
核桃楸无性系生理生化及光合指标的遗传变异及相关性1)
2022-05-16 04:28王玮槐李志新张含国刘劲宏代伟昭石宝英
东北林业大学学报 2022年4期
王玮槐 李志新 张含国 刘劲宏 代伟昭 石宝英
(林木遗传育种国家重点实验室(东北林业大学),哈尔滨,150040) (五常市宝龙店种子林场)
核桃楸(JuglansmandshuricaMaxim.)属胡桃科(Juglandaceae)胡桃属(Juglans)落叶阔叶乔木,与水曲柳、黄波椤并称“东北三大硬阔”,被列为国家Ⅱ级珍稀树种和珍稀濒危树种的三级保护植物[1]。主要分布于我国东北地区的小兴安岭、完达山脉、长白山区和辽宁东部,华北地区也有零星分布[2-5],是东北阔叶红松林重要的伴生树种。因此,核桃楸选育及定向培育研究意义重大。核桃楸因其具有丰富的材用及果用价值,越来越多的研究者对种源及无性系生长、结实特性、种实性状进行研究[6-20],取得了较多优秀成果,为核桃楸的优良培育和价值的深度开发提供了相应的技术支撑。但是,目前关于核桃楸的研究,多数集中于用材林、种仁营养成分及生物活性的测定方面;而关于核桃楸各性状早期与晚期相关性及早期选择研究较少。
核桃楸比核桃(JuglansregiaL.)开花晚,始花树龄一般为5~8 a,开展早晚期相关性、树种各性状相关性研究,缩短育种周期十分必要。为此,本研究以1年生、5年生2个群体共11个核桃楸无性系为研究对象,以生理生化指标、光合指标为评价指标,应用变异分析、方差分析、相关性分析方法,分析各性状在嫁接当年及进入结实初期(嫁接5 a)无性系的变化规律,旨在为了解核桃楸无性系早期与晚期相关性及其优良品种选育的早期选择提供参考。
1 材料与方法
试验地:五常市宝龙店林场,位于五常市东部,属于长白山系,张广才岭西坡,低山丘陵地带,海拔197~621 m。地理坐标为东经127°38′~127°55′,北纬44°50′~44°58′。寒温带大陆性季风气候,年降水量620.9 mm,最低气温-40.9 ℃,最高气温35.6 ℃,年平均气温3.4 ℃,年无霜期120 d左右。土壤以典型暗棕壤分布最广,其次为潜育暗棕壤。
试验材料:接穗来源于黑龙江省的金山屯林业局(JST)、东京城林业局(DJC)、虎林县林业局(HL)和吉林省三岔子林业局(SC),砧木来源于五常市宝龙店林场。材料共2批,2016年在五常市宝龙店林场进行核桃楸嫁接,2017年建立无性系收集区(现5年生,已部分开花结实);2021年春季进行核桃楸嫁接,并在林场苗圃培育(1年生)。从收集区中选择与1年生相同的11个无性系进行对比分析,其中5个无性系开花结实(结实群体,SC10、SC15、SC21、DJC25、HL12)、6个无性系未开花结实(未结实群体,JST6、JST7、JST8、JST11、JST13、JST19)。为便于比较分析,也将1年生对应视为结实无性系(结实群体)、未结实无性系(未结实群体)。
结实量的调查:对结实初期(嫁接5 a)无性系,2020年结实量进行每株调查。
净光合速率的测定:2021年7月底,采用LI-6400便携式光合作用测定仪(LI-COR Inc.,USA)测定每个无性系的3株树上,健康、完整、朝南向阳方位的3片树叶的净光合速率(Pn)、气孔导度(Gs)、胞间CO2摩尔分数(Ci),蒸腾速率(Tr)。测定时间08:00—11:30,测定过程中光合有效辐射约1 800 μmol·m-2·s-1,CO2摩尔分数变化范围(400±10)μmol·mol-1。
生理生化指标的测定:2021年7月底,采集每个无性系上面3个单株健康、完整的叶片,应用BCA法蛋白含量测定试剂盒(苏州格锐思生物科技有限公司生产)测定可溶性蛋白质量分数,应用可溶性糖含量测定试剂盒(苏州格锐思生物科技有限公司生产)测定可溶性糖质量分数,使用SPAD-502便携式叶绿素测定仪测定叶绿素相对吸光值。
数据处理:数据分析主要包括变异分析、方差分析及皮尔逊、斯皮尔曼相关分析,采用Excel2016、SPSS18.0处理与分析。成活率、可溶性蛋白质量分数、可溶性糖质量分数等百分率性状经过反正弦转换后进行分析。
2 结果与分析
2.1 不同树龄核桃楸无性系生理生化及光合性状的遗传变异
遗传变异是基因型与环境交互作用的结果,遗传变异大说明可供选择范围大,从中选择优良个体进行嫁接繁殖,保持优良特性;用无性繁殖,还可缩短生殖周期,达到早开花、结实的目的。
对宝龙店核桃楸1年生11个无性系7个生理生化及光合性状进行分析(见表1),核桃楸无性系的可溶性蛋白质量分数、可溶性糖质量分数、叶绿素相对吸光值、净光合速率、气孔导度、胞间CO2摩尔分数、蒸腾速率等指标的变异幅度(平均值),分别为2.50%~7.08%(4.94%)、1.96%~4.12%(2.90%)、24.7~49.8(36.81)、1.57~18.9 μmol·m-2·s-1(10.97 μmol·m-2·s-1)、0.05~0.68 mol·m-2·s-1(0.35 mol·m-2·s-1)、235~923 μmol·mol-1(314.66 μmol·mol-1)、2.1~12.9 mmol·m-2·s-1(7.40 mmol·m-2·s-1)。各项单个指标中,气孔导度变异系数最高(为52.09%),叶绿素相对吸光值的变异系数最低(为14.70%),各项指标平均变异系数为29.31%。
对核桃楸5年生11个无性系7个生理生化及光合性状进行分析(见表1),核桃楸无性系的可溶性蛋白质量分数、可溶性糖质量分数、叶绿素相对吸光值、净光合速率、气孔导度、胞间CO2摩尔分数、蒸腾速率等指标的变异幅度(平均值),分别为3.52%~7.08%(5.72%)、1.53%~3.42%(2.20%)、26.50~52.00(41.25)、4.93~18.90 μmol·m-2·s-1(11.65 μmol·m-2·s-1)、0.14~0.81 mol·m-2·s-1(0.44 mol·m-2·s-1)、234~352 μmol·mol-1(318.81 μmol·mol-1)、4.13~13.40 mmol·m-2·s-1(9.14 mmol·m-2·s-1)。各项单个指标中,气孔导度变异系数最高(为35.83%),胞间CO2摩尔分数的变异系数最低为(6.30%),各项指标平均变异系数为21.66%。
对核桃楸11个无性系1年生、5年生7个生理生化及光合性状进行对比分析,5年生比1年生核桃楸无性系的可溶性蛋白质量分数、叶绿素相对吸光值、净光合速率、气孔导度、胞间CO2摩尔分数、蒸腾速率,分别高7.89%、6.03%、3.10%、11.43%、0.66%、11.76%;1年生比5年生核桃楸无性系的可溶性糖质量分数高15.91%。2个树龄阶段,都是气孔导度变异较大,胞间CO2摩尔分数、叶绿素相对吸光值变异相对较小。
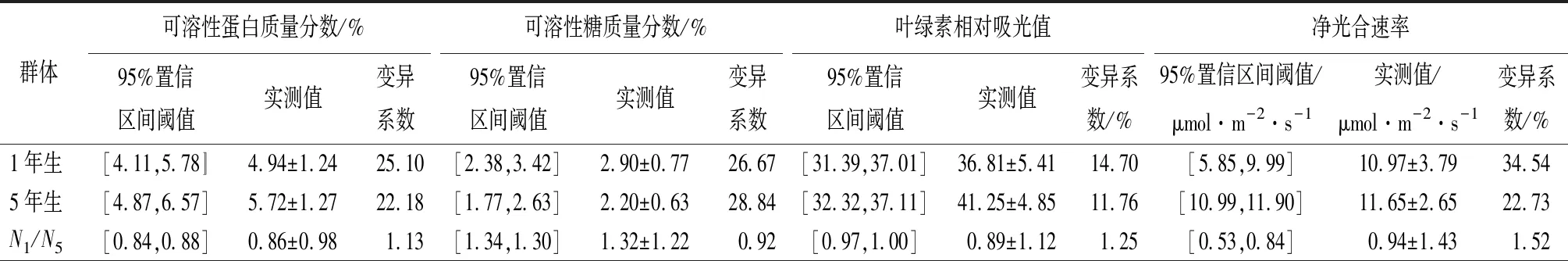
表1 1年生、5年生群体核桃楸无性系的生理生化及光合性状
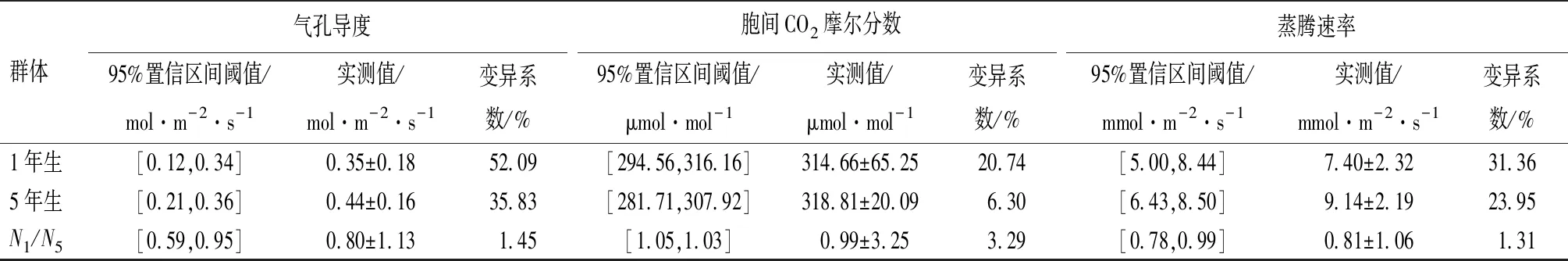
群体气孔导度95%置信区间阈值/mol·m-2·s-1实测值/mol·m-2·s-1变异系数/%胞间CO2摩尔分数95%置信区间阈值/μmol·mol-1实测值/μmol·mol-1变异系数/%蒸腾速率95%置信区间阈值/mmol·m-2·s-1实测值/mmol·m-2·s-1变异系数/%1年生[0.12,0.34]0.35±0.1852.09[294.56,316.16]314.66±65.2520.74[5.00,8.44]7.40±2.3231.365年生[0.21,0.36]0.44±0.1635.83[281.71,307.92]318.81±20.096.30[6.43,8.50]9.14±2.1923.95N1/N5[0.59,0.95]0.80±1.131.45[1.05,1.03]0.99±3.253.29[0.78,0.99]0.81±1.061.31
2.2 核桃楸无性系各性状差异显著性
2.2.1 1年生群体各性状差异显著性
1年生结实群体各性状差异显著性——对1年生结实群体进行各性状差异显著性分析(见表2)表明:可溶性蛋白质量分数、可溶性糖质量分数,在无性系间差异不显著,可溶性蛋白质量分数为2.50%(HL12)~5.97%(DJC25)、可溶性糖质量分数为1.96%(HL12)~4.12%(SC10)。叶绿素相对吸光值、净光合速率、气孔导度、胞间CO2摩尔分数、蒸腾速率,在无性系间差异显著,叶绿素相对吸光值为32.15(SC21)~37.56(HL12)、净光合速率为9.38(DJC25)~15.28 μmol·m-2·s-1(SC15)、气孔导度为0.29(SC10)~0.57 mol·m-2·s-1(SC15)、胞间CO2摩尔分数为293.67(SC10)~325.11 μmol·mol-1(SC21)、蒸腾速率为6.77(DJC25)~9.62 mmol·m-2·s-1(SC15)。HL12无性系,可溶性蛋白质量分数、可溶性糖质量分数较低,叶绿素相对吸光值较高;SC21无性系,叶绿素相对吸光值较低,而胞间CO2摩尔分数较高;SC15无性系,净光合速率、气孔导度、蒸腾速率较高。
1年生未结实群体各性状差异显著性——对1年生未结实群体进行各性状差异显著性分析(见表2)表明:可溶性蛋白质量分数、可溶性糖质量分数、胞间CO2摩尔分数,在无性系间差异不显著,可溶性蛋白质量分数为3.92%(JST13)~7.08%(JST6)、可溶性糖质量分数为2.15%(JST13)~3.99%(JST8)、胞间CO2摩尔分数为282(JST7)~374.78 μmol·mol-1(JST8)。叶绿素相对吸光值、净光合速率、气孔导度、蒸腾速率,在无性系间差异显著,叶绿素相对吸光值为32.15(JST6)~37.56(JST11)、净光合速率为5.99(JST7)~15.43 μmol·m-2·s-1(JST13)、气孔导度为0.11(JST7)~0.53 mol·m-2·s-1(JST19)、蒸腾速率为3.88(JST7)~9.52 mmol·m-2·s-1(JST19)。JST13无性系,可溶性蛋白质量分数、可溶性糖质量分数较低,而净光合速率较高;JST8无性系,可溶性糖质量分数、胞间CO2摩尔分数较高;JST6无性系,可溶性蛋白质量分数较高,而叶绿素相对吸光值较低;JST7无性系,胞间CO2摩尔分数、净光合速率、气孔导度、蒸腾速率较低;JST19无性系,气孔导度、蒸腾速率较高。
1年生11个无性系整体各性状差异显著性——对所有无性系进行各性状差异显著性分析(见表2)表明:11个无性系可溶性蛋白质量分数、可溶性糖质量分数、胞间CO2摩尔分数,在无性系间差异不显著,可溶性蛋白质量分数为2.50%(HL12)~7.08%(JST6)、可溶性糖质量分数为1.96%(HL12)~4.12%(SC10)、胞间CO2摩尔分数为235(JST7)~923 μmol·mol-1(JST8)。叶绿素相对吸光值、净光合速率、气孔导度、蒸腾速率,在无性系间差异显著,叶绿素相对吸光值为24.70(SC21)~49.80(JST11)、净光合速率为1.57(JST7)~18.90 μmol·m-2·s-1(JST13)、气孔导度为0.05(JST7)~0.68 mol·m-2·s-1(SC15)、蒸腾速率为2.10(JST7)~12.90 mmol·m-2·s-1(SC15)。HL12无性系,可溶性蛋白质量分数、可溶性糖质量分数较低;SC15无性系,气孔导度、蒸腾速率较高;JST7无性系,胞间CO2摩尔分数、净光合速率、气孔导度、蒸腾速率均较低。
综合对上述3个群体分析表明:①除结实群体胞间CO2摩尔分数无性系间差异显著,而整体与未结实群体不显著外,其余性状3个群体表现一致;可溶性蛋白质量分数、可溶性糖质量分数差异不显著,叶绿素相对吸光值、净光合速率、气孔导度、蒸腾速率差异显著。②对结实与未结实群体的T检验表明,可溶性蛋白质量分数、可溶性糖质量分数、胞间CO2摩尔分数,在结实群体与未结实群体间差异不显著,未结实群体均值较大;叶绿素相对吸光值,在2个群体间差异显著,未结实群体均值较大;净光合速率、气孔导度、蒸腾速率,在2个群体间差异显著,结实群体均值较大;该结论与具体无性系性状表现一致。
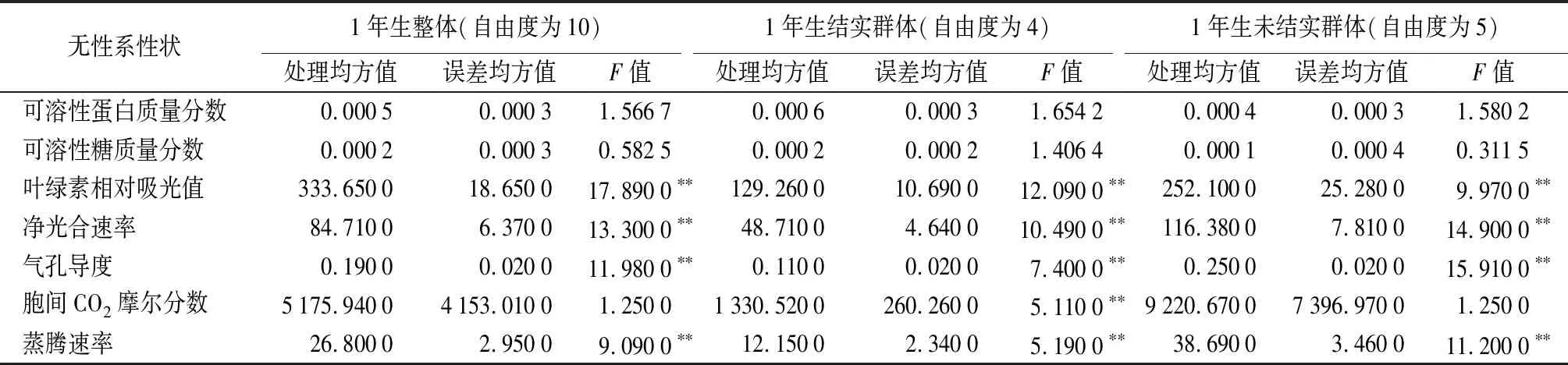
表2 1年生核桃楸无性系各性状差异显著性
2.2.2 5年生群体各性状差异显著性
5年生结实群体各性状差异显著性——对5年生结实群体进行各性状差异显著性分析(见表3)表明:可溶性蛋白质量分数、可溶性糖质量分数、胞间CO2摩尔分数、蒸腾速率,在无性系间差异不显著,可溶性蛋白质量分数为3.96%(DJC25)~7.07%(SC10)、可溶性糖质量分数为1.66%(SC15)~2.86%(DJC25)、胞间CO2摩尔分数为308.11(HL12)~325.56 μmol·mol-1(SC21)、蒸腾速率为7.52(SC10)~9.49 mmol·m-2·s-1(SC15)。叶绿素相对吸光值、净光合速率、气孔导度,在无性系间差异显著,叶绿素相对吸光值为35.92(SC10)~41.64(HL12)、净光合速率为6.49(SC21)~15.7 μmol·m-2·s-1(HL12)、气孔导度为0.28(SC21)~0.49 mol·m-2·s-1(SC10)。DJC25无性系,可溶性糖质量分数较高,而可溶性蛋白质量分数较低;SC15无性系,可溶性糖质量分数较低,蒸腾速率较高;SC10无性系,可溶性蛋白质量分数、气孔导度较高,而蒸腾速率、叶绿素相对吸光值较低;SC21无性系,净光合速率、气孔导度较低;HL12无性系,胞间CO2摩尔分数较低,而叶绿素相对吸光值、净光合速率较高。
5年生未结实群体各性状差异显著性——对5年生未结实群体进行各性状差异显著性分析(见表3)表明:可溶性蛋白质量分数、可溶性糖质量分数,在无性系间差异不显著,可溶性蛋白质量分数为2.50%(JST19)~5.97%(JST6)、可溶性糖质量分数为1.53%(JST11)~3.42%(JST6)。叶绿素相对吸光值、净光合速率、气孔导度、胞间CO2摩尔分数、蒸腾速率,在无性系间差异显著,叶绿素相对吸光值为40.03(JST6)~44.79(JST8)、净光合速率为10.21(JST19)~13.49 μmol·m-2·s-1(JST11)、气孔导度为0.25(JST6)~0.59 mol·m-2·s-1(JST13)、胞间CO2摩尔分数为291.00(JST6)~336.44 μmol·mol-1(JST8)、蒸腾速率为7.07(JST6)~12.54 mmol·m-2·s-1(JST8)。JST6无性系,可溶性蛋白质量分数、可溶性糖质量分数较高,而叶绿素相对吸光值、气孔导度、胞间CO2摩尔分数、蒸腾速率均较低;JST19无性系,可溶性蛋白质量分数、净光合速率较低;JST8无性系,叶绿素相对吸光值、胞间CO2摩尔分数、蒸腾速率较高;JST11无性系,可溶性糖质量分数较低,而净光合速率较高。
5年生11个无性系整体各性状差异显著性——对核桃楸11个无性系5年生7个生理生化及光合性状差异显著性分析(见表3)表明:可溶性蛋白质量分数、可溶性糖质量分数,在无性系间差异不显著,可溶性蛋白质量分数为3.52%(JST19)~7.08%(SC10)、可溶性糖质量分数为1.53%(JST11)~3.42%(JST6)。叶绿素相对吸光值、净光合速率、气孔导度、胞间CO2摩尔分数、蒸腾速率,在无性系间差异显著,叶绿素相对吸光值为26.50(SC10)~52.00(JST8)、净光合速率为4.93(SC21)~18.9 μmol·m-2·s-1(HL12)、气孔导度为0.14(JST6)~0.81 mol·m-2·s-1(JST13)、胞间CO2摩尔分数为234(JST6)~352 μmol·mol-1(JST8)、蒸腾速率为4.13(JST6)~13.40 mmol·m-2·s-1(JST8)。SC10无性系,可溶性蛋白质量分数较高,而叶绿素相对吸光值较低;JST6无性系,可溶性糖质量分数较高,而气孔导度、胞间CO2摩尔分数、蒸腾速率均较低;JST8无性系,叶绿素相对吸光值、胞间CO2摩尔分数、蒸腾速率较高。
综合对上述3个群体分析表明:①除结实群体的胞间CO2摩尔分数和蒸腾速率与其他群体表现不同外,其余性状3个群体表现一致。可溶性蛋白质量分数、可溶性糖质量分数差异不显著,叶绿素相对吸光值、净光合速率、气孔导度差异显著。②结实与未结实群体T检验(见表4)表明,可溶性蛋白质量分数、气孔导度、胞间CO2摩尔分数,在结实群体与未结实群体间差异不显著,未结实群体均值较大。可溶性糖质量分数、净光合速率,在2个群体间差异不显著,结实群体均值较大。叶绿素相对吸光值、蒸腾速率,在2个群体间差异显著,未结实群体均值较大。③从2个树龄性状独立样本T检验看,叶绿素相对吸光值在结实与未结实群体之间差异显著,且均为未结实群体相对较高;胞间CO2摩尔分数在2个群体之间差异不显著,且均为未结实群体相对较高;蒸腾速率在2个群体之间差异显著,1年生群体结实群体均值较大,5年生群体未结实群体均值较大。
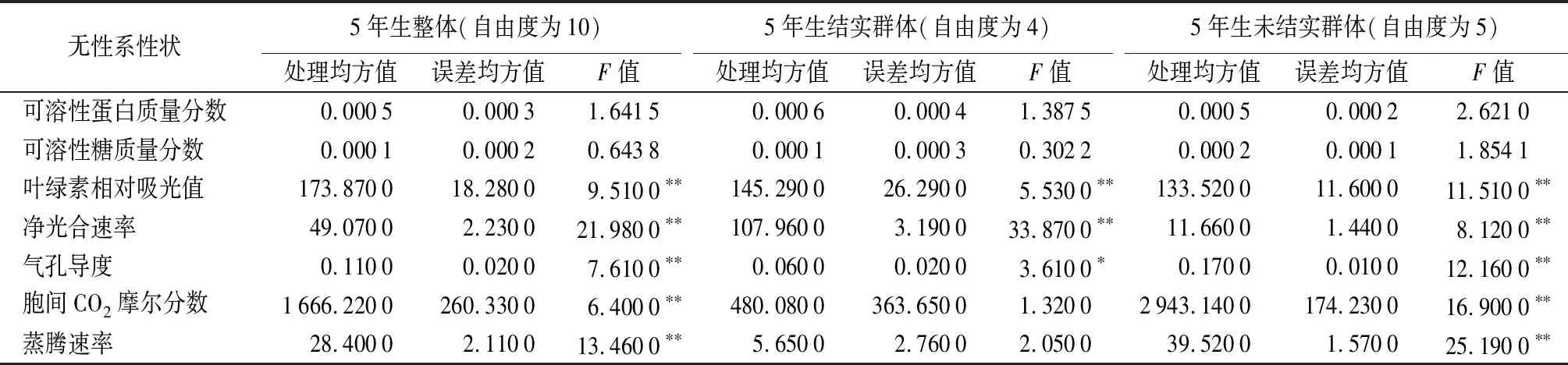
表3 5年生核桃楸无性系各性状差异显著性
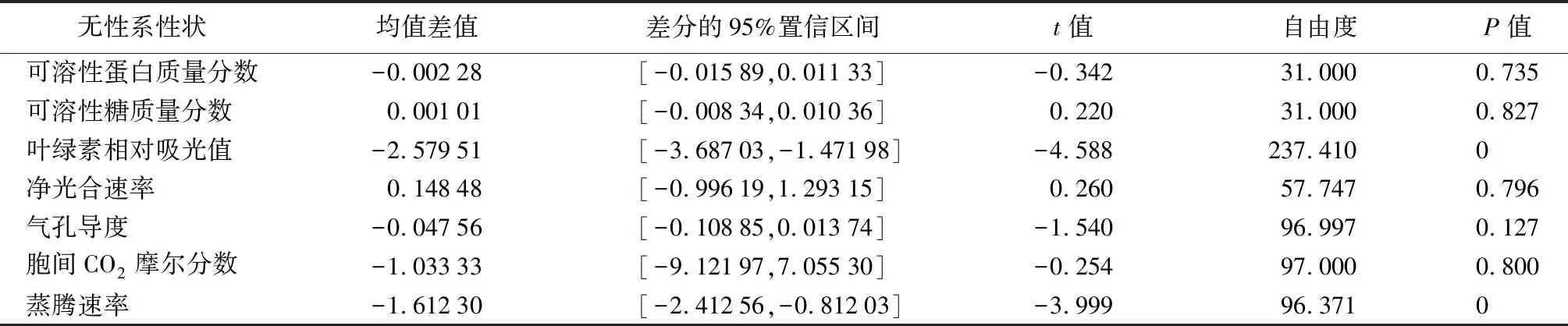
表4 5年生结实群体与未结实群体独立样本T检验
2.2.3 1年生和5年生群体之间各性状差异显著性
对未结实群体进行配对T检验发现:叶绿素相对吸光值、气孔导度、蒸腾速率,在1年生和5年生群体直降差异显著;5年生群体叶绿素相对吸光值、气孔导度、蒸腾速率相对较高。
对结实群体进行配对T检验发现:叶绿素相对吸光值在1年生和5年生群体之间差异显著,5年生群体叶绿素相对吸光值相对较高。
对整个群体进行配对T检验发现:叶绿素相对吸光值、气孔导度、蒸腾速率在1年生和5年生群体之间差异显著,5年生群体叶绿素相对吸光值、气孔导度、蒸腾速率相对较高。表明随着树龄增大,3个性状平均值均有增大趋势。
2.3 核桃楸无性系各性状之间的相关性
2.3.1 核桃楸无性系早期与晚期各性状相关性
对1年生、5年生群体各性状进行相关性分析(见表5)表明:叶绿素相对吸光值,在整体群体内各无性系之间、未结实群体内各无性系之间均呈极显著正相关,在结实群体内各无性系之间呈显著正相关,相关系数分别为0.322、0.484、0.149。叶绿素相对吸光值在早期与晚期一致性较好,有利于早期选择。胞间CO2摩尔分数,在结实群体内各无性系之间、未结实群体内各无性系之间均呈极显著正相关,在整体群体内各无性系之间呈显著正相关,相关系数分别为0.296、0.512、0.442。蒸腾速率,在结实群体内各无性系之间、未结实群体内各无性系之间均呈显著正相关。净光合速率在未结实群体内各无性系之间呈显著正相关,气孔导度在未结实群体内各无性系之间呈显著负相关。叶绿素相对吸光值、胞间CO2摩尔分数在各无性系间的相关性,经斯皮尔曼相关性分析结果表明,早期与晚期都达到了相关显著水平,相关系数分别为0.326、0.227。从2个性状绝对值排序也证明其相关性较高,1年生群体内胞间CO2摩尔分数绝对值的前三位为JST8、SC21、JST19,5年生群体内胞间CO2摩尔分数绝对值的前三位为JST8、JST13、SC21;1年生群体内叶绿素相对吸光值绝对值的前三位为JST11、JST8、JST7,5年生群体内叶绿素相对吸光值绝对值的前三位为JST8、JST11、JST19。
2.3.2 核桃楸无性系同一群体内各性状相关性
1年生群体内各性状相关性——对1年生群体各性状进行相关性分析表明:①在结实群体中,胞间CO2摩尔分数与气孔导度、与蒸腾速率呈极显著正相关;净光合速率与气孔导度、与蒸腾速率呈极显著正相关;气孔导度与蒸腾速率呈极显著正相关。②在未结实群体中,叶绿素相对吸光值与净光合速率呈极显著正相关、与气孔导度呈显著正相关;净光合速率与气孔导度、与蒸腾速率呈极显著正相关;气孔导度与蒸腾速率呈极显著正相关。③在整体群体中,叶绿素相对吸光值与净光合速率呈显著正相关、与胞间CO2摩尔分数呈显著负相关;胞间CO2摩尔分数与气孔导度、与蒸腾速率呈极显著正相关;气孔导度与净光合速率、与蒸腾速率呈极显著正相关;气孔导度与蒸腾速率呈极显著正相关。
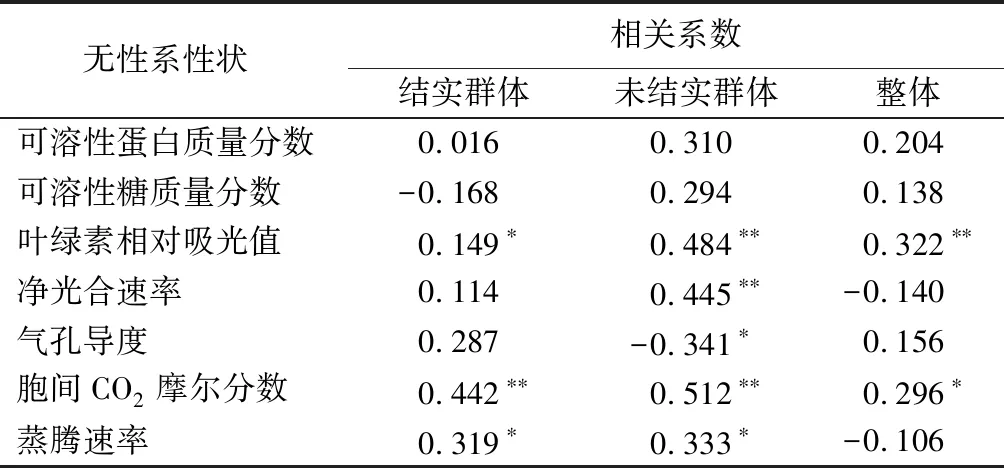
表5 3个群体核桃楸无性系各性状早期与晚期的相关性
5年生群体内各性状相关性——对5年生群体各性状进行相关分析(见表6)表明:①在结实群体中,叶绿素相对吸光值与净光合速率呈显著负相关、与胞间CO2摩尔分数呈显著正相关;胞间CO2摩尔分数,与气孔导度、蒸腾速率呈极显著正相关,与净光合速率呈显著负相关;气孔导度与净光合速率、蒸腾速率呈极显著正相关。②在未结实群体中,胞间CO2摩尔分数与气孔导度、与蒸腾速率呈极显著正相关;气孔导度与蒸腾速率呈极显著正相关。③在整体群体中,结实量与生理生化指标呈负相关、与光合指标呈正相关;叶绿素相对吸光值与结实量呈显著负相关;气孔导度与蒸腾速率呈极显著正相关、与结实量呈显著正相关;胞间CO2摩尔分数,与净光合速率呈极显著负相关,与气孔导度、与蒸腾速率呈极显著正相关。
综合对上述2个群体分析表明:在1年生、5年生所有群体中,胞间CO2摩尔分数与气孔导度、与蒸腾速率均呈极显著正相关;气孔导度与净光合速率、与蒸腾速率呈极显著正相关。除1年生结实群体、5年生整体外,其他群体的可溶性蛋白质量分数与净光合速率呈正相关。除1年生结实群体外,其他群体的可溶性蛋白质量分数与气孔导度、与胞间CO2摩尔分数、与蒸腾速率均呈正相关。除1年生结实群体外,其他群体的可溶性糖质量分数与净光合速率呈负相关。在1年生的3个群体中,叶绿素相对吸光值与净光合速率呈正相关;在5年生的3个群体中,叶绿素相对吸光值与净光合速率呈负相关。叶绿素相对吸光值与净光合速率无显著相关关系,与易干军等[21]研究结果相近。
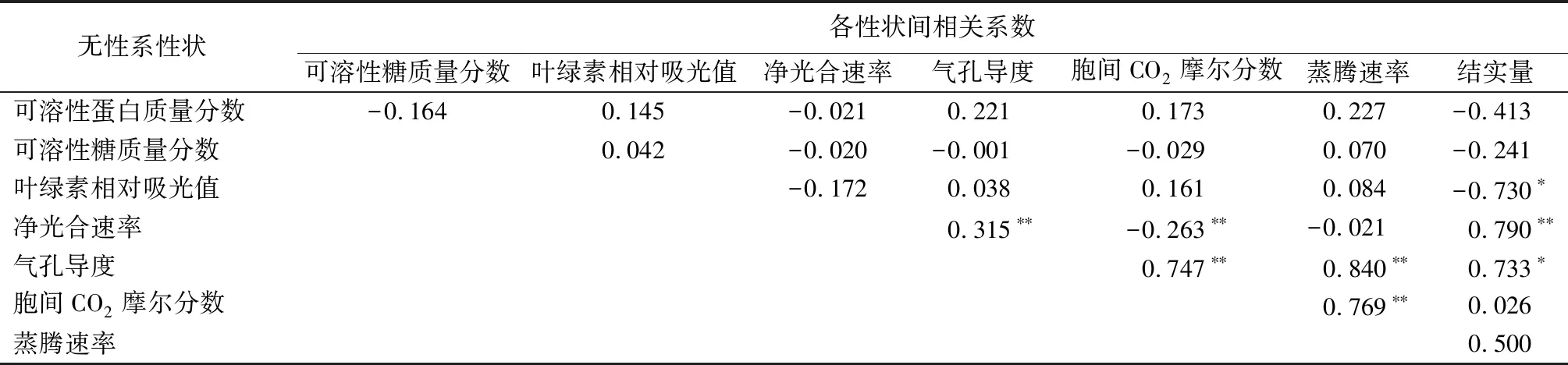
表6 5年生整体群体各性状间相关系数
3 结论与讨论
遗传变异是生物逐渐适应环境和不断进化的产物,是遗传信息的重要表征[22]。核桃楸1年生无性系性状的变异系数为14.70%~52.09%,变异最小的是叶绿素相对吸光值,最大的是气孔导度。核桃楸5年生无性系性状的变异系数为6.30%~35.83%,变异最小的是胞间CO2摩尔分数,最大的是气孔导度。5年生群体可溶性糖质量分数变异系数比1年生的高,其他6个性状都比1年生的低。
各性状差异显著性分析中,1年生群体,除结实群体的胞间CO2摩尔分数外,结实、未结实及整体群体的其余性状均表现一致,即可溶性蛋白质量分数和可溶性糖质量分数差异不显著,叶绿素相对吸光值、净光合速率、气孔导度、蒸腾速率4个性状在无性系间均差异显著。5年生群体,除胞间CO2摩尔分数、蒸腾速率外,结实、未结实及整体群体的其余性状均表现一致,即可溶性蛋白质量分数和可溶性糖质量分数差异不显著,叶绿素相对吸光值、净光合速率、气孔导度3个性状在无性系间均差异显著。1年生和5年生群体,除胞间CO2摩尔分数、气孔导度、蒸腾速率外,其余性状均表现一致,即可溶性蛋白质量分数差异不显著,未结实群体均值较大;可溶性糖质量分数差异不显著,未结实群体均值较大;叶绿素相对吸光值差异显著,未结实群体均值较大;净光合速率差异显著,结实群体均值较大。
在早期与晚期相关性分析中,叶绿素相对吸光值和胞间CO2摩尔分数在早期和晚期均变化较小,且2个树龄阶段都是胞间CO2摩尔分数和叶绿素相对吸光值变异相对较小,早期选择效果较好。高红霞等[23]对红砂家系进行叶绿素相对吸光值测定,做早期选择,筛选12个家系为抗旱性优良家系。柴文敏等[24]对唐古特白刺以叶绿素相对吸光值为标准做早期选择,筛选出5个家系作为抗旱优良家系。李森[25]认为,叶绿素相对吸光值可作为核桃品种选育的参考指标。本研究对核桃楸无性系同一群体内各性状相关性分析中,叶绿素相对吸光值早期与晚期相关显著,叶绿素相对吸光值越低,核桃楸结实量越高,因此,早期应重视叶绿素相对吸光值低的无性系的观察和培育。胞间CO2摩尔分数与净光合速率呈负相关,这与林兵等[26]的研究结果相近。性状间的相关关系影响着良种选择的效率和效果,本研究中,在1年生和5年生整体群体中,胞间CO2摩尔分数与气孔导度、与蒸腾速率均呈极显著正相关,净光合速率与气孔导度呈极显著正相关,气孔导度与蒸腾速率呈极显著正相关。
猜你喜欢
农业工程学报(2022年4期)2022-04-24
农业工程学报(2022年1期)2022-03-25
河南科技(2022年1期)2022-03-11
农业与技术(2020年12期)2020-07-04
阅读(快乐英语高年级)(2019年8期)2019-09-10
小火炬·智漫悦读(2018年12期)2018-03-13
莫愁(2017年36期)2017-12-25
江苏农业科学(2017年19期)2017-11-22
新课程·下旬(2017年7期)2017-08-14
农家顾问(2016年7期)2016-05-14
