋近一致。晴天比阴天的模拟效果更好,这可能是晴天气孔导度相比与阴天受环境因子的影响较小,比较稳定,阴天模拟值整体上稍微偏高。夏季(5月31日-7月22日)气孔导度模拟值比秋季(7月23日-9月13日)好,夏季模拟值与实测值基本贴近,秋季模拟值则稍微偏大,这可能是因为秋季风速较夏季大,仪器所测气孔导度值受风速的一定影响。在本试验区土壤含水率较低时,日变化模拟值更好,土壤含水率较高时,气孔导度模拟值高于实测值,但在气孔导度最大时模拟值却低于实测值,不过偏离程度非常小。虽然不同情况下日变化模拟值有所差异,但影响很小,几乎可以忽略不计。模拟值与实测值间的日变化规律十分接近,验证了BBL-模型对本地区的适用性。
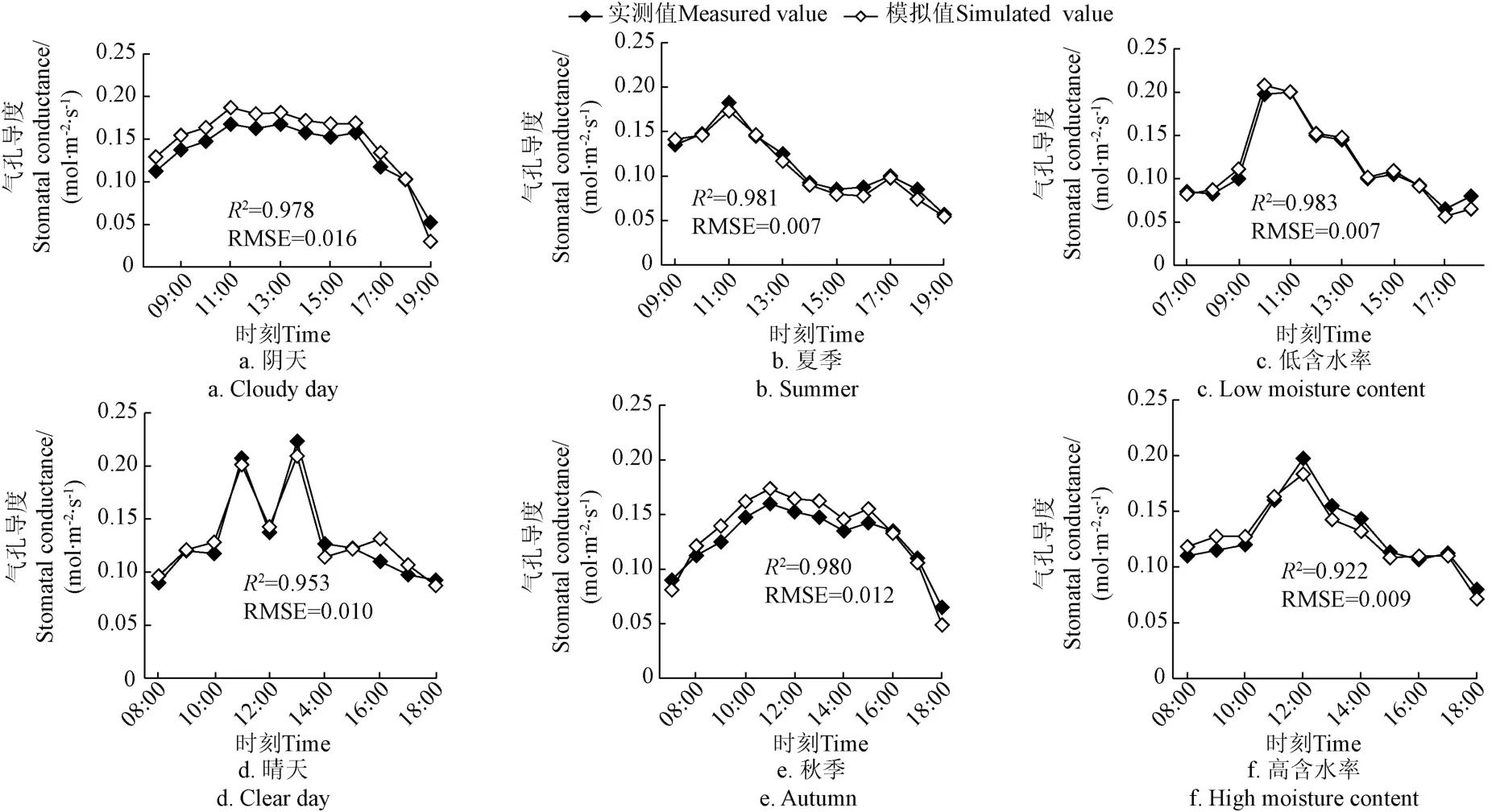
图4 不同场景气孔导度日变化模拟验证比较Fig.4 Simulation and comparison of stomatal conductance diurnal variation in different scenarios
3 讨 论
3.1 气孔导度模型比较
本研究选择应用广泛的Jarvis模型、BBL模型和BWB模型对油茶树气孔导度进行了模拟,研究表明三类气孔导度模型均可以较好地模拟南方丘陵区油茶的气孔导度,其中BBL模型最好,Jarvis-8 模型次之,BWB模型较差。这可能是在南方丘陵区温度对气孔导度影响较大导致,BWB模型具有一定的机理性,耦合了光和作用与气孔导度之间的关系,但没有体现温度对气孔导度影响。BBL模中二氧化碳补偿点拟合了温度对气孔导度的影响(式(13))。Jarvis-8 模型是经验模型,反映了温度因子对气孔导度的响应,但缺乏生理意义。温度通过改变叶片的气孔形态特征以及气孔在叶片表面的规则程度来来影响净光合速率,从而影响叶片气孔导度。如随着温度的增加,植物叶片会产生更多的、更小的维管束鞘,能够有效缩短叶肉细胞内由 PEP 羧激酶固定二氧化碳生成的 C酸从叶肉细胞通过胞间连丝转移到维管束鞘细胞的距离,在一定程度上减小了其所受的叶肉阻力,因此气孔导度增大。其他学者也运用气孔导度模型对植物气孔导度进行了模拟,李泽卿等在青藏高原地区研究高山嵩草叶片气孔导度时,发现BBL 模型比Jarvis 模型能更好地解释叶片气孔导度对环境因子的响应变化,高冠龙等对荒漠河岸柽柳叶气孔导度进行模拟,得出Jarvis 模型比BBL模型的模拟效果更好,鱼腾飞等对柳树叶片气孔导度研究发现BWB模型的模拟效果更好。大量学者对气孔导度模型研究的结论存在着一定的差异,这可能与研究地区的地质环境、气候条件差异相关,且研究方法存在差异,各类植物对影响因子的响应情况也不同,导致叶片气孔行为对环境因子响应复杂、机理不明确,使其在不同的地区得不到统一。
3.2 气孔导度模型修正
本研究引入了叶气温差来改进BBL模型和Jarvis模型,模拟结果显示,引入叶气温差后,两类模型的模拟效果都有所提高,但效果不显著。适宜的叶气温差才能保证植物生理机能的正常运行,叶气温差过高会使得叶片厚度减少,叶片内部的细胞器和组织受到伤害;温差过低会抑制Rubisco 酶的含量与活性,从而抑制气孔的开度。彭世彰等研究表明叶气温差能够较好地反映作物水分和土壤水分从而提高水稻气孔导度模型。本文研究结论与之一致,但效果没有水稻显著,可能是由于水稻和油茶所处环境不同,当叶气温差过大时,叶片可以通过气孔来调控蒸腾,减少水分散失达到降低温差的效果。但水稻比油茶更容易从土壤中吸收水分,以至于模型模拟效果不一致。
本研究引入气孔内外二氧化碳浓度差后,对Jarvis模型的模拟效果影响不大,但显著提高了BBL模型的模拟效果,其改进后的BBL模型决定系数达0.95,值达891.737,决定系数提高了10%。BBL-C模型模拟结果以及对不同情况下的日变化模拟与实测值都极为贴近。二氧化碳浓度对气孔导度的影响尚无定论,但有研究指明二氧化碳浓度可以改变叶片气孔个数、单个气孔形态特征和气孔空间分布格局,从而影响叶片气体交换过程。植物体细胞间二氧化碳浓度会随大气二氧化碳浓度增加而增加,为维持细胞间二氧化碳分压低于大气二氧化碳,植物通过调节气孔开度或减少气孔数量来达到细胞间二氧化碳浓度适宜的目的,因此气孔内外二氧化碳浓度差对气孔的开闭起着重要的调节作用。引入气孔内外二氧化碳浓度差后出现的结果可能是由气孔导度模型的机理所导致的,Jarvis模型是经验模型,其主要受净光合辐射、饱和水汽压差、温度的影响,气孔内外浓度差对其影响并不显著所致。BBL模型则反映了环境二氧化碳浓度、净光合速率、相对湿度对气孔导度的影响,根据费克定律得知,二氧化碳进入叶片时会受到气孔阻抗的影响,气孔导度是气孔阻抗的倒数,与气孔内外二氧化碳浓度差成反比关系。因此在BBL模型中用气孔内外二氧化碳浓度差(0~250mol/mol)代替C-的模拟效果更好,这也说明低二氧化碳浓度可能对气孔导度的影响较大,范金杰等也发现小麦在高二氧化碳浓度条件下叶片气孔导度的增大幅度逐渐缩小。本文直接在BBL模型上引入气孔内外浓度差进行修正,与叶子飘等所构建的模型结论一致,这也证实了BBL-模型对小麦和油茶都是非常适用的。
本研究为南方丘陵区油茶叶片建立了适合的气孔导度模型,为量化油茶气孔导度以及南方丘陵区其他植物的气孔导度提供了参考,存在的不足在于BBL-模型虽然能很好地模拟本地区的气孔导度,参数也比较少,但模型需要获取光合速率值,可能会影响气孔导度模型的精度。BBL-模型能否适用于大范围以至于所有作物还有待研究,因所试验研究对象是C3植物,C4植物还没有涉及,试验样本也较少。因此探究光合速率模型与气BBL-模型之间的耦合问题把BBL-模型推广到更大范围尺度以及推到冠层尺度是未来研究的重点工作。
4 结 论
本文主要对南方丘陵区油茶叶片气孔导度g进行了模拟,比较了Jarvis 模型、BBL模型和 BWB 模型的适用情况,对Jarvis模型和BBL模型进行了多种方法的优化与修正,主要的研究结论如下:
1)南方丘陵油茶树气孔导度晴天日变化呈双峰趋势,上午峰值出现在11:00左右;随后由于温度继续升高,蒸腾强烈导致气孔关闭,气孔导度减小;并于15:00-17:00 回升出现第二次峰值。阴天波动幅度较晴天小。
2)Jarvis模型中,去掉土壤水分函数后,模型的模拟效果得到了提高,说明在南方丘陵区土壤水分对气孔导度的影响不大,因此可忽略其对Jarvis模型的影响。从气孔导度对单一因子不同响应模型中得出Jarvis-8的模拟效果最优。BBL模型和BWB模型中,用叶片二氧化碳浓度的拟合效果优于用胞间二氧化碳浓度的拟合效果。对Jarvis模型、BWB模型、BBL模型进行验证比较,结果表明BBL模型的模拟效果最优,其次是Jarvis模型,BWB气孔导度模型效果最差。
3)对Jarvis模型和BBL模型进行修正验证比较发现,叶气温差对两类模型的修正效果有所提升但不显著,气孔内外二氧化浓度差对Jarvis模型影响不大,但对BBL模型影响显著,改进后的BBL-模型的模拟值与实测值的决定系数从BBL模型的0.76提高到了0.95,并很好地模拟了2020和2021的气孔导度值,分别为0.92和0.95,同时在不同情景下,BBL-模型的模拟值与实测值具有十分接近的日变化规律。整体上模拟效果从好到差依次为:BBL-模型、BBL-模型、BBL模型、Jarvis-模型、Jarvis模型、Jarvis-模型。推荐BBL-气孔导度模型作为南方丘陵区油茶叶片气孔导度响应模型。
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
干旱地区农业研究(2022年1期)2022-01-28 04:51:30
环境保护与循环经济(2021年7期)2021-11-02 08:10:54
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
哈尔滨轴承(2020年1期)2020-11-03 09:16:22
节水灌溉(2020年9期)2020-09-16 00:32:12
装备制造技术(2019年12期)2019-12-25 03:06:52
中国奶牛(2019年10期)2019-10-28 06:23:36
科技创新与品牌(2019年12期)2019-02-06 04:03:29
