
7种北美橡树的光合生理特性分析
2018-03-17 01:52吴晓龙曹基武彭继庆江良为
中南林业科技大学学报 2018年3期
吴晓龙,吴 毅,张 斌,曹基武,彭继庆,李 琪,江良为,李 杰
(中南林业科技大学 林学院,湖南 长沙 410004)
光合作用是植物的基本生命活动之一,是植物吸收太阳能量、同化二氧化碳和水,制造有机物质、释放氧气的过程;是太阳辐射能进入生态系统并转化为化学能的主要形式,也是制约生态系统生物生产力的最重要的生理过程[1-4]。植物的光合作用是一个复杂的生理过程,受植物本身内部和外界环境的双重影响,光合特性的测定也要从植物内部影响因素如气孔导度、胞间CO2浓度、蒸腾速率等和外界环境因子如光照强度、空气温度、湿度等两方面展开。作者利用Li-6400P光合作用测定仪对7种北美橡树的光合特性进行测定,测定内容包括不同北美橡树的光合响应曲线与光合日变化。通过比较分析,确定7种北美橡树的光合规律以及不同树种之间的光合差异,旨在为探讨其生长发育的一般规律、科学利用这一种质资源提供基础数据。
1 材料与方法
1.1 试验材料
实验在长沙中南林业科技大学苗圃进行,10月初,选择3年生、胸径5~7 cm、树高1.5~2.0 m的北美橡树引种幼林进行光合作用的测定。7种北美橡树分别是柳栎Quercus phellos、娜塔栎Quercus nuttallii、沼生栎Quercus michauxii、猩红栎Quercus coccinea、舒玛栎Quercus shumardii、南方红栎Quercus falcate、无柄栎Quercus imbricaria。实验区长沙属亚热带季风性湿润气候,气候温和,降水充沛,雨热同期,四季分明,年平均气温17.2 ℃,年积温为5 457 ℃,市区年均降水量1 361.6 mm。
1.2 光响应曲线的测定
在10月初晴朗的天气,利用Li-6400便携式光合仪在自然空气CO2浓度下(380~400 μmol·mol-1),由红蓝光源控制光照辐射范围在0 ~ 2 000 μmol·m-2s-1,将光强梯度设置为:0、50、100、200、400、600、800、1 000、1 200、1 400、1 600、1 800、2 000 μmol·m-2s-1共13个梯度,间隔时间2~3 min,测量光合生态因子的变化。测定时叶片温度为25 ℃左右,空气相对湿度为(60.00%±1.00%)。每个树种分别选择5株健壮幼树,并选取长势、生长部位、叶龄基本一致的成熟叶片挂牌标记,多次测定数据取均值。
以光合有效辐射(PAR)为横轴,净光合速率(Pn)为纵轴绘制光响应曲线,依据非直角双曲线方程进行线性拟合,计算光合参数[5]。拟合方程为:

式中:Pn为净光合速率,Pmax为最大净光合速率,K为曲角,为暗呼吸速率,PAR为光合有效辐射,α为表观量子效率。在拟合曲线中,Pn=0时的Q为光补偿点(LCP),Pn=Pmax时的Q为光饱和点(LSP)。
1.3 光合日变化测定
采用LI-6400便携式光合仪测定净光合速率(Pn)蒸腾速率(Tr)、胞间CO2浓度(Ci)、叶片气孔导度(Gs)等光合参数的日变化,测定时段为8:00—18:00。测定时利用标准叶室,空气流量设置为 500 μmol·s-1,每隔 2 h 测定 1 次,仪器自动记录光合有效辐射(PAR)、空气温度(Ta)和湿度(RH)等环境因子,测定4组数据取均值。
1.4 数据分析
借助Photosyn Assistant计算LCP和LSP;采用GraphPad Prism软件,结合编程功能,将拟合方程作为自定义公式,进行光响应曲线拟合;采用SPSS 17.0和Excel 2010软件进行数据统计分析和绘图。
2 结果与分析
2.1 不同北美橡树的光响应曲线及特征参数分析
根据7种北美橡树光响应曲线(见图1),将7种北美橡树分为光合能力强和光合能力弱两组。光合能力强组有其娜塔栎、柳栎、舒玛栎、沼生栎4种北美橡树,光合能力弱组有猩红栎、无柄栎、南方红栎3种北美橡树。7种北美橡树的净光合速率日变化(见图4)也证明这一结论。
2.1.1 不同北美橡树的光响应曲线分析
如图1所示,光合能力强组的光响应曲线增长趋势基本一致,在PAR为0~160 μmol·m-2s-1时,4种北美橡树的Pn值与PAR有极好的线性关系,呈直线上升,表明此时光照强度是控制北美橡树光合作用的主要因子;在PAR为160~550 μmol·m-2s-1时,Pn仍随 PAR 增强而升高,但上升的速率趋缓,而舒玛栎和沼生栎已经逐渐达到光饱和点。当PAR>550 μmol·m-2s-1时,4种北美橡树的净光合速率达到光饱和点,并且保持在Pmax上下波动。
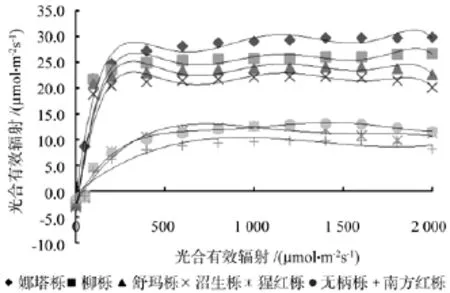
图1 不同北美橡树的光响应曲线Fig.1 Different north American oaks of light response curves
光合能力弱组的猩红栎、无柄栎、南方红栎3种北美橡树光响应曲线增长趋势也基本一致,在PAR为0~250 μmol·m-2s-1时,3种北美橡树的Pn值与PAR呈直线上升,此时光照强度是控制3种北美橡树光合作用的主要因子;在PAR为250~700 μmol·m-2s-1,Pn仍随 PAR 增强而升高,但上升的速率趋缓,并逐渐达到光饱和点。当PAR>700 μmol·m-2s-1时,Pn达到光饱和点,而猩红栎因强光的抑制,Pn有下降趋势。
2.1.2 不同北美橡树光响应曲线特征参数分析
如表1所示,娜塔栎的Pmax最大为31.3 μmol·m-2s-1,而在 7 种北美橡树中,Pmax最小的南 方 红 栎 为 11 μmol·m-2s-1, 娜 塔 栎 的Pmax是 南方红栎Pmax的2.8倍,差异性显著(显著性水平0.012),7种北美橡树的Pmax具体表现为:娜塔栎>柳栎>舒玛栎>沼生栎>猩红栎>无柄栎>南方红栎,α大小表现为娜塔栎>舒玛栎>柳栎>沼生栎>猩红栎>无柄栎>南方红栎。7种橡树的光补偿点LCP表现为娜塔栎和沼生栎最低 (8 μmol·m-2s-1),猩红栎最高 (40 μmol·m-2s-1)。光饱和点 LSP 表现为舒玛栎最低 (312 μmol·m-2s-1),无柄栎最高(708 μmol·m-2s-1),综合以上提到的光响应曲线特征参数,发现娜塔栎、柳栎和舒玛栎具有较高的光合潜力,不仅夜间对同化物的氧化分解速率低,而且白天对光能的利用效率还高,能在光合作用中积累更多的生物量,提高植株的生长速度[6-7]。
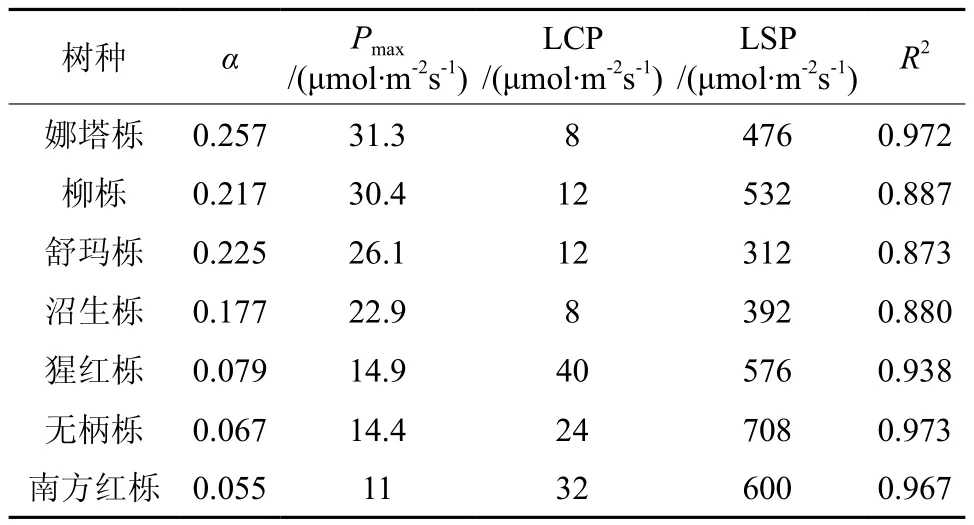
表1 不同北美橡树光响应曲线特征参数Table 1 Different light response curve characteristic parameters of north America oak
LSP和LCP是植物利用光强能力的重要指标,LSP高的植物被认为能更有效地利用强光,LCP低的植物被认为能更有效地利用弱光[8-9],由所求出的光补偿点可以看出光合能力强组的4种北美橡树有较低 LCP(LCP 在 10 μmol·m-2s-1左右)和 LSP(LSP 在 310 ~ 540 μmol·m-2s-1之间),差异性显著(显著性水平0.011),表明这4种北美橡树有很较强的弱光利用能力和耐阴性。而光合能力弱组的猩红栎、无柄栎和南方红栎的LCP和LSP都要比光合能力强组的高,但LCP和LSP都在 24 ~ 40 μmol·m-2s-1、576 ~ 708 μmol·m-2s-1之间,表明光合能力弱组的3种北美橡树也具有耐阴性。
2.2 主要环境因子与不同北美橡树的光合参数日变化
2.2.1 光合有效辐射、相对湿度与空气温度日变化
光合有效辐射(PAR)、相对湿度(RH)和空气温度(Ta)日变化规律对应图2和图3。在一天之中,光合有效辐射与空气温度的变化趋势大致同步。在8:00—14:00,光合有效辐射和空气温度都随着时间的增加而不断上升,下午14:00,光合有效辐射达到一天中的最大值2 000 μmol·m-2s-1,一天中的空气温度同样对应达到最大值26℃;在下午14:00—18:00光合有效辐射和空气温度都随时间的变化开始急剧下降,光合有效辐射下降到一天中的最低值 600 μmol·m-2s-1,相比中午下降 70%,下降速度较快。光合有效辐射与空气温度出现同步现象,主要是因为长沙秋季辐射中的紫外还是较强,短波光产生的能量较大,能迅速提升空气温度所致,因此光照强弱会直接影响空气温度的高低。
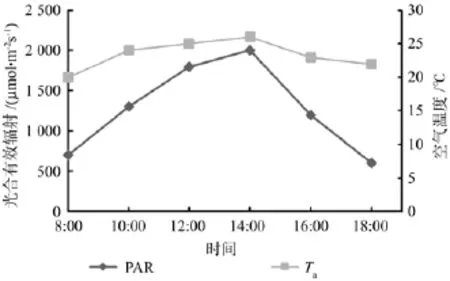
图2 光合有效辐射和空气温度日变化Fig.2 Photosynthetic active radiation and air temperature daily variation
如图2所示,相对湿度的日变化表现出总体下降的趋势,但在下午12:00—14:00相对湿度有短暂的回升。对比图2、图3可以看出,8:00—12:00随着辐射增强与空气温度升高,相对湿度则逐渐降低;在下午12:00—14:00,由于阵雨的出现,相对湿度并未出现持续降低,而是略有升高;下午14:00—18:00时,由于长沙的下午空气温度依然很高,空气积温,导致相对湿度持续降低,下午18:00达到一天中的最低值(53.00%)。从相对湿度的日变化还可以看出,长沙地区秋季湿度较大,常日保持在60.00%左右。
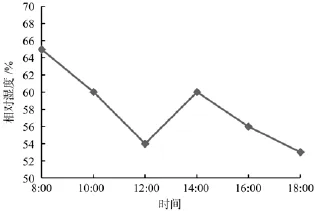
图3 相对湿度日变化Fig.3 Relative humidity daily variation
2.2.2 不同北美橡树的光合日变化
从图4可以看出,大多数北美橡树的叶片净光合速率(Pn)日变化呈单峰型,不存在明显“午休”现象,峰值在12:00—14:00时间段,但猩红栎和无柄栎的叶片净光合速率呈双峰型变化,第一个峰值出现在10:00,第二个峰值出现在14:00,10:00—14:00可能是猩红栎和无柄栎逐渐进入光合“午休”阶段。Pn日变化总体表现为柳栎>娜塔栎>沼生栎>舒玛栎>无柄栎>南方红栎>猩红栎。
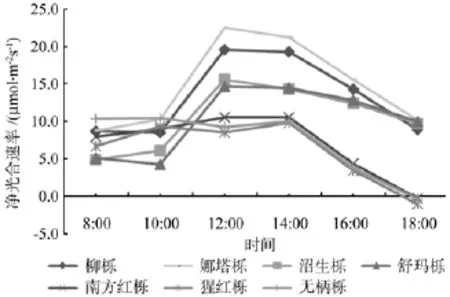
图4 不同北美橡树的净光合速率日变化Fig.4 The daily variation of net photosynthetic rate of different north American oak
如图5所示,不同北美橡树的气孔导度(Gs)日变化大多呈上午高下午低的趋势,并且上午和下午的差距很大,在下午18:00时7种北美橡树的气孔导度值下降到一天中的最低点,但是,沼生栎和娜塔栎在14:00—16:00和12:00—14:00依然保持较高的气孔导度值,此时这2种北美橡树可能依然保持较快的光合作用物质交换。不同北美橡树的胞间CO2浓度(Ci)大多呈早晚高中午低的“V”形变化(见图6),并在下午14:00达到最低值,但柳栎和娜塔栎一直呈下降趋势,这应该和柳栎、娜塔栎全天一直具有较多的物质转换和较高的光合速率有关。7种北美橡树的蒸腾速率日变化(Tr)整体呈早高晚低下降的趋势,但是部分树种呈早低中高晚低的单峰型变化(见图7),峰值在10:00—12:00,并且都在下午18:00下降到最小值。由此可见,一天中7种北美橡树的气孔导度(Gs)和蒸腾速率(Tr)变化呈相近趋势,和胞间CO2浓度(Ci)变化大致呈相反趋势。
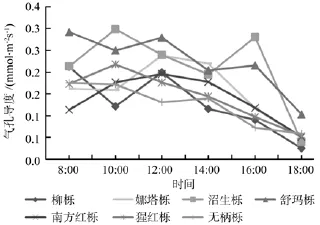
图5 不同北美橡树的气孔导度日变化Fig.5 The daily variation of stomatal conductance of different north American oak
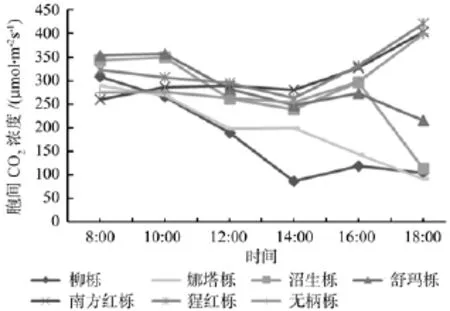
图6 不同北美橡树的胞间CO2浓度日变化Fig.6 The daily variation of intercellular CO2 concentration of different north American oak
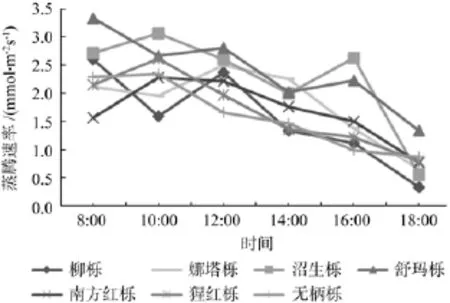
图7 不同北美橡树的蒸腾速率日变化Fig.7 The daily variation of transpiration rate of different north American oak
2.3 不同北美橡树净光合速率的影响因子分析
影响植物光合速率的因素既来自外部,如光照强度、空气温度和相对湿度等,也来自于内部,如气孔导度、胞间CO2浓度、蒸腾速率、生长年龄等[10-13]。如表2,通过相关系数分析表明:①柳栎叶片Pn的相关性PAR>Ta>Ci>RH>Gs>Tr,在所有内外因素中只有PAR与Pn呈显著正相关性,这表明柳栎的Pn随着PAR的增加不断加快。影响因素中柳栎的Gs和Tr、PAR和Ta呈显著正相关,表明柳栎的蒸腾速率随着气孔的张开而不断加快。因为PAR、Ta为外部因素,所以7种北美橡树的PAR与Ta均呈显著正相关,下面不一一赘述。②沼生栎叶片Pn的相关性Ta>PAR>RH>Ci>Gs>Tr,但Pn与所有影响因素相关性均不显著。影响因素中Gs、Ci、Tr3个影响因子互为显著正相关。③舒玛栎Pn的相关性Ci>Ta>PAR>RH>Tr>Gs,但Pn与所有内外因素都不显著,影响因素中Tr分别于Gs、Ci呈显著正相关。④南方红栎Pn的相关性Gs>Ci>Tr>PAR>Ta>RH,在所有内外因素中Pn与Gs、Tr呈显著正相关,Pn与Ci呈显著负相关,表明数值较高的气孔导度与强烈的蒸腾作用,会导致南方红栎叶片胞间二氧化碳浓度降低,产生较高的净光合速率。影响因素中Gs与Tr、PAR呈显著正相关。⑤猩红栎Pn的相关性Ci>Gs>PAR>Tr>Ta>RH,在所有内外因素中Pn与Gs呈显著正相关,Pn与Ci呈负相关。影响因素中Gs与Tr呈显著正相关,Ci与PAR呈显著负相关。⑥无柄栎Pn的相关性Gs>Ci>Tr>RH>PAR>Ta,在所有内外因素中Pn与Gs、Tr呈显著正相关,Pn与Ci呈负相关。影响因素中Gs与Tr呈显著正相关。⑦娜塔栎Pn的相关性PAR>Ta>Gs>Tr>RH>Ci,在所有内外因素中Pn与PAR、Ta呈显著正相关。影响因素中Gs与PAR、Tr呈显著正相关,Ci与RH呈显著正相关。通过内外因素的相关性分析,可以看出柳栎和娜塔栎的光合影响因素具有一定相似性,两者Pn均与PAR呈显著正相关;南方红栎、猩红栎和无柄栎的光合影响因素同样具有相似性,三者的Pn与Gs都呈显著正相关,与Ci都呈显著负相关;另外舒玛栎和沼生栎的Tr与Gs、Ci都具有显著正相关,并且内外影响因素相关性系数相近,表明舒玛栎和沼生栎的光合作用具有相似性。
3 结论与讨论
(1)不同北美橡树的光响应曲线和净光合速率日变化表明:可以将7种北美橡树分为有娜塔栎、柳栎、舒玛栎、沼生栎的光合能力强组和有猩红栎、无柄栎、南方红栎光合能力弱组。表现量子效率(α)可以反映绿色植物对光的利用能力,α值越大,表明绿色植物利用光的能力越强[14]。在7种北美橡树中,光合能力强组的α值要远大于光合能力弱组,其中娜塔栎的α值最高,光利用能力最强,反映该树种较其他树种在湖南地区有最好的光利用能力与适应性。植物叶片的光补偿点(LCP)与光饱和点(LSP)反映出植物对光的利用条件,一般情况下,拥有较低LCP和LSP被认为耐隐性植物,反之则属于阳生植物[15]。参照姜琳[16]对栎树光合特性的研究,7种北美橡树耐弱光,但不耐强光,属于耐隐性植物。植物叶片的最大净光合速率(Pmax)反映植物的光合潜力。娜塔栎与柳栎的Pmax在7种北美橡树中最大,可见娜塔栎与柳栎的光合潜力最强。在园林绿地造景时,根据不同橡树的光合生理特性,选择适宜的栽培地点,更好发挥树种的生态效益。
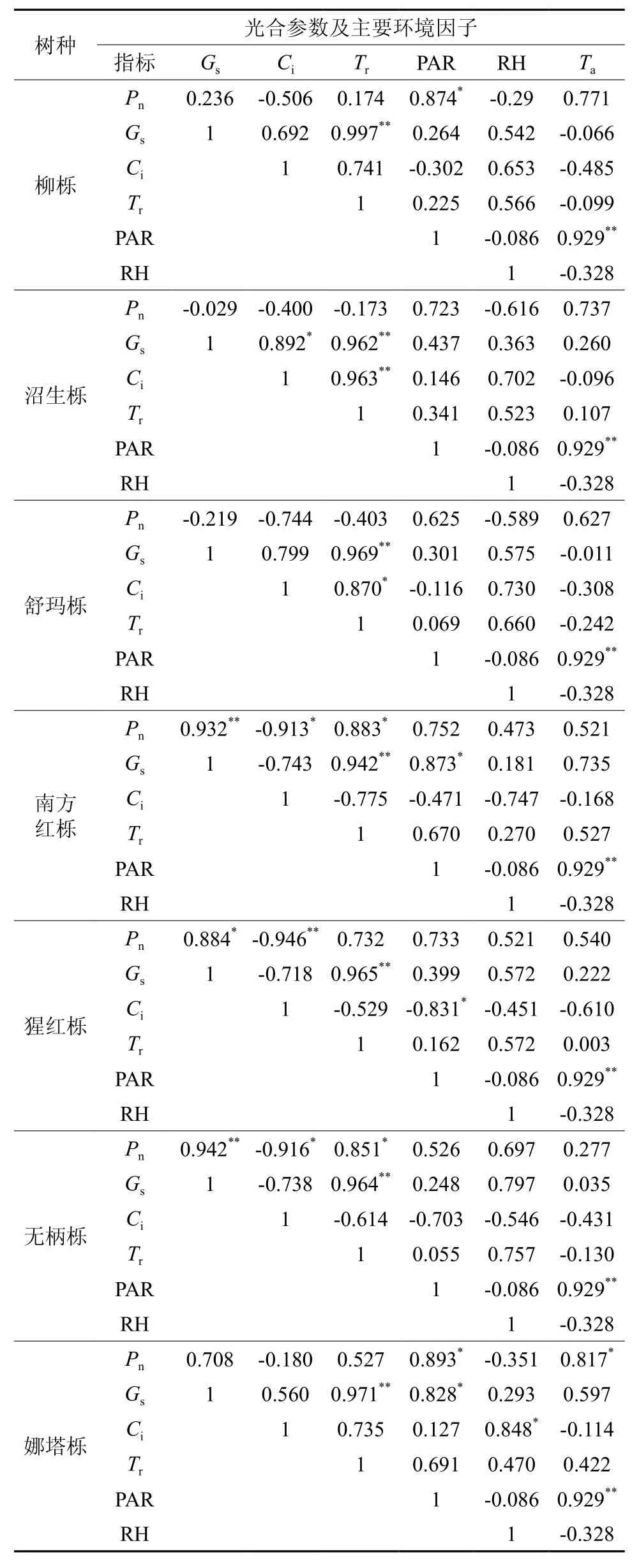
表2 不同北美橡树光合参数及其主要环境因子相关系数†Table 2 The correlation coefficient of different north American oak photosynthetic parameters and the main environmental factor
(2)通过对7种北美橡树的光合日变化进行分析,发现绝大多数北美橡树的净光合速率(Pn)日变化呈单峰型,不存在明显光合“午休”现象,这与姜琳[16]对3种栎树秋季光合日变化的研究结果基本一致。不同北美橡树的气孔导度(Gs)日变化呈上午高、下午低的趋势,这或许与植物在上午进行较快的光合作用有关,上午较快的光合速率加快了植物光合作用物质的交换。对于北美橡树的胞间CO2浓度(Ci)和蒸腾速率(Tr)日变化,大多数北美橡树的Ci呈早晚高、中午低的“V”形变化,Tr呈早高、晚低下降趋势。通过对比7种北美橡树的Gs、Ci、Tr日变化,发现气孔导度(Gs)的变化趋势大致与蒸腾速率(Tr)相近,与胞间CO2浓度(Ci)相反。这或许与一天中光照强度的变化有关,光照的增强和植物蒸腾速率的增加,使得植物细胞气孔开张,加快了细胞间的物质交换,导致细胞间CO2浓度下降。
(3)光合指标的大小受到诸多因素的影响如生态因子、生理因子和生化因子。在生产活动中,可以通过调节植物光合影响因素来提高植物光合指标,增强植物光合作用[17-20]。相关性分析显示,柳栎叶片Pn的相关性PAR>Ta>Ci>RH>Gs>Tr,娜塔栎叶片Pn的相关性PAR>Ta>Gs>Tr>RH>Ci,可以发现柳栎、娜塔栎的光合影响因素具有一定的相似性,两者的Pn均与PAR呈显著正相关,相关系数分别为0.874和0.893。增加光照条件可以加快柳栎、娜塔栎的光合速率,促进柳栎、娜塔栎幼苗的生长。
作为在我国近几年引种的园林绿化树种,目前针对北美橡树的系统性研究还很少,且对于北美橡树光合特性的研究在国内更是旱见。本研究综合地分析了以上7种北美橡树在秋季10月份的光合特性,比较7种北美橡树光合作用的差异性,为以后该树种在我国的培育与栽植提供光合生理方面的理论指导。同时,在此基础上对以上7种北美橡树做更加系统的研究,期待能更加全面地指导北美橡树规范化栽培。
[1]杨 超,鲁绍伟,陈 波,等.北京地区常见果树的光合速率和固碳释氧效应[J].经济林研究,2016,34(1):57-64.
[2]权 红,李连强,袁 芳,等.7个颠茄品系的光合特性比较研究[J].中药材,2016,39(7):1457-1461.
[3]吴家兵,关德新,张 弥,等.长白山地区蒙古栎光合特性[J].中国科学院研究生院学报,2006,23(4):549-554.
[4]丁瑞霞,聂俊峰,杨宝平.便携式光合作用测定仪测定作物指标问题探讨[J].现代农业科技,2015(24):147-150.
[5]吴泽龙,谭晓风,袁 军,等.油茶不同叶龄叶片形态与光合参数的测定[J].经济林研究,2016,34(2):24-29.
[6]曹军胜,刘广全.刺槐光合特性的研究[J].西北农业学报,2005, 14(3):118-122,136.
[7]朱万泽,王金锡,薛建辉.台湾桤木引种的光合生理特性研究[J].西北植物学报2004,24(11):2012-2019.
[8]薛 雪,李娟娟,郑云峰,等.5个常绿园林树种夏季光合蒸腾特性[J].林业科学,2015,51(9):150-155.
[9]全先奎,王传宽.兴安落叶松对环境变化的物候驯化和光合能力适应[J].生态学报2015,35(13):4538-4546.
[10]阚祥绪,施利民,周恭伟.植物光合作用的测定及不同波长的光对光合作用的影响[J].生物学通报,2005,40(4):50-51.
[11]许大全.光合作用测定及研究中一些值得注意的问题[J].植物生理学通讯,2006,42(6):1163-1167.
[12]余 刚,邢 昊,张永君.东北红豆杉光合特性研究[J].吉林林业科技,2015,44(5):19-30.
[13]胡启鹏,孙玲玲,郭志华,等.常绿阔叶林中几种不同功能组树种叶形态、光合及光诱导对不同环境的响应[J].华南农业大学学报,2013,34(2):207-212.
[14]王泽瑞.几种红色系彩叶树种光合特性及呈色机理的研究[D].合肥:安徽农业大学,2001.
[15]高 鹤,宗俊勤,陈静波,等.7种优良观赏草光合生理日变化及光响应特征研究[J].草业学报,2010,19(4):87-93.
[16]姜 琳,杨 暖,姜官恒,等.三种不同叶色栎树光合特性研究[J].山东林业科技,2016(2):12-16.
[17]李林锋.4种湿地植物光合作用特性的比较研究[J].西北植学报,2008,28(10):2094-2102.
[18]唐建宁,康建宏,许 强,等.秦艽与小秦艽光合日变化的研究[J].西北植物学报,2006,26(4):836-841.
[19]高 超,闫文德,田大伦,等.杜仲光合速率日变化及其与环境因子的关系[J].中南林业科技大学学报,2011,31(5):100-104.
[20]蒋 燚,李志辉,朱积余,等.不同红锥种源幼林光合特性日变化测定与分析[J].中南林业科技大学学报,2013,33(6):43-47.
猜你喜欢
疯狂英语·新读写(2022年1期)2022-01-28
小哥白尼(神奇星球)(2021年12期)2021-03-08
小哥白尼(野生动物)(2020年10期)2021-01-07
农业与技术(2020年12期)2020-07-04
农学学报(2019年1期)2019-02-21
新课程·下旬(2017年7期)2017-08-14
中国诗歌(2016年6期)2016-11-25
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
