
基于广义加性模型的天津近海鱼卵、仔稚鱼群落结构研究
2022-01-26 08:43:00张树林张达娟姚冬梅
水产科学 2022年1期
张 迎,张树林,张达娟,姚冬梅
( 天津农学院 水产学院,天津市水产生态及养殖重点实验室,天津 300384 )

目前,对鱼卵、仔稚鱼的研究多集中在江河口和海湾,如长江口及邻近水域的鱼卵、仔稚鱼群落结构的研究[9-11],黄河口水域及邻近水域的鱼卵、仔稚鱼种类和数量分布的研究[12-13],以及浙江省杭州湾[14]、福建省三沙湾与诏安湾[15-16]、江苏省连云港的海州湾[17]等地均有关于鱼卵、仔稚鱼群落结构的研究。此外,还有针对不同海区的鱼卵、仔稚鱼的研究,如对黄海[18]、东海[5]和南海[19]的鱼卵、仔稚鱼群落结构特征的研究。关于渤海的相关研究主要集中在莱州湾[8,20-22]和辽东湾[23],而天津近岸海域鱼卵、仔稚鱼的系统性研究可追溯至20世纪80年代,张闰生等[1]较为系统地描述了天津近岸海域的鱼卵、仔稚鱼的种类组成、时空分布、经济鱼类的产卵时间。20世纪80年代后对鱼卵和仔稚鱼的研究出现了一定断层,至2018年为止,1984、1989—1991、1993—2005年以及2009—2011年间对鱼卵、仔稚鱼的相关研究未见报道[24-28];对天津海域鱼卵、仔稚鱼的研究多集中于鱼卵、仔稚鱼密度、丰度等方面,而针对天津近岸海域鱼卵和仔稚鱼的群落结构特征及其与环境因子关系的研究罕见报道,也缺乏全年的监测结果。笔者旨在通过2018、2019年4—11月天津近岸海域鱼卵、仔稚鱼及环境调查数据,分析天津近岸海域鱼卵、仔稚鱼及环境系统的变化特征,以期为天津近岸海域渔业资源的可持续开发利用提供科学资料,并为合理利用海洋资源提供参考依据。
1 材料与方法
1.1 站点布设
分别于2018、2019年4—11月在天津近海开展逐月调查,共16个航次,调查区域为N 38°30′~38°50′,E 117°40′~118°00′,设9个调查站位(图1)。

图1 2018、2019年天津近岸海域调查站位Fig.1 Survey stations in the coastal area of Tianjin in 2018 and 2019
1.2 采样方法
按照GB/T 12763.6—2007《海洋调查规范》采样。调查采用110.3 kW渔船,鱼卵、仔稚鱼采集用大型浮游生物网(网口内径80 cm,全长280 cm,网衣孔径0.505 mm,网口面积0.5 m2,校正值0.3),航速2 kn下,在海水表层进行水平拖网10 min,网口系有网口流量计。采集到的样品用体积分数5%甲醛溶液固定,然后带回实验室对鱼卵、仔稚鱼标本进行分拣统计,参考《中国近海鱼卵与仔鱼》[29]进行种类鉴定。采用pH meter、ORP instrument、flexi溶解氧测定仪、Portable Refractometer现场测定pH、氧化还原电位、水温、溶解氧、盐度等环境因子。
1.3 数据分析
1.3.1 鱼卵、仔稚鱼丰度
鱼卵、仔稚鱼丰度按下式计算:
式中,Ga为单位体积海水中鱼卵或仔稚鱼个体数(个/m3);Na为全网鱼卵或仔稚鱼个体数(个);S为网口面积(m2);L为流量计转数;c为流量计校正值。
1.3.2 优势种
鱼卵、仔稚鱼的优势度(I)利用相对重要性指数简化公式计算:
I=(N/%)×(F/%)
式中,N为每航次调查过程中某种鱼卵、仔稚鱼个体数量占鱼卵、仔稚鱼总量的百分比(%);F的百分比为每航次调查过程中某种鱼卵、仔稚鱼出现的频率百分比(%)。
对鱼卵、仔稚鱼优势种和重要种的判断标准:取优势度大于1000的为优势种;鱼卵优势度500~1000为重要种,仔稚鱼优势度350~1000为重要种;鱼卵优势度50~500为一般种,仔稚鱼优势度50~350为一般种;优势度10~50为少见种;优势度小于10为稀有种。
1.3.3 鱼卵、仔稚鱼多样性指数
Shannon-Wiener多样性指数(H′)按下式计算:
式中,H′为多样性指数;S为物种总数;Pi为样品中第i种的丰度(ni)与各站位总丰度(N)比值。
Margale丰富度指数(D)按下式计算:
D=(S-1)/log2N
式中,D为丰富度指数;S为物种总数;N为各站位总丰度。
Pielou均匀度指数(J)按下式计算:
J=H′/log2S
式中,J为均匀度指数;H′为多样性指数;S为物种总数。
1.3.4 鱼卵、仔稚鱼群落结构分析
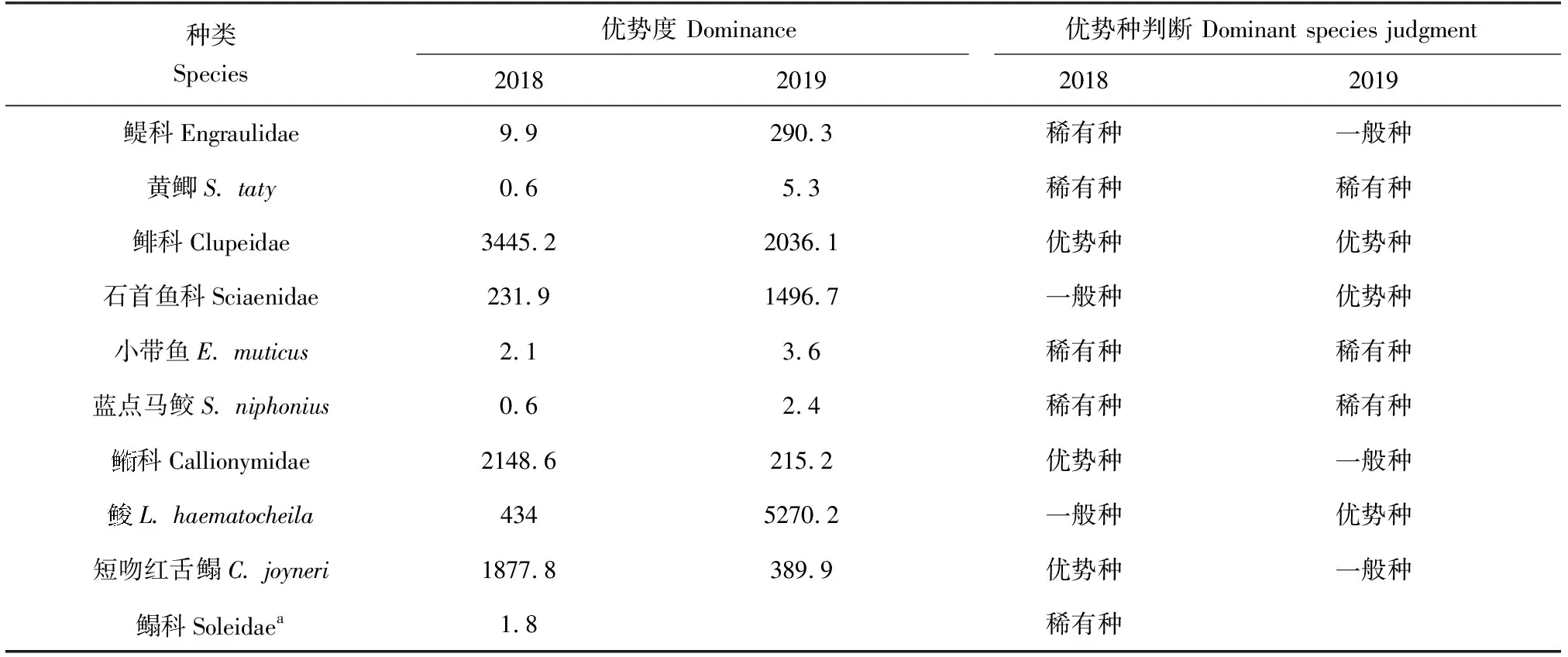
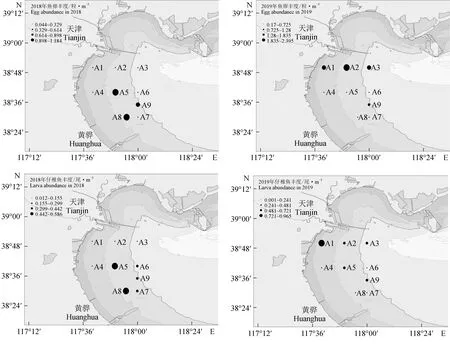
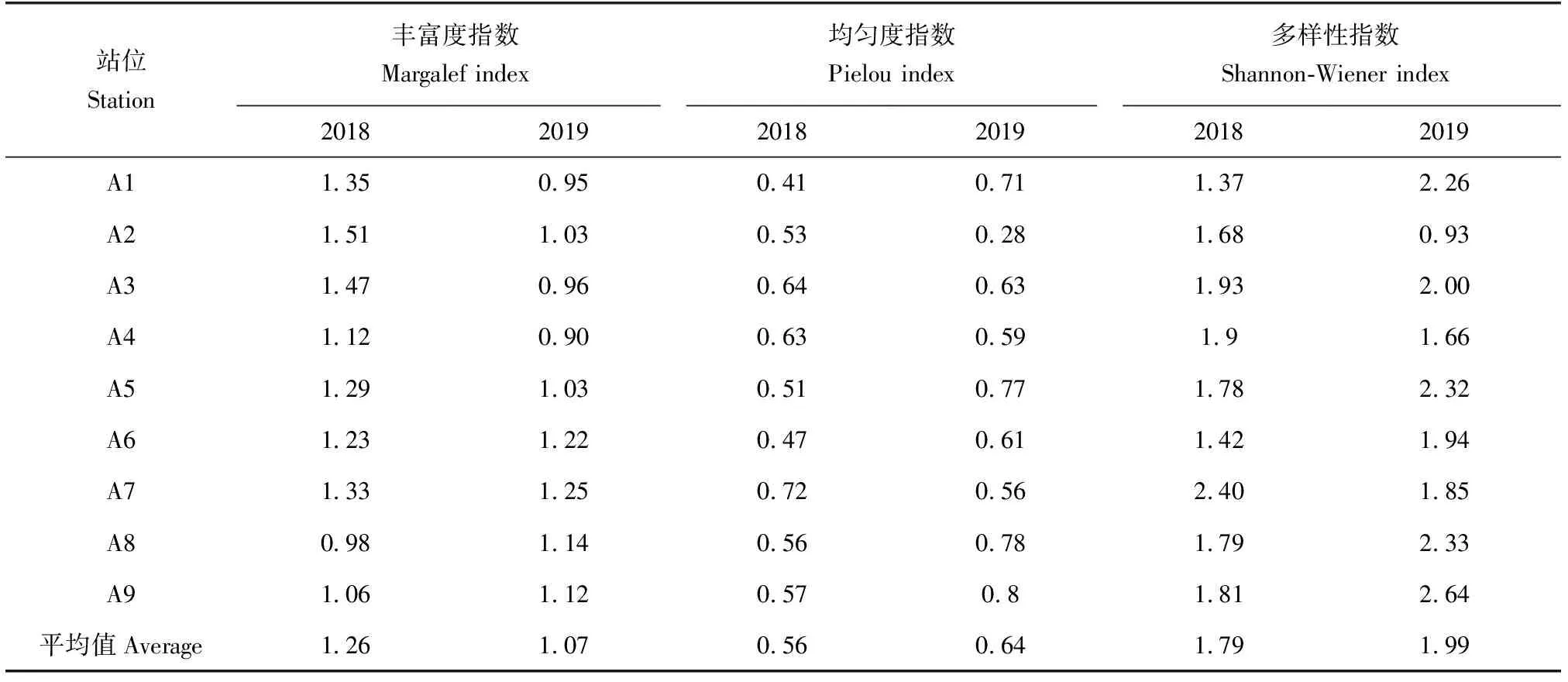
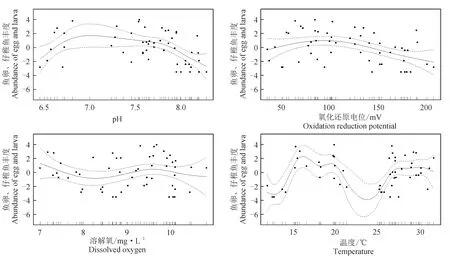
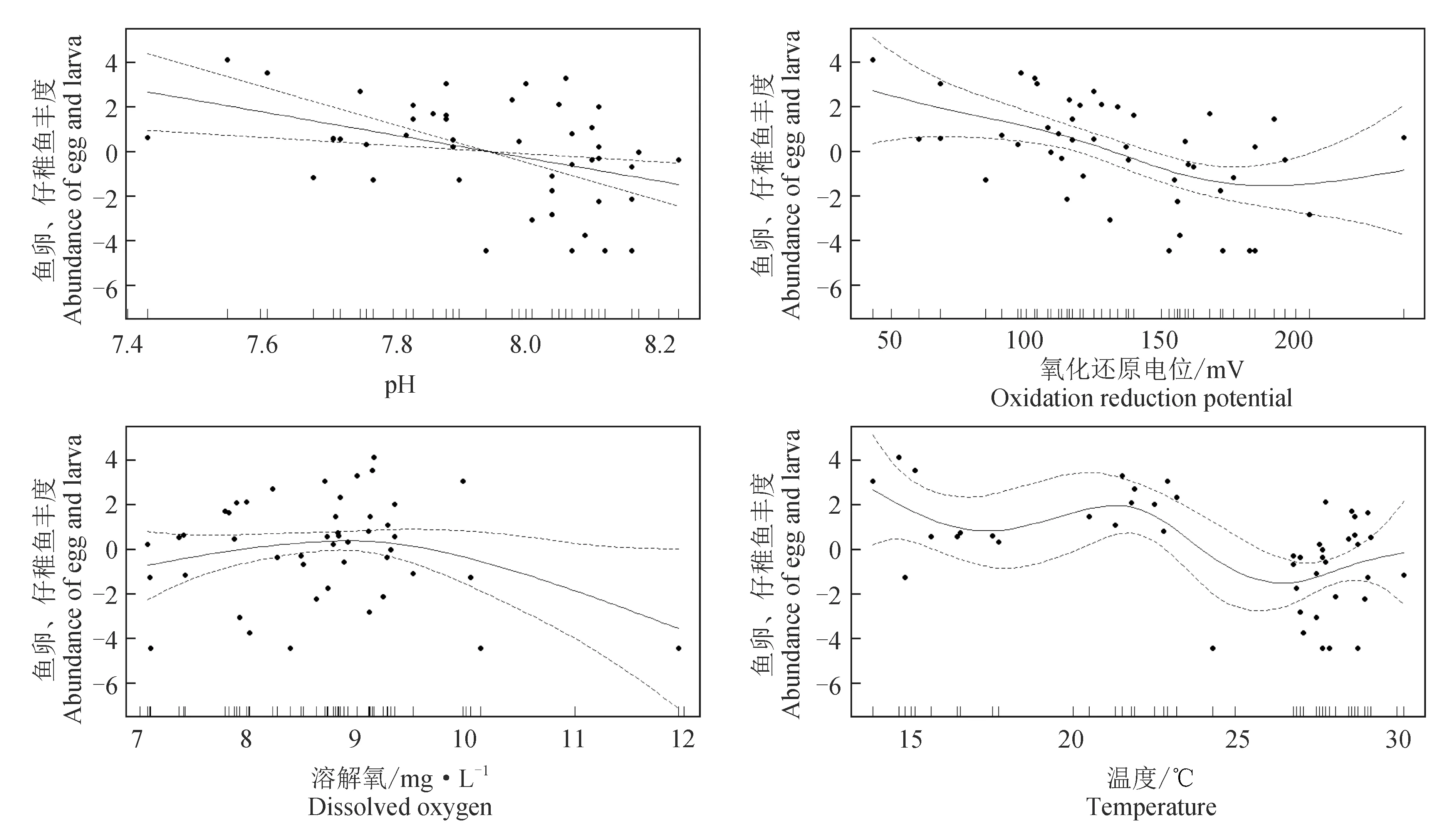
采用PRIMER 6.0进行多元统计分析:首先对物种的丰度数据进行对数转换log(x+1)并计算站位间的Bray-Curtis相似性系数,构建相似性矩阵,然后采用CLUSTER等级聚类方法和非度量多维标度(MDS)排序方法分析鱼卵、仔稚鱼的群落分布格局,并通过单因素相似性(ANOSIM)分析不同聚类组间的差异性。检验非度量多维标度排序结果的优劣,用协强系数(Stress)来衡量,当0 1.3.5 鱼卵、仔稚鱼与环境因子相关性分析 应用广义加性模型(GAM)分析鱼卵、仔稚鱼与影响因素之间的关系,模型表示为: 式中,Y为鱼卵、仔稚鱼、鱼卵和仔稚鱼总量;g(Y)为连接函数;xi为各环境因子;fi(xi)为g(Y)与第i个环境因子的非参数函数;k为环境因子个数;ε为误差项。 本研究中,连接函数为自然对数,模型采用样条平滑法,分布函数族为泊松分布,模型的误差分布估计为高斯分布,具体计算过程参见文献[17]中的方法,分析通过R 3.6.3软件及mgcv包实现。 2.1.1 鱼卵、仔稚鱼种类组成及种类数变化 2018年采集到鱼卵、仔稚鱼18种,包括鱼卵10种,隶属4目9科,仔稚鱼9种,隶属5目8科,其中10月、11月未采集到鱼卵,4月、9月、10月未采集到仔稚鱼;2019年采集到鱼卵、仔稚鱼16种,包括鱼卵9种,隶属4目8科,仔稚鱼8种,隶属5目7科,其中9月、10月、11月未采集到鱼卵,4月、10月、11月未采集到仔稚鱼。2018和2019年鱼卵、仔稚鱼共有种为15种(表1)。 表1 2018年和2019年天津近岸海域鱼卵、仔稚鱼种类及其生态特征Tab.1 Species and ecological characteristics of eggs and larvae in coastal waters of Tianjin in 2018 and 2019 随着季节的变更,2018、2019年鱼卵种类数整体呈先升后降的趋势,2018年仔稚鱼种类数随时间呈先升后降的趋势,而2019年则呈下降的趋势,表明2019年较2018年鱼卵、仔稚鱼种类数随季节变化更明显。随着区域的变动,自北部向南部延伸,2018年、2019年鱼卵种类数整体呈上升的趋势,2018年仔稚鱼种类数呈下降的趋势,而2019年则呈上升的趋势,表明2019年较2018年鱼卵、仔稚鱼种类数随空间变化更明显(图2)。 图2 2018、2019年鱼卵、仔稚鱼种类数随时间、空间变化Fig.2 The variation in the number of egg and larva species with time and space in 2018 and 2019 2.1.2 鱼卵、仔稚鱼优势种 对2018、2019年调查结果进行鱼卵、仔稚鱼优势度分析发现,2018年鱼卵优势种为3种,一般种2种,稀有种5种;2019年鱼卵优势种为3种,一般种3种,稀有种3种,共有鱼卵优势种为鲱科鱼类(表2)。 表2 2018年和2019年鱼卵优势种组成Tab.2 Dominant species composition of fish eggs in 2018 and 2019 2018年仔稚鱼优势种为3种,少见种2种,稀有种4种;2019年仔稚鱼优势种为2种,重要种3种,少见种2种,稀有种1种,共有仔稚鱼优势种为赤鼻棱鳀、有明银鱼(表3)。 表3 2018年和2019年仔稚鱼优势种组成Tab.3 Dominant species composition of larvae in 2018 and 2019 2.1.3 鱼卵、仔稚鱼丰度分布及变化 2018年和2019年分别采集鱼卵7397粒和18 339 粒,仔稚鱼3090尾和3954尾。两年度调查的鱼卵、仔稚鱼丰度空间分布不均匀,站位间有较大差距(图3)。2018年和2019年鱼卵平均丰度分别为0.418 粒/m3和0.961 粒/m3,仔稚鱼平均丰度分别为0.205 尾/m3和0.240 尾/m3。鱼卵、仔稚鱼年平均丰度趋势由远岸向近岸延展,高值区位于近岸区域。以年际与区域(A1~A3、A4~A6、A7~A9)为因素对鱼卵、仔稚鱼丰度进行双因素方差分析,结果显示,鱼卵丰度在年际及区域间均有显著性差异(F=4.843,P<0.05;F=14.212,P<0.01),仔稚鱼丰度在年际及区域间均无显著性差异(F=0.191,P>0.05;F=0.759,P>0.05)。 图3 2018、2019年鱼卵、仔稚鱼丰度空间分布Fig.3 Spatial distribution of abundance of egg and larvae in 2018 and 2019 随着季节的变更,2018、2019年鱼卵、仔稚鱼丰度整体呈下降的趋势,2019年较2018年鱼卵、仔稚鱼丰度随季节变化更明显。随着区域的变动,自北部向南部延伸,2018年鱼卵、仔稚鱼丰度整体呈上升的趋势,2019年鱼卵、仔稚鱼丰度整体呈下降的趋势,2019年较2018年鱼卵、仔稚鱼丰度随空间变化更明显(图4)。 图4 2018年和2019年鱼卵、仔稚鱼丰度随时间、空间变化Fig.4 Variation in abundance of eggs and larvae with time and space in 2018 and 2019 2.1.4 鱼卵、仔稚鱼多样性变化 天津近海鱼卵仔稚鱼丰富度指数、均匀度指数和多样性指数在不同年际间无显著性差异(P>0.05)(表4),2019年丰富度指数平均值低于2018年,是因为捕获到的鱼卵、仔稚鱼种类数低于2018年;均匀度指数和多样性指数平均值高于2018年,是因为捕获到的鱼卵、仔稚鱼的数量高于2018年,从整体来看,2019年较2018年鱼卵、仔稚鱼丰度更高、个体分布更为均匀。 表4 天津近海鱼卵、仔稚鱼物种多样性变化 聚类分析结果表明(图5a、图5b),2018年鱼卵、仔稚鱼群落在61%的相似水平上可分为3组,分别为A组、B组和C组,多维尺度排序分析结果协强系数(Stress=0.05)显示,多维尺度排序分析二维点图对群落的划分具有很好的代表性,单因素相似性分析表明,鱼卵、仔稚鱼群落之间差异极显著(R=0.926,P=0.004)。其中A、B、C 3组均为3个站位,区域分布明显,分组等级一致(Pairwise Test)。 聚类分析结果表明(图5c、图5d),2019年鱼卵、仔稚鱼群落在65%的相似水平上可分为2组,分别为A组、B组,多维尺度排序分析排序结果协强系数(Stress=0.06)显示,多维尺度排序二维点图对群落的划分代表性较差,单因素相似性分析表明,鱼卵、仔稚鱼群落之间差异极显著(R=0.775,P=0.008)。其中A组为4个站位,区域分布北部集中,分组等级较高(Pairwise Test)。B组分别为5个站位,区域分布南部集中,分组等级较高(Pairwise Test)。 图5 2018年、2019年鱼卵、仔稚鱼群落聚类分析和多维尺度排序分析示意Fig.5 Cluster analysis and multi-dimensional scale sequencing analysis of eggs and larvae in 2018 and 2019 基于2018年和2019年鱼卵、仔稚鱼丰度及环境调查结果,运用广义加性模型分析鱼卵、仔稚鱼丰度与温度、氧化还原电位、溶解氧、pH等环境因子的关系(表5)。结果表明,除溶解氧外(P>0.05),温度、pH、氧化还原电位对2018年鱼卵、仔稚鱼丰度有显著影响(P<0.01),解释变量依次为47.9%、33.5%、22.8%。除溶解氧外(P>0.05),温度、氧化还原电位、pH对2019年鱼卵、仔稚鱼丰度有显著影响(P<0.01),解释变量依次为40.3%、32.1%、17.9%。 表5 鱼卵、仔稚鱼丰度与相关环境因子广义加性模型的解释偏差及显著性Tab.5 Explanatory deviation and significance in GAM of abundance of eggs, larvae and related environmental factors 基于鱼卵、仔稚鱼丰度与环境因子的广义加性模型预测结果可见,除溶解氧外,pH、氧化还原电位、温度对2018、2019年鱼卵、仔稚鱼丰度预测的置信区间小,预测的可靠度高,相关环境因子对2018年鱼卵、仔稚鱼丰度的影响较2019年大。随着pH、氧化还原电位、温度的升高,鱼卵、仔稚鱼丰度整体上呈降低的趋势(图6、图7)。 图6 2018年鱼卵、仔稚鱼丰度与环境因子广义加性模型分析Fig.6 GAM analysis of abundance of egg and larva and environmental factors in 2018 图7 2019年鱼卵、仔稚鱼丰度与环境因子广义加性模型分析Fig.7 GAM analysis of abundance of egg and larva and environmental factors in 2019 结合20世纪80年代天津市海岸带和海涂资源综合调查[1]和2008年天津海域鱼卵、仔稚鱼调查[25]数据可知,20世纪80年代鱼卵、仔稚鱼共33种,2008年共25种,而本次调查2018年共采集到18种,2019年共采集到16种。从较长历史跨度分析,2008年和本次调查较20世纪80年代共同减少的种类为褐牙鲆(Paralichthysolivaceus)、鳓(Ilishaelongata)、褐菖鲉(Sebastiscusmarmoratus)、红娘鱼(Lepidotriglasp.)和长绵鳚(Zoarceselongatus,长绵鳚产卵期为12至翌年2月份,本次调查未在其产卵期),而本次调查较20世纪80年代减少种类为钝尖尾虾虎鱼(C.hexanema)、中华栉孔虾虎鱼(Ctenotrypauchenchinensis)、竿虾虎鱼(Luciogobiusguttatus)、黄姑鱼(Nibeaalbiflora)、小黄鱼(Larimichthyspolyactis)、黑鳃梅童鱼(Collichthysniveatus)、皮氏叫姑鱼(Johniusbelengerii)、白姑鱼(Argyrosomusargentatus)、多鳞(Sillagosihama)、银汉鱼(Allanettableekeri)和尖海龙(Syngnathusacus)。增加的种类有有明银鱼、矛尾虾虎鱼、髭缟虾虎鱼。再结合20世纪80年代至今的调查资料,可知渤海渔业资源衰退,资源结构发生了很大的变化,种类交替明显[26],天津近岸海域产卵场功能有衰退的迹象。 通过研究2018年和2019年天津近岸海域鱼卵、仔稚鱼群落结构发现,由于2019年捕获到的鱼卵、仔稚鱼种类数低于2018年,导致其丰富度指数较低,但从均匀度指数和多样性指数来看,2019年较2018年鱼卵和仔稚鱼丰度更高、个体分布更为均匀。再结合聚类分析及单因素相似性分析可知,从鱼卵、仔稚鱼丰度结果上可将2018年、2019年鱼卵、仔稚鱼群落按站位依次分为3组、2组,2018年的3组均为3个站位,区域分布明显,分组等级一致。2019年的2组分别为4、5个站位,区域分布两极分化,分组等级较高。表明不同年份的相同区域鱼卵、仔稚鱼丰度均不相同,受空间影响较大。相关研究也表明,由于不同区域水深、海流以及一些环境因子的差异,使鱼卵、仔稚鱼群落结构呈不同的表现[33],因此需要对不同区域条件的差异情况进行总结来进一步分析。 2018年和2019年鱼卵总平均丰度分别为0.418 粒/m3和0.961 粒/m3;仔稚鱼总平均丰度分别为0.205 尾/m3和0.240 尾/m3。根据报道,1999—2002年[34]及2008年渤海湾鱼卵及仔稚鱼丰度分别为6.05、2.67、2.40、6.0、2.88 个/m3,从丰度数据上看,本次调查明显少于历史调查数据,表明天津近岸海域产卵场功能衰退的迹象已经非常严重。从丰度的季节、空间变化来看,春、夏两季为天津近岸海域的产卵盛期,鱼卵、仔稚鱼丰度受季节变动大,2019年较2018年鱼卵、仔稚鱼丰度随季节变化幅度更明显;随着调查区域由北部向南部延伸,鱼卵、仔稚鱼丰度整体呈上升趋势,2018年鱼卵、仔稚鱼丰度整体呈上升的趋势,2019年鱼卵、仔稚鱼丰度整体呈下降的趋势,表明不同年份相同区域的鱼卵、仔稚鱼丰度呈两极变化。因产卵场水质环境下降[35]及鱼类资源衰退严重[36],2018年鱼卵、仔稚鱼丰度的下降符合历年来鱼卵、仔稚鱼数量的衰退现象;而2019年鱼卵、仔稚鱼丰度的上升可能由于禁渔期提前,滥捕的现象得到制约,鱼卵、仔稚鱼资源得到了一定的恢复。 海洋环境是海洋生物的生产基础,对海洋生物生长的各个阶段都具有重要影响[37]。影响鱼卵孵化和仔鱼发育的主要影响因子主要是溶解氧含量、温度、pH等[38]。大量研究表明,温度为鱼卵、仔稚鱼的主要影响因素[22,27],此外鱼卵无游泳能力,仔稚鱼的游泳能力弱,海流也是影响其分布的重要因素,但影响范围及程度尚无准确的结论。 笔者选取了pH、氧化还原电位、溶解氧含量和温度等环境因子,结合鱼卵、仔稚鱼丰度进行广义加性模型分析。结果表明,温度、氧化还原电位和pH为鱼卵、仔稚鱼丰度的主要影响因素(P<0.001),随着pH、氧化还原电位、温度的升高,鱼卵、仔稚鱼丰度整体上呈降低的趋势,与王麒麟等[28]认为鱼卵、仔稚鱼丰度与温度呈显著正相关的结果相反,与肖瑜璋等[2]的研究结果相似。宋秀凯等[21]研究表明,渤海莱州湾鱼卵、仔稚鱼丰度与温度无相关性;樊紫薇等[39]研究表明,东海鱼卵、仔稚鱼丰度与温度呈负相关。这些报道多是研究鱼卵、仔稚鱼丰度与环境因子的关联程度,未从丰度的整体变化来综合考虑,因此运用广义加性模型分析鱼卵、仔稚鱼丰度与环境因子的整体变动,更能体现鱼卵、仔稚鱼与环境因子的关系。 2018年共采集到鱼卵7397粒,仔稚鱼3090尾;2019年共采集到鱼卵18 339粒,仔稚鱼3954尾。与1999—2002年及2008年数据相比,本次调查鱼卵、仔稚鱼丰度明显少于历史调查数据,鱼卵、仔稚鱼丰度也受季节和空间变动影响,天津近岸海域产卵场功能衰退的迹象已经非常严重。将pH、氧化还原电位、溶解氧含量和温度等环境因子结合鱼卵、仔稚鱼丰度进行广义加性模型分析,结果显示,温度、氧化还原电位和pH鱼卵、仔稚鱼丰度的主要影响因素(P<0.001)。 对天津近岸海域鱼卵、仔稚鱼和环境因子进行周年监测,将其变化情况可视化,一方面为渔业资源的可持续开发利用提供科学资料,另一方面了解鱼卵、仔稚鱼群落变化的规律和影响因素,可适时进行一系列的措施来应对鱼卵、仔稚鱼群落的剧烈变化:(1)应当延长禁渔期与加大执法力度来保障天津近岸渔业资源的恢复;(2)对越冬场、春季产卵场等重要海域实行常年禁渔,促进资源恢复;(3)减少或禁止围填海工程对海洋资源环境的破坏。2 结果与分析
2.1 天津近岸海域鱼卵、仔稚鱼群落结构特征




2.2 天津近岸海域鱼卵、仔稚鱼群落结构分析

2.3 环境因子相关性分析

3 讨 论
3.1 天津近岸鱼卵、仔稚鱼种类、优势种及群落结构变化
3.2 鱼卵、仔稚鱼丰度变化及其与环境因子的关系
4 结 论
猜你喜欢
渔业科学进展(2021年6期)2022-01-04 06:26:42
今日农业(2021年8期)2021-11-28 05:07:50
水上消防(2021年3期)2021-08-21 03:12:00
浙江海洋大学学报(自然科学版)(2021年1期)2021-07-06 06:58:28
食品工业(2020年6期)2020-07-18 04:07:20
创新作文(3-4年级)(2019年8期)2019-12-17 07:59:21
人大建设(2019年5期)2019-10-08 08:55:10
创新作文(小学版)(2019年23期)2019-01-11 10:37:45
海洋渔业(2016年6期)2016-04-16 03:00:32
应用海洋学学报(2015年3期)2015-11-22 07:39:22
