
截顶后红松幼树光合生理响应研究
2017-06-05 14:20NguyenThanhTuan沈海龙王琴香张鹏VuDinhDuy
森林工程 2017年4期
Nguyen Thanh Tuan,沈海龙,王琴香,张鹏,Vu Dinh Duy
(1.东北林业大学 林学院,哈尔滨 150040;2.越南林业大学第二分校 林学系,越南 同奈 810000;3.越南技术与科学院 自然博物馆,越南 河内 100803)
截顶后红松幼树光合生理响应研究
Nguyen Thanh Tuan1,2,沈海龙1*,王琴香1,张鹏1,Vu Dinh Duy3
(1.东北林业大学 林学院,哈尔滨 150040;2.越南林业大学第二分校 林学系,越南 同奈 810000;3.越南技术与科学院 自然博物馆,越南 河内 100803)
以14年生红松人工林为研究对象,研究不同截顶处理(CK:不截顶、P1:截去第2轮枝及以上部分、P2:截去第3轮枝及以上部分)对树木光合生理特征的影响。结果表明:截去第3轮枝后在短时间内(2个月)对红松树体光合特征各指标有显著提高作用,但是效应随着时间增加而减少,2个月后影响就不再显著;截顶后不同叶龄针叶的光合生理响应也有所不同:对于当年生针叶,两种截顶处理对净光合速率、气孔导度、胞间CO2浓度的影响不显著,但两种截顶对蒸腾速率影响显著,并且P2处理显著提高了蒸腾速率;对于一年生针叶,两种处理中P1处理只显著提高了净光合速率,P2处理对所有光合指标都没有产生显著影响;对于二年生针叶,P2处理显著增加了针叶的各光合特征指标(净光合速率、气孔导度、蒸腾速率和胞间CO2浓度),而P1处理对所有光合指标的影响都不显著;截去第3轮枝的红松一年生、二年生针叶叶绿素a、叶绿素b、总叶绿素及类胡萝卜素含量显著高于未截顶红松。本试验揭示了截去第3轮枝及以上部分可以增加红松叶片光合作用,可为进一步探究截顶后树木的复壮机制以及改善经营水平提供依据。
红松(Pinus koraiensis);截顶;光合特征;色素含量
0 引言
树木的胸径、树高、材积、冠幅和叶面积指数等生长特性因子与其生理特性密切相关,也是树木生理特性的表现结果。因此,研究树木生理指标对环境的变化,不仅能了解树木生长发育的规律,也可以揭示树木生长发育过程的机理[1]。光合特性是植物利用太阳辐射进行光合作用后转化为干物质能力的重要反映,而树冠是林木进行光合作用的主要部分,其他部位对光合速率影响不大[2],树冠的特征则直接影响着林木的生长发育,修剪去梢是调控林木冠层的关键手段[3]。国外有关于修剪去梢对树木的生理研究已经被广泛关注[4-10],国内对其研究开展较晚,多集中于用材林树种的研究,而且关于树冠调控研究成果在用材林树种定向培育中的应用推广率仍较低[3]。
红松(PinuskoraiensisSieb.etZucc.)是东北地区的多功能乡土树种,其材质轻软,结构细腻,形色美观又不容易变形,并且耐腐朽力强,是建筑、桥梁、枕木、家具制作的上等木材。此外,红松种子具有较高的食用价值和滋补功能,越来越受到市场的欢迎[11]。近些年来出现了针对红松人工林进行的双向培育、果材兼用培育等理论、实践和科学研究,多地正着手改建红松人工林成为红松果材兼用林,而改建的主要技术措施就是截顶促进及调控杈干发生[12-15]。为了寻找最佳的果材兼用林培育技术和科学依据,国内学者已对红松截顶展开了研究[16]。不过,目前的研究主要集中在截干对红松生长结实方面的影响,对红松截干后其生理特征的变化研究较少。探讨截顶对红松生理特征的影响,能从生理角度上对一些树木生长特性的变化作出解释,因此有着重要而深远的意义。
本文以红松人工林为研究对象,通过设置不同截顶处理,探索截干对树木光合生理的影响,从光合生理角度出发解释截干对红松生长的影响,以期为今后红松果材兼用林定向培育的截干技术给予理论支持。
1 研究方法
1.1 试验设计
研究地点位于黑龙江省东北林业大学帽儿山实验林场王家沟红松更新试验区,地理坐标:127°30′~127°34′E,45°21′~45°25′N,海拔高度500 m左右,属温带大陆性季风气候区。该地区年平均气温2.8 ℃,最低温度在1月,为 -19.6 ℃,7月平均温度20.9 ℃。年平均降水量723 mm。
针对试验地14年生红松幼树,设置3个截顶处理:截去第二轮枝及以上部分的处理(P1)、截去第三轮枝及以上部分的处理(P2)、未截顶对照(CK)。每个处理选取3株,一共9株树木,截顶前测定实验区各处理树木的基本生长指标(表1)。于2015年5月中旬按照实验设计进行截顶处理,并测定光合作用各项指标,取样时间为:2015年6月中旬,7月中旬,8月中旬,9月中旬。

表1 王家沟试验地红松幼树截顶前基本生长指标
1.2 试验方法
1.2.1 树体光合特征的测定
于2015年6月到9月进行光合特征指标测定。为保证测定的准确性,在每株红松的树体中部向阳方向上选择生长状况良好及针叶数量比较多的一个侧枝,并采摘健康的各年生针叶来测定光响应,以此来保证实验的顺利进行和实验数据的可靠性。测光响应曲线尽量选择在晴朗且云量较少的天气,测定时间为上午9:00-12:00,第一株测定完毕之后再进行下一株的测定,尽量缩短同一株树不同叶龄间测定时间的差异。每个侧枝每个叶龄选择几束针叶并做好标记以便下次测定。
LI-COR公司生产的Li-6400光合测定仪,用Li-6400-02B红蓝光源提供标准光强,光强度设置为1 000μmol/(m2.s);CO2浓度为350 L/L;温度根据当时的环境条件,设定为25 ℃,读取6个重复。测量的光合参数值主要是叶片单位面积的净光合速率(Pn)、气孔导度(Cond)、蒸腾速率(Tr)、胞间CO2浓度(Ci)等。结果取实测数据的平均值。
1.2.2 针叶色素含量的测定
从6月份到9月份,每月分别从9株树木的各轮枝层摘取不同龄级的针叶后,放入有冰块的保温箱,带回实验室用于测定色素含量。叶样取回后随即采用Arnon方法进行针叶色素含量的测定。叶样不用研磨,直接将针叶剪碎后称取0.3 g置于20 ml体积分数为80%的丙酮溶液中,放在避光的冰盒中提取12 h,期间摇动3~4次,直到碎片完全变白为止。样品直接取浸提的上清液用紫外分光光度计(Hitachi-U2001;Hitachi,Tokyo,Japan)进行比色测定,在波长663、645、470 nm下测定吸光度,每个样品测定3次,结果取实际测得数据的平均值。通过吸光度值计算叶绿素a、b的浓度及其叶绿素总浓度和类胡萝卜素的浓度,公式如下:
叶绿素a (mg/L):
Chla=12.21×A663-2.69×A645。
(1)
叶绿素b (mg/L):
Chlb=22.9×A645-24.68×A663。
(2)
总叶绿素 (mg/L):
CT= Chla+Chlb。
(3)
类胡萝卜素(mg/L):
Car= (1 000×A470-3.27×Chla-104×Chlb)/229。
(4)
其中A663、A645、A470分别为提取液在波长663、645、470 nm出的吸光度值。
本研究进行一下换算来计算单位质量的色素含量,公式如下:
单位质量色素含量(mg/g)= (溶液中色素浓度×提取液总量) /样品鲜重。
(5)
所有数据的统计以及图表绘制均使用 Microsoft Excel 2007和Sigmaplot软件完成,采用SPSS 21.0进行方差分析和多重比较等分析。
2 结果与分析
2.1 截顶对红松树体光合特征的影响
不同处理对当年生、一年生和二年生叶龄针叶的光合特性指标季节变化情况及方差分析结果如图1所示。由图1可以看出,从6月中旬到9月中旬,不同处理间各叶龄的针叶净光合速率均显著呈现逐渐增加的趋势,在9月份达到最大值,而且当年生针叶的净光合速率比其他叶龄针叶的都要大。由7月和9月两次的观测结果可以看出,截顶对当年生针叶的净光合速率影响不显著(P>0.05)。而对于一年生针叶,6月各处理间的净光合速率差异显著(P<0.05),光合速率:P1>CK>P2。同样,6月份各处理间的二年生针叶净光合速率差异也达到了显著水平(P<0.05),光合速率:P2>CK>P1。7月、9月各处理间的一年生和二年生针叶净光合速率差异均未达到显著水平(P>0.05)。总体来看,截顶在短时间内会(1个月内)对红松一年生和二年生的针叶净光合速率产生显著增加的影响,而从2个月后这种影响就不显著了。
气孔导度是表示气孔张开程度的一个指标,它是植物叶片与外界进行气体交换的主要通道。气孔导度直接影响树木光合作用、呼吸作用及蒸腾作用。由图1可知,当年生针叶的气孔导度高于一年生和二年生的针叶。不同处理的一年生和二年生针叶在不同季节的变化规律呈现单峰曲线,气孔导度值显著:7月>9月>6月。相反的是,7月的当年生针叶气孔导度却小于9月的。截顶对红松一年生针叶的气孔导度影响不显著(P>0.05),而在7月,截顶对红松当年生、二年生针叶的气孔导度均产生显著影响(P<0.05),而且处理P2的气孔导度值最大。
蒸腾速率是植物在一定的时间内单位面积蒸腾的水量。由结果分析可知,从季节方面来看,红松一年生、二年生针叶的蒸腾速率在6月份最小,7月份最大,到9月蒸腾速率降低,变化规律为:7月份>9月份>6月份,且当年生针叶7月的蒸腾速率高于9月。从当年生的红松针叶来看,截顶对蒸腾速率产生显著影响,在7月、9月各处理间蒸腾速率为:P2>CK>P1,且差异达到显著水平(P<0.05)。在6月,各处理间的一年生针叶蒸腾速率差异显著(P<0.05):CK>P1>P2,除了6月份以外,在其余时间观察截顶对一年生针叶的蒸腾速率均无影响。从二年生针叶来看,在7月份各处理的蒸腾速率差异显著,而且P2处理的蒸腾速率最高(P<0.05)。
胞间CO2浓度变化是分析光合速率和气孔导度变化之间的因果关系重要的基础。各处理当年生、一年生针叶的胞间CO2浓度变化规律不同。而对于二年生针叶,7月、9月的胞间CO2浓度均显著高于6月,从6月到7月改变较为明显。通过方差分析不同处理对各叶龄针叶的胞间CO2浓度影响表明:截顶对当年生针叶的胞间CO2浓度均不产生影响(P>0.05);在7月测定结果表明,CK、P1的一年生针叶胞间CO2浓度都显著高于P2(P<0.05);从二年生针叶看来,6月各处理的胞间CO2浓度差异显著(P<0.05),即P2>P1>CK。
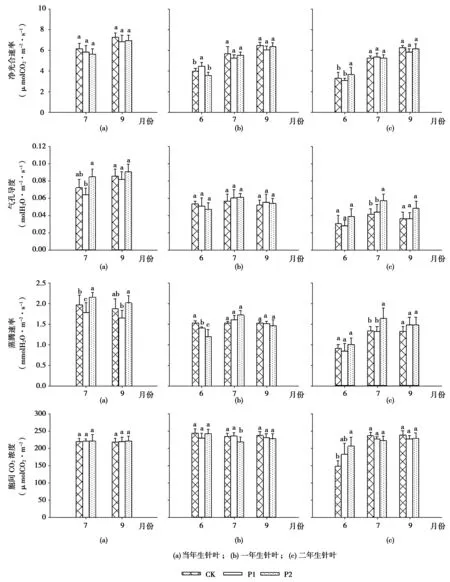
图1 截顶对红松幼树针叶光合特征的影响Fig.1 Effect of top pruning on photosynthetic characteristics of Pinus koraiensis needles
2.2 截顶对红松针叶色素含量的影响
叶绿素是绿色植物进行光合作用的主要色素,叶绿素含量增加有利于光能的吸收和转化。其中,叶绿素a有利于吸收长波光,叶绿素b有利于吸收短波光。截顶后各处理红松针叶色素含量指标变化情况如图2所示。
从当年生针叶色素含量变化来看,3种处理红松针叶叶绿素a、叶绿素b、总叶绿素、类胡萝卜素差异不显著(P>0.05),说明截顶对红松当年生针叶的色素含量末产生影响。

图2 截顶后红松幼树不同叶龄针叶的色素含量Fig.2 Pigment content of different age needles for Pinus koraiensis after top pruning
从一年生针叶响应来看,在7月,截顶对红松一年生针叶各色素含量指标均产生影响,而且P2处理与CK相比,一年生针叶的叶绿素a、叶绿素b、总叶绿素、类胡萝卜素分别增加了38.32%(P<0.05)、59.99%(P<0.05)、43.90%(P<0.05)、32.92%(P<0.05),P1和与CK之间的一年生针叶色素含量差异不显著(P>0.05)。此外,8月P2处理与P1处理相比,一年生针叶的类胡萝卜素增加了54.09%(P<0.05)。
从二年生的红松针叶角度来看,从6月份到9月份P1处理和CK对照之间的叶绿素a、叶绿素b、总叶绿素、类胡萝卜含量差异均不显著。而对于叶绿素a而言,除了7月以外,其他月份P2处理与对照之间的差异均显著,并且6、8、9月P2处理与对照CK相比,二年生红松叶片叶绿素a分别增加了55.44%(P<0.05)、49.45%(P<0.05)、11.61%(P<0.05)。在6月和8月,P2处理与对照之间叶绿素b差异显著,7月和9月差异不显著。6、8月P2处理与CK对照相比,二年生针叶叶绿素b增加了66.20%(P<0.05)、102.48%(P<0.05)。9月P2处理与P1处理相比,二年生针叶叶绿素b增加了52.41%(P<0.05)。截顶对红松二年生针叶的总叶绿素含量影响显著(P<0.05)。6月份、8月份P2处理与CK对照相比,针叶总叶绿素含量增加了58.65%(P<0.05)、63.88%(P<0.05)。除了8月份以外,其余月份各处理间红松针叶类胡萝卜素含量差异显著。6月P2处理与CK对照相比,二年生红松叶片类胡萝卜素含量增加了53.39%(P<0.05)。7、9月P2处理与P1处理相比,类胡萝卜素含量分别增加了41.48%(P<0.05)、83.02%(P<0.05)。
总体来看,除了当年生的红松针叶外,截顶对红松针叶色素含量产生显著影响,而截去第三轮枝处理后可以提高红松针叶的各种色素含量。
3 讨论与结论
光合作用及其效率的大小是植物生长的关键因素,决定着植物产量的高低[17]。此外,光合作用以树木消耗水分和CO2为主要生理活动,直接影响林木生长状况。通过测定树木光合特性,可以得到林木整体生长变化情况的信息。本研究通过截去第二轮枝、第三轮枝后,分析截顶对14年生红松针叶光合参数、色素含量的影响,初步揭示了红松对截干的光合生理响应特征。
以往很多研究表明树冠修剪对树木的光合特征产生影响[10,18-19]。与本研究结果一致,本研究结果表明截顶对红松净光合速率、蒸腾速率、气孔导度和胞间CO2浓度等生理指标均产生影响,而且各叶龄的针叶对截顶的反应有所不同,截顶对当年生针叶的净光合速率、胞间CO2浓度未产生影响,而对一年生和二年生针叶光合特性各指标却有显著影响。树冠修剪后在树木生理特征上具有一种短期效应趋势[20-21],本实验对光合速率反应的研究也发现,截顶会在短时间内(1个月)对红松净光合速率显著增加作用,但随着时间增加影响作用在不断减小,到2个月后影响就不再显著。不同叶龄红松叶片光合速率大小不同,依次为当年生叶>一年生叶>二年生叶,截去第二轮枝能提高红松一年生针叶的光合速率,而截去第3轮枝能提高红松二年生针叶的光合速率,说明树冠修剪能增加叶片光合速率,这是与尚富华[22]、Forrester et al.[23]、泰柱南[1]等研究的结论相一致。
树冠修剪后叶片的气孔密度增加而使叶片气孔导度增加,提高其光合速率[5,24]。与前人研究一致,本研究发现截去第3轮枝能提高二年生针叶的气孔导度,同时增加其生光合速率。净光合速率是气孔导度、胞间CO2浓度等共同作用的结果,而且胞间CO2浓度作为光合过程中CO2的中介,受到气孔导度和光合速率两个因素的互相限制[25-26]。本研究结果也显示截去第3轮枝后红松二年生针叶的胞间CO2浓度增加显著,说明胞间CO2浓度的增加主要由截顶后叶片光合速率和气孔导度改变的综合作用而引起。从上述结果可知,净光合速率与叶片气孔导度以及蒸腾速率之间均呈极显著正相关,符合生理学上三个指标的正常关系[27-29]。这说明红松幼树截顶后,从针叶生理功能方面改善了整个树木的光合作用性能。不过,还尚不清楚这是否与截顶后红松叶片的生理结构有了改善,从而提高了其生理活动能力有关联,宜继续扩大研究范围。
关于树冠修剪后树木自身恢复平衡状态的机制存在很多争议,有学者曾经认为光合速率增加与叶片氮浓度有关,但也有一些学者不赞同这个观点,他们通过研究却没找到光合速率增加与叶片氮浓度有任何相关的证据[5,30]。也有相关研究表明通过树冠修剪能增加叶片的叶绿素含量,从而影响树木吸收CO2能力,而且叶绿素是参与光合作用中光能吸收、传递和转化的重要色素[31]。尚富华研究表明修枝提高了叶绿素含量,而且叶绿素含量与光合能力相联系表现为呈正相关[22]。与前人相同、本研究结果表明,截去第3轮枝的红松一年生、二年生针叶叶绿素a、b、总叶绿素显著高于未截顶红松,说明截顶能提高针叶叶绿素含量。叶片是树木进行光合作用的主要器官,而叶绿体是含有叶绿素能进行光合作用的细胞器。叶绿体的色素有下列两类:叶绿素和类胡萝卜素,在这两类色素中,叶绿素是最主要的,而且叶绿素具有接受和转换能量的作用,在一定的范围内,植物叶片叶绿素含量越高,光合作用越强[2]。从理论上和实验结果证明,截顶有助于叶绿素的积累,提高树木光合作用。
总体上是截顶初期各效应有所提高,中后期提升作用消失,但规律性趋势不明显;截去第3轮枝显著提高红松叶片光合特征及叶绿素含量。
[1]秦柱南.修枝对L35杨的生长生理特性及林下间作物产量影响的研究[D].济南:山东农业大学,2014.
[2]潘瑞炽,董愚得.植物生理学[M].北京:高等教育出版社,2001.
[3]任世奇,陈健波,周维,等.修枝对尾巨桉生长及光合生理的影响[J].生态学杂志,2013,32(11):2978-2984.
[4]Pinkard E A,Battaglia M,Beadle C L,et al.Modeling the Effect of Physiological Responses to Green Pruning on Net Biomass Production ofEucalyptusnitens[J].Tree Physiology,1999,19(1):1-12.
[5]Pinkard E A,Beadle C L.Regulation of Photosynthesis inEucalyptusnitens(Deane and Maiden)Maiden Following Green Pruning [J].Trees,1998,12(6):366-376.
[6]Neilsen W A,Pinkard E A.Effects of Green Pruning on Growth ofPinusradiata[J].Canadian Journal of Forest Research,2011,33(11):2067-2073.
[7]Bayala J,Teklehaimanot Z,Ouedraogo SJ.Millet Production Under Pruned Tree Crowns in a Parkland System in Burkina Faso[J].Agroforestry Systems,2002,54(3):203-214
[8]Frank B,Eduardo S.Biomass Dynamics ofErythrinalanceolata,as Influenced by Shoot-pruning Intensity in Costa Rica[J].Agroforestry Systems,2003,57(1):19-28.
[9]Medhurst J L,Pinkard E A,Beadle C L,et al.Photosynthetic Capacity increases inAcaciaMelanoxylon,Following Form Pruning in a Two-species Plantation[J].Forest Ecology & Management,2006,233(2-3):250-259.
[10]Philipj A,Jürgen B,Danes T,et al.Photosynthetic Response to Green Crown Pruning in Young Plantation-grownEucalyptuspilularisandEucalyptuscloeziana[J].Forest Ecology & Management,2008,255(11):3827-3838.
[11]Wang Z Y,Chen X Q.Functional Evaluation for Effective Compositions in Seed Oil of Korean pine[J].Journal of Forestry Research,2004,15(3):215-217.
[12]齐鸿儒.红松人工林[M].北京:中国林业出版社,1991.
[13]倪柏春,倪薇,郑在军,等.空间指数图在红松人工林生长抚育中的应用[J].林业科技,2012,37(4):44-46.
[14]张利民,王行轩,王玉光.红松生长结实与分杈关系的研究[J].辽宁林业科技,2002(5):19-36.
[15]谭学仁,胡万良,王忠利,等.红松人工林大径材培育及种材兼用效果分析[J].东北林业大学学报,2000,28(3):75-77.
[16]沈海龙,张金虎,王龙.红松分杈现象研究现状及展望[J].森林工程,2015,31(2):46-50.
[17]Raines C A.Increasing Photosynthetic Carbon Assimilation in C3 Plants to Improve Crop Yield:Current and Future Strategies[M].The time of the goats.St.Clement of Ohrid,National and University Library,2011.
[18]Lisboa M,Acuna E,Cancino J,et al.Physiological Response to Pruning Severity inEucalyptusRegnans,Plantations[J].New Forests,2014,45(6):753-764.
[19]Barry K M,Pinkard E A.Growth and Photosynthetic Responses Following Defoliation and Bud Removal in Eucalypts[J].Forest Ecology & Management,2013,293(2):9-16.
[20]Pinkard E A,Battaglia M,Mohammed CL.Defoliation and Nitrogen Effects on Photosynthesis and Growth ofEucalyptusGlobulus[J].Tree Physiology,2007,27(7):1053-1063.
[21]Eyles A,Pinkard E A,Mohammed C.Shifts in Biomass and Resource Allocation Patterns Following Defoliation inEucalyptusGlobulusGrowing with Varying Water and Nutrient Supplies[J].Tree Physiology,2009,29(6):753-764.
[22]尚富华.毛白杨对修枝的形态与生理响应研究[D].北京:北京林业大学,2010.
[23]Forrester D I,Collopy J J,Beadle C L,et al.Effect of Thinning,Pruning and Nitrogen Fertiliser Application on Transpiration,Photosynthesis and Water-use Efficiency in a YoungEucalyptusNitensPlantation[J].Forest Ecology & Management,2012,266(288):286-300.
[24]Maurin V,Desrochers A.Physiological and Growth Responses to Pruning Season and Intensity ofHybridpoplar[J].Forest Ecology & Management,2013,304(4):399-406.
[25]杨玲,沈海龙,崔晓涛.NaHCO3胁迫下新西伯利亚银白杨幼苗生长和光合能力变化[J].林业科学,2012,48(7):50-55.
[26]马艳丽,朱虹,王鹏.水分胁迫对3个枣品种电导率和叶片相对含水量的影响[J].林业科技,2016,41(6):15-17.
[27]黄开勇,陈琴,戴俊,等.截干对杉木种子园老龄化母树光合特性的影响[J].基因组学与应用生物学,2016,35(6):1503-1511.
[28]刘芳,张立敏,陈斌,等.NaCl胁迫对露水草种子萌发与幼苗生长及β-蜕皮激素含量的影响[J].林业科技,2014,39(6):6-9.
[29]杨跃文,季蒙,包哈森高娃,等.干旱胁迫对不同种源北沙柳幼苗生长特性的影响[J].林业科技,2013,38(6):6-9.
[30]Turnbull T L,Adams M A,Warren CR.Increased Photosynthesis Following Partial Defoliation of Field-grownEucalyptusGlobulusSeedlings is not Caused by Increased Leaf Nitrogen[J].Tree Physiology,2007,27(10):1481-1492.
[31]Von C S,Farquhar G D.Effects of Partial Defoliation,Changes of Irradiance During Growth,Short-term Water Stress and Growth at Enhanced p(CO2)on the Photosynthetic Capacity of Leaves ofPhaseolusVulgarisL.[J].Planta,1984,160(4):320-329.
Response of Photosynthetic Physiology to Top Pruning of YoungPinusKoraiensis
Nguyen Thanh Tuan1,2,Shen Hailong1*,Wang Qinxiang1,Zhang Peng1,Vu Dinh Duy3
(1.School of Forestry,Northeast Forestry University,Harbin,150040;2.Department of Forestry,Vietnam Forestry University-Second Campus,Dongnai,810000,Vietnam;3.Vietnam National Museum of Nature,Vietnam Academy of Science and Technology,Hanoi,100803,Vietnam)
The effect of different top pruning methods(CK:no top pruning,P1:cutting down the last two layers of branches,P2:cutting down the last three layers of branches)on photosynthetic physiology in 14-year-oldPinuskoreaiensistrees was determined in this study.The result indicated that the P2 treatment significantly raised the photosynthetic physiology parameters during the first 2 months after treatment,and then such effect declined over time and eventually disappeared two months later.The photosynthetic physiological response of different age needles to top pruning had certain variation:no marked promotion effect of pruning on net photosynthetic rate,stomatal conductance and intercellular carbon dioxide concentration was detected in current year needles,except for transpiration,which was the highest in P2 treatment;P1 treatment had obvious promotion effect on net photosynthetic rate in 1-year-old needles while photosynthetic parameters were not significantly affected by P2 treatment.In 2-year-old needles,P2 showed greater effect than P1 in improving photosynthetic parameters(net photosynthetic rate,stomatal conductance,transpiration,and intercellular carbon dioxide concentration).In both one and two year-old needles,P2 treatment caused greater chlorophyll a,chlorophyll b,the total chlorophyll,and carotenoids than CK.This study revealed that the photosynthetic performance can be markedly improved by cutting down the last three layers of branches and provided a theoretical basis for further studies of rejuvenation mechanisms in the top pruning treatment to improve management quality.
Pinuskoraiensis;top pruning;photosynthetic characteristics;pigment contents
2017-04-06
中央高校基本科研业务费专项资金(2572016AA39);林业公益性行业科研专项(201204320)。
Nguyen Thanh Tuan,博士研究生。研究方向:森林定向培育、森林经理和生态学 。
*通信作者:沈海龙,博士,教授。研究方向:森林定向培育。E-mail:shenhl-cf@nefu.edu.cn
Nguyen Thanh Tuan,沈海龙,王琴香,等.截顶后红松幼树光合生理响应研究[J].森林工程,2017,33(4):1-7.
S 792
A
1001-005X(2017)04-0001-07
猜你喜欢
东北林业大学学报(2022年4期)2022-05-16
农业工程学报(2022年3期)2022-04-16
湖北农业科学(2021年13期)2021-08-04
新疆农业科技(2021年2期)2021-07-03
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
林业科学(2020年10期)2020-11-30
节水灌溉(2020年9期)2020-09-16
中学生物学(2020年12期)2020-03-30
作文周刊·小学四年级版(2019年8期)2019-04-23
