
匙吻鲟稚鱼摄食节律与饥饿对其体脂肪酸组成的影响
2017-12-18 11:07聂文强田晶晶周继术
水产科学 2017年1期
聂文强,李 婧,吉 红,3,田晶晶,周继术,王 涛
( 1. 西北农林科技大学 动物科技学院,陕西 杨凌 712100; 2.安康市瀛湖生态旅游区渔业局,陕西 安康 725000; 3. 西北农林科技大学 安康水产试验示范站,陕西 安康 725000 )
匙吻鲟稚鱼摄食节律与饥饿对其体脂肪酸组成的影响
聂文强1,李 婧2,吉 红1,3,田晶晶1,周继术1,王 涛1
( 1. 西北农林科技大学 动物科技学院,陕西 杨凌 712100; 2.安康市瀛湖生态旅游区渔业局,陕西 安康 725000; 3. 西北农林科技大学 安康水产试验示范站,陕西 安康 725000 )
在水温20~27 ℃下,将体质量2~3 g的匙吻鲟稚鱼饲养在55 cm×45 cm×35 cm的塑料箱(约86 L)内,投喂天然浮游动物,采用肠充塞度和饱满指数法,研究其摄食节律和饥饿72 h后全鱼脂肪酸组成的变化。结果显示,匙吻鲟稚鱼摄食的最高峰在17:00,次高峰在5:00,摄食低谷在23:00,胃内食物6 h排空,而肠在36 h仍未排空;经过0~72 h饥饿后,匙吻鲟全鱼饱和脂肪酸和单不饱和脂肪酸含量随饥饿时间的延长而显著下降(P<0.05),而多不饱和脂肪酸水平和二十碳五烯酸比例显著上升(P<0.05),二十二碳六烯酸比例升高,但差异不显著(P>0.05)。结果表明,匙吻鲟稚鱼为晨昏摄食型;饥饿期间主要消耗饱和脂肪酸和单不饱和脂肪酸供能,而多保留多不饱和脂肪酸,尤其是选择性保留二十碳五烯酸和二十二碳六烯酸。
匙吻鲟;摄食节律;排空时间;饥饿
鱼类的摄食量随时间变化而呈一定的规律(即摄食节律)。这是鱼类对其所在环境条件主动适应的结果。鱼类早期摄食行为生态学已成为水产学科研究的重要领域。了解鱼类的摄食节律可以指导渔业生产,例如按照鱼类摄食节律投喂,可增加饲料的转化率,减少饲料的浪费。胃肠排空时间是影响鱼类食欲的主要因子之一,胃肠饱满度及其排空速率决定了鱼类摄入食物的量[1-3]。在确定鱼类摄食节律和胃肠排空时间的前提下,对其停食可以进一步研究饥饿对鱼类的影响。近年来,鱼类饥饿状态下生理及生化变化以及饥饿后补偿生长效应已经成为鱼类营养研究引人注目的领域[4]。饥饿期间,鱼体内贮存的能量物质随饥饿时间的延长而不断消耗,体质量也随饥饿时间的延长而逐渐下降,体内水分比例却逐渐上升[5-7]。然而,匙吻鲟(Polyodonspathula)的摄食节律、胃肠排空时间以及饥饿状态下体成分变化尚未见报道。
匙吻鲟又称匙吻白鲟、鸭嘴鲟,属鲟形目、匙吻鲟科,是北美洲密西西比河流域特有的滤食性淡水经济鱼类,适应性强、生长迅速。目前有关匙吻鲟养殖的研究主要集中于生物学、养殖及繁殖技术、消化酶的研究及肉质评价等[8-15]方面。因此,深入研究匙吻鲟摄食特性,以及饥饿对匙吻鲟稚鱼脂肪酸组成变化的影响,揭示匙吻鲟在饥饿状态下的脂肪酸组成变化和饥饿时对脂肪酸的利用顺序,对匙吻鲟养殖具有一定的指导意义。本试验研究了匙吻鲟摄食节律和胃肠排空时间,探讨了在饥饿状态下全鱼脂肪酸组成的变化和饥饿时匙吻鲟的主要供能脂肪酸以及脂肪酸比例变化规律,为匙吻鲟养殖及营养生理生化研究提供参考。
1 材料与方法
1.1 匙吻鲟稚鱼昼夜摄食节律试验
1.1.1 试验鱼的饲养
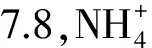
1.1.2 摄食节律评估
试验期间,每隔3 h采样1次,即8:00、11:00、14:00、17:00、20:00、23:00、2:00、5:00连续采样8次。每次随机取鱼10尾。逐尾测量全长、体质量,测定温度和溶解氧及充塞度和肠饱满指数。胃、肠充塞度采用0~5六个等级(0级:空肠管或者肠管内有极少量食物;1级:只有部分肠管中有少量食物即食物占肠管的1/4;2级:全部肠管中有少量食物或食物占肠管的1/2;3级:食物较多,充塞度中等,食物占肠管的3/4;4级:食物多,充塞全部肠管;5级:食物极多,肠管膨胀)。
饱满指数是在对消化道内的食物种类鉴定、计数后,按下式计算:
饱满指数=m1/m2×10000
式中,m1为食物质量(mg),m2为鱼体质量(mg)。
逐尾计算被测鱼的饱满指数后,再求各次的平均饱满指数,以此作为各次摄食饱满指数。
1.1.3 肠胃排空时间
从西北农林科技大学安康水产试验示范站苗种培育池随机取出60尾鱼(来源、规格同上),放在55 cm×45 cm×35 cm盛有清水的平底塑料箱内中饥饿,每3 h取5尾鱼,逐尾测量全长、体质量,测定温度和溶解氧,解剖测定充塞度和饱满指数。观察匙吻鲟稚鱼36 h内肠和胃的排空情况。
1.2 饥饿对匙吻鲟稚鱼脂肪酸组成的影响试验
在摄食旺盛时,取饱食鱼80尾,放入上述规格的塑料箱内。根据上述胃排空时间的结果确定饥饿时间的起点。刚停食时作为0点,6、12、24、48、72 h时各取6尾全鱼,放入-20 ℃保存,以测定脂肪酸。
脂肪酸测定方法:称取0.3~0.5 g粉碎后的全鱼于10 mL离心管中,加入甲醇∶氯仿(1∶2)5 mL,高速分散器(XHF-D,SCIENIZ®,宁波)匀浆,静置1~2 h后定量滤纸过滤,加4 mL蒸馏水,3000 r/min离心5 min,去上清液,下层用水浴锅负压抽干(40 ℃)。随后加入1 mL色谱纯正己烷,将油脂溶解,加1 mL、0.4 mol/L KOH-甲醇溶液静置30 min进行甲酯化,之后加2 mL去离子水,待分层后提取上层溶液在气相色谱仪(安捷伦7820a,安捷伦科技,美国)上进行测定。不同脂肪酸含量按面积归一化法计算,以总脂肪酸的百分比表示。
在此基础上,参照Rombenso等[16]的方法,以0 h为对照计算距离系数(Djh值)。其计算公式如下:
式中,h代表不同的饥饿时间,即为6、12、24、48、72 h。先计算不同饥饿时间每种脂肪酸与0 h之差,再把所有脂肪酸差值平方加起来再开方。
1.3 数据处理
脂肪酸数据用平均值±标准差表示,平均值用SPSS 18.0软件中的Duncan进行多重检验,P<0.05为差异性显著,P>0.05定为无显著性差异。
2 结 果
2.1 匙吻鲟稚鱼昼夜摄食节律
一昼夜中,17:00匙吻鲟稚鱼的饱满指数最高,摄食最旺盛(表1)。24 h内的饱满指数表明,匙吻鲟稚鱼有2个摄食高峰,一个摄食低谷。摄食的最高峰在17:00,其饱满指数为777.06,次高峰在5:00,其饱满指数为640.71,摄食低谷在23:00,其饱满指数为441.76。匙吻鲟稚鱼的摄食为明显的晨昏摄食型。
2.2 匙吻鲟稚鱼肠胃排空时间
匙吻鲟稚鱼的胃在2:30彻底排空,即经过了6 h胃彻底排空。在36 h内匙吻鲟稚鱼的肠充塞度变化较小,在36 h后仍未排空(图1)。

表1 匙吻鲟稚鱼摄食节律的测定结果
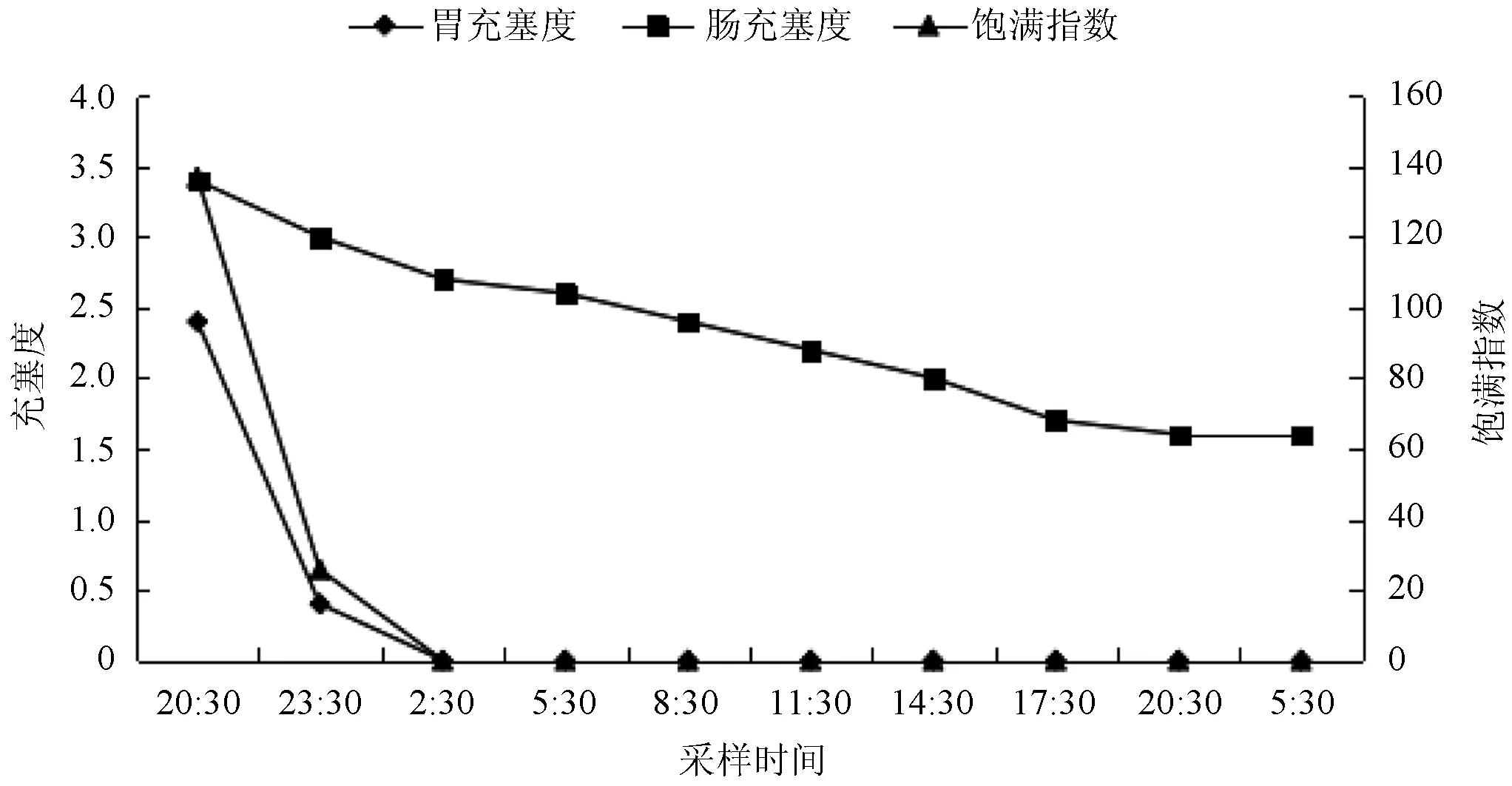
图1 匙吻鲟稚鱼胃、肠充塞度及饱满指数变化
2.3 饥饿后匙吻鲟稚鱼脂肪酸的组成
在饥饿不同时间的匙吻鲟稚鱼中共检测出14种脂肪酸,碳链长度在14碳至22碳之间,其中饱和脂肪酸3种,单不饱和脂肪酸4种,多不饱和脂肪酸7种(表2)。随饥饿时间的延长,饱和脂肪酸含量显著下降(P<0.05),从0 h的(40.29±0.46)%降至72 h的(33.03±1.95)%,但并非所有的饱和脂肪酸都随着饥饿时间的延长而下降,其中C14:0呈先升后降、再上升的趋势。单不饱和脂肪酸也随着饥饿时间的延长而显著下降(P<0.05),从0 h的(34.83±1.76)%降至72 h的(24.04±2.20)%,但C20:1 n-9却先升后降再升。多不饱和脂肪酸呈上升趋势,由0 h的(24.88±1.67)%升至72 h的(42.92±1.09)%。但并非所有的多不饱和脂肪酸都呈上升趋势,C18:2 n-6呈先降后升再降而后升的波浪线趋势,C20:4 n-6呈先升后降再升的波浪线趋势,C18:3 n-3呈先降后升再降的趋势;而二十碳五烯酸和二十二碳六烯酸则一直升高,这与多不饱和脂肪酸的趋势一致。饱和脂肪酸、单不饱和脂肪酸和多不饱和脂肪酸的变化表明,饥饿期间匙吻鲟稚鱼主要消耗饱和脂肪酸和单不饱和脂肪酸供能,而多不饱和脂肪酸多予以保留,较少消耗供能;而二十碳五烯酸和二十二碳六烯酸呈上升趋势,表明匙吻鲟稚鱼对二十碳五烯酸和二十二碳六烯酸有较强的选择性保留能力。
由表2可知,Σ n-6 多不饱和脂肪酸呈先升后降趋势,而Σ n-3 多不饱和脂肪酸则一直呈上升趋势。距离系数显示,随着饥饿时间的延长,距离系数持续升高。

表2 饥饿对匙吻鲟稚鱼脂肪酸组成变化的影响 %
注:标相同字母的平均值间无显著性差异(P>0.05),标不同字母的平均值间有显著性差异(P<0.05),n=3.
3 讨 论
3.1 匙吻鲟稚鱼的摄食节律
鱼类的摄食节律可分为白天摄食、晚上摄食、晨昏摄食和无明显节律4种类型。不同种类摄食节律不同;同种鱼类不同生长阶段摄食节律也可能不同[17]。邹桂伟等[18]研究表明,大口鲇(Silurusmeridionalis)仔鱼在晴朗或少云的天气下,昼夜摄食节律明显,即白天摄食很少,夜间摄食强度增大,具有两个摄食高峰期(清晨和傍晚)。本研究结果表明,匙吻鲟稚鱼多在晨昏摄食,属于晨昏摄食型。在17:00时匙吻鲟的摄食强度达到最高峰,饱满指数为647.71;5:00还有另一个次摄食高峰;23:00为摄食低谷。这一研究结果与大口鲇仔鱼的研究结果一致。朱爱民等[19]研究表明,匙吻鲟仔鱼全天只在16:00—18:00有一个摄食高峰。而本研究中匙吻鲟稚鱼摄食最高峰发生在17:00,也与16:00-18:00所得结果相吻合,但除此之外还发现了一个次高峰(5:00),这一现象很可能与两次试验所用匙吻鲟的规格不同有关。

3.2 饥饿对匙吻鲟稚鱼体脂肪酸组成的影响
不同鱼类耐受饥饿的能力不同,即使同一种类在不同阶段其耐受饥饿的能力也不同。一般认为,仔、稚鱼耐受饥饿的能力弱,饥饿是鱼类早期死亡率高的主要原因之一。仔、稚鱼体内贮存的能量物质少,饥饿期间主要消耗体内蛋白质来提供维持生命活动所需要的能量,导致仔、稚鱼体质量急剧下降,体形异常,体内呈现不可恢复的损害[24-26],直至死亡。而大规格鱼种和成鱼体内贮存的能量物质多,在饥饿状态下,主要消耗贮存的能量物质来提供维持其生命活动所需要的能量,因此大规格鱼种和成鱼在饥饿状态下,体质量下降缓慢,体形变化不大,机体损伤较轻[27-28]。匙吻鲟终生摄食浮游生物,仔、稚鱼阶段摄食量大,一旦饵料不足就会饥饿。由于匙吻鲟仔、稚鱼耐受饥饿的能力弱,此阶段会大量死亡,因此在匙吻鲟仔稚鱼培育阶段一定要保证饵料充足。
饥饿状态下,不同鱼类对脂肪中脂肪酸的利用情况不同,如非洲鲇(Clariasgariepinus)在饥饿状态下体内饱和脂肪酸下降,单不饱和脂肪酸和多不饱和脂肪酸上升[26],而虹鳟(Oncorhynchusmykiss)[29]和锦鲤(Cyprinuscarpio)[30]在饥饿状态下体内饱和脂肪酸和单不饱和脂肪酸含量比例下降,多不饱和脂肪酸上升。饥饿状态下,金头鲷(Sparusaurata)[31]和尼罗罗非鱼(Oreochromisnilotica)[32]对脂肪酸的利用顺序为:∑ n-6 多不饱和脂肪酸> ∑ n-9 多不饱和脂肪酸>∑ n-3 多不饱和脂肪酸。这说明脂肪酸在饥饿状态下的损失速度因双键位置的不同而异。柳敏海[33]等研究表明,在饥饿状态下,鱼(Miichthysmiiuy)对脂肪酸的利用顺序为:∑ n-9多不饱和脂肪酸>∑ n-6多不饱和脂肪酸> ∑ n-3多不饱和脂肪酸。施兆鸿[34]等发现,随着饥饿时间的延长,点带石斑鱼(Epinephelusmalabaricus)幼鱼饱和脂肪酸和单不饱和脂肪酸含量下降,多不饱和脂肪酸含量上升,∑ n-9多不饱和脂肪酸下降,∑ n-3多不饱和脂肪酸和∑ n-6多不饱和脂肪酸在饥饿期间一直上升,但∑ n-6多不饱和脂肪酸比∑ n-3多不饱和脂肪酸上升幅度小,这说明饥饿状态下点带石斑鱼幼鱼对脂肪酸的利用顺序为:∑n-9多不饱和脂肪酸> ∑ n-6多不饱和脂肪酸>∑n-3多不饱和脂肪酸。然而,马珊[35]等指出,大鳍(Mystusmacropterus)在饥饿期间,∑ n-3多不饱和脂肪酸总体呈先降后升的趋势,∑n-6多不饱和脂肪酸总体呈上升趋势,∑ n-9多不饱和脂肪酸总体呈下降趋势,说明在饥饿期间大鳍对脂肪酸的利用顺序为:∑n-9多不饱和脂肪酸>∑n-3多不饱和脂肪酸>∑n-6多不饱和脂肪酸。本研究发现,在饥饿状态下,匙吻鲟稚鱼对体内脂肪酸的利用顺序有一定规律,即首先利用饱和脂肪酸,其次利用单不饱和脂肪酸,最后才利用多不饱和脂肪酸,顺序为:∑n-6多不饱和脂肪酸>∑n-3多不饱和脂肪酸,与金头鲷[31]、尼罗罗非鱼[32]、鱼[33]及点带石斑鱼[34]相似,而却与大鳍[35]有区别,这可能是不同物种间的差异。
距离系数是不同饥饿时间与0 h时脂肪酸的距离系数。距离系数越小表明该时间点的脂肪酸组成与第0 h的相似程度越大,距离系数越大表明与第0 h的相似程度越小。本研究发现随着饥饿时间延长,距离系数越来越大,表明随着饥饿时间延长,与0 h的相似程度越来越小,暗示脂肪酸组成在饥饿影响下呈现规律性的变化。随着饥饿时间的延长,匙吻鲟稚鱼首先利用饱和脂肪酸,然后利用不饱和脂肪酸,最后才动用高不饱和脂肪酸,这可能是造成距离系数一直升高的原因。
4 结 论
匙吻鲟稚鱼的摄食节律为晨昏摄食型,停食后6 h胃排空,而肠在36 h内仍未排空;随着饥饿时间的延长,匙吻鲟稚鱼趋向于主要消耗饱和脂肪酸和单不饱和脂肪酸供能,多不饱和脂肪酸多予以保留,而对二十碳五烯酸和二十二碳六烯酸有较强的选择性保留能力。
[1] Bromley P J. The effects of food type, meal size and body weight on digestion and gastric evacuation in turbot,ScophthalmusmaximusL[J]. Journal of Fish Biology, 1987, 30(4):501-512.
[2] Storebakken T, Kvien I S, Shearer K D, et al. Estimation of gastrointestinal evacuation rate in Atlantic salmon (Salmosalar) using inert markers and collection of faeces by sieving: evacuation of diets with fish meal, soybean meal or bacterial meal[J]. Aquaculture, 1999, 172 (3):291-299.
[3] Riche M, Haley D I,Oetker M, et al. Effect of feeding frequency on gastric evacuation and the return of appetite in tilapiaOreochromisniloticus(L.) [J]. Aquaculture, 2004, 234(1):657-673.
[4] 陈斌, 冯健, 吴彬, 等. 饥饿对太平洋鲑鱼体脂肪与脂肪酸的影响[J]. 海洋与湖沼, 2012, 43(6):1247-1253.
[5] Bilton H T, Robins G L. The effects of starvation and subsequent feeding on survival and growth of Fulton channel sockeye salmon fry (Oncorhynchusnerka)[J]. Journal of the Fisheries Board of Canada, 1973, 30(1):1-5.
[6] Weatherley A H, Gill H S. Recovery growth following periods of restricted rations and starvation in rainbow troutSalmogairdneriRichardson[J]. Journal of Fish Biology, 1981, 18(2):195-208.
[7] Quinton J C, Blake R W. The effect of feed cycling and ration level on the compensatory growth response in rainbow trout,Oncorhynchusmykiss[J]. Journal of Fish Biology, 1990, 37(1):33-41.
[8] 杨明生, 熊邦喜, 黄孝湘. 匙吻鲟人工繁殖F2的早期发育[J]. 华中农业大学学报, 2005, 24(4):391-393.
[9] 王凡. 匙吻鲟的生物学特性及养殖技术[J]. 湖北农业科学, 2007,46(6):985-986.
[10] 董宏伟, 韩志忠, 康志平, 等. 匙吻鲟含肉率及肌肉营养成分分析[J]. 淡水渔业, 2007,37(4):49-51.
[11] 殷守仁, 赵文, 刘保占. 匙吻鲟的生物学特性、成鱼养殖技术及消化系统的解剖[J]. 北京农业, 2009(33):52-56.
[12] 吉红, 单世涛, 曹福余, 等. 安康地区利用家鱼产卵池培育匙吻鲟苗种技术研究[J]. 中国水产,2009(3):43-44.
[13] 吉红, 孙海涛, 单世涛. 池塘与网箱养殖匙吻鲟肌肉营养成分及品质评价[J]. 水产学报, 2011, 35(2):261-267.
[14] Ji H, Sun H T, Xiong D M. Studies on activity,distribution and zymogram of protease,α-amylase and lipase in the paddlefishPolyodonspathula[J]. Fish Physiology and Biochemistry, 2012, 38(3):603-613.
[15] 王丽宏, 吉红, 胡家, 等. 匙吻鲟、杂交鲟和鳙肌肉品质的比较研究[J]. 食品科学, 2014, 35(1):62-68.
[16] Rombenso A N, Trushenski J T, Jirsa D, et al. Successful fish oil sparing in white seabass feeds using saturated fatty acid-rich soybean oil and 22:6n-3 (DHA) supplementation[J]. Aquaculture, 2015(448):176-185.
[17] Helfman G S. Fish behaviour by day, night and twilight [G]//Pitcher T J. The behaviour of teleost fishes, New York:Springer US, 1986:366-387.
[18] 邹桂伟, 潘光碧, 胡德高, 等. 大口鲇仔鱼摄食行为的初步观察[J].水利渔业, 1994(6):15-17.
[19] 朱爱民,梁银铨,黄道明,等.匙吻鲟仔稚鱼摄食特性研究[J].华中农业大学学报, 2008, 27(6):755-758.
[20] Booth M A, Tucker B J, Allan G L, et al. Effect of feeding regime and fish size on weight gain, feed intake and gastric evacuation in juvenile Australian snapperPagrusauratus[J]. Aquaculture, 2008, 282(1):104-110.
[21] Adamidou S, Nengas I, Alexis M, et al. Apparent nutrient digestibility and gastrointestinal evacuation time in European seabass (Dicentrarchuslabrax) fed diets containing different levels of legumes[J]. Aquaculture, 2009, 289(1):106-112.
[23] 董桂芳,杨严鸥,陈路,等. 斑点叉尾和杂交鲟幼鱼昼夜摄食节律和胃肠排空时间的研究 [J]. 水生生物学报, 2013, 37(5):876-884.
[24] Miglavs I, Jobling M. Effects of feeding regime on food consumption, growth rates and tissue nucleic acids in juvenile Arctic charr,Salvelinusalpinus, with particular respect to compensatory growth[J]. Journal of Fish Biology, 1989, 34(6):947-957.
[25] Mehner T, Wieser W. Energetics and metabolic correlates of starvation in juvenile perch (Percafluviatilis)[J]. Journal of Fish Biology, 1994, 45(2):325-333.
[26] Zamal H, Ollevier F. Effect of feeding and lack of food on the growth, gross biochemical and fatty acid composition of juvenile catfish [J]. Journal of Fish Biology, 1995, 46(3):404-414.
[27] Dobson S H, Holmes R M. Compensatory growth in the rainbow trout,SalmogairdneriRichardson [J]. Journal of Fish Biology, 1984, 25(6):649-656.
[28] Kim M K, Lovell R T. Effect of restricted feeding regimes on compensatory weight gain and body tissue changes in channel catfishIctaluruspunctatusin ponds[J]. Aquaculture, 1995, 135(4):285-293.
[29] Kiessling A, Johansson L, Storebakken T. Effects of reduced feed ration levels on fat content and fatty acid composition in white and red muscle from rainbow trout[J]. Aquaculture, 1989, 79(1):169-175.
[30] Murata H, Higashi T. Selective utilization of fatty acid as energy source in carp [J]. Bulletin of the Japanese Society of Scientific Fisheries, 1980, 46(11):1333-1338.
[31] Koven W M, Kissil G W, Tandler A. Lipid and n-3 requirement ofSparusauratalarvae during starvation and feeding[J]. Aquaculture, 1989, 79(1/4):185-191.
[32] Satoch S, Takeuchi T, Watanabe T. Effect of starvation and environmental temperature on proximate and fatty acid composition ofTilapianilotica[J]. Bull Jpn Soc Sci Fish, 1984, 50(1):79-84.
[33] 柳敏海,罗海忠,傅荣兵,等.短期饥饿胁迫对鱼生化组成、脂肪酸和氨基酸组成的影响[J]. 水生生物学报, 2009, 33(2):230-235.
[34] 陈波,柳敏海,施兆鸿,等.饥饿和再投饲对点带石斑鱼幼鱼脂肪酸和氨基酸组成的影响[J].上海水产大学学报, 2008, 17(6):674-679.

DailyFeedingRhythmandEffectsofStarvationonFattyAcidCompositioninJuvenilePaddlefish(Polyodonspathula)
NIE Wenqiang1, LI Jing2, JI Hong1,3, TIAN Jingjing1, ZHOU Jishu1, WANG Tao1
( 1. College of Animal Science and Technology, Northwest Agricultural & Forestry University,Yangling 712100, China; 2. Fisheries Bureau of Yinghu Lake Ecological Tourist Area in Ankang, Ankang 725000, China;
3. Ankang Fisheries Research Station of Northwest Agricultural & Forestry University, Ankang 725021, China )
The feeding rhythm and gastrointestinal evacuation were studied in juvenile paddlefish (Polyodonspathula) with body weight of 2—3 g which was reared in 55 cm×45 cm× 35cm tanks and fed wild zooplankton at water temperature of 20—27 ℃ by fullness degree and intestine fullness methods. The results showed that the feeding peak was observed at 17:00, followed by at 5:00, and the minimal feeding at 23:00, and stomach evacuation was found in 6 hours, without intestinal evacuation in thirty-six hours starvation. There were significant decrease in contents of saturated fatty acids(SFA) and monounsaturated fatty acids(MUFA)in juvenile paddlefish fasted for 0—72 hours(P<0.05)with the elapse of starvation. However, there were significant increase in the content of polyunsaturated fatty acids(PUFA)(P<0.05) with the elapse of starvation, especially eicosapentaenoic acid(EPA) ratio(P<0.05), without significant difference in docosahexaenoic acid(DHA) ratio (P>0.05). The findings indicated that paddlefish juveniles had twilight feeding rhythm, and mainly consumed SFA and MUFA to supply energy during starving period, while PUFA including EPA and DHA was retention rather than utilization for energy supply during starving period.
paddlefish; feeding rhythm; evacuation time; starvation
10.16378/j.cnki.1003-1111.2017.01.006
S965.199
A
1003-1111(2017)01-0036-06
2016-02-26;
2016-04-18.
陕西省科技统筹创新工程计划项目(2015KTTSNY01-05);安康市科技计划项目(2013AK02-11).
聂文强(1990—) ,男,硕士研究生;研究方向:水产动物营养与饲料学.E-mail:171138058@qq.com.通讯作者:吉红(1967—),男,教授;研究方向:水生动物营养与饲料学.E-mail:jihong@nwsuaf.edu.cn.
猜你喜欢
大电机技术(2022年4期)2022-08-30
水产科学(2022年1期)2022-01-26
渔业科学进展(2021年6期)2022-01-04
浙江海洋大学学报(自然科学版)(2021年1期)2021-07-06
南方水产科学(2021年3期)2021-06-23
临床超声医学杂志(2020年4期)2020-04-30
水产科学(2018年2期)2018-03-27
水产科学(2017年3期)2017-12-18
海洋世界(2017年1期)2017-02-13
中国粮油学报(2016年5期)2016-01-23
