
LC3基因沉默对小鼠肝癌细胞Heap1-6凋亡影响的研究
2017-05-13 06:02任冰霜雷艳孔旭辉王水良路君黄梁浒
中华细胞与干细胞杂志(电子版) 2017年2期
任冰霜雷艳孔旭辉王水良路君黄梁浒
LC3基因沉默对小鼠肝癌细胞Heap1-6凋亡影响的研究
任冰霜1,2雷艳1孔旭辉1,2王水良1路君1黄梁浒1
目的 建立小鼠LC3基因沉默的Heap1-6稳定表达细胞系,探讨其对衣霉素诱导的肝癌细胞凋亡的影响。方法 设计并合成一段针对小鼠LC3基因的shRNA及不针对任何基因的shRNA作为阴性对照,将它们退火后连接构建重组载体,转化扩增及酶切鉴定之后将重组质粒与病毒包装、包膜质粒共同转染293T,收集病毒上清转染Heap1-6细胞,嘌呤霉素筛选10 d,获得稳定的细胞株。以导入LC3基因shRNA的Heap1-6为实验组(Heap1-6 shLC3),以导入shcoo2的Heap1-6为对照组(Heap1-6 shctrl)。衣霉素处理实验组和对照组的细胞,Western Blot检测LC3Ⅱ、c-Caspase3、Caspase9蛋白的表达,流式细胞术检测细胞凋亡。Western Blot条带灰度值之间两组比较采用独立样本的t检验。结果 成功构建了pLKO.1-shLC3重组慢病毒载体,与野生型的Heap1-6相比,LC3基因沉默之后,LC3Ⅱ蛋白表达水平降低了62.9﹪(P < 0.01);野生型的Heap1-6和LC3基因沉默之后的Heap1-6 shLC3都经Tm处理12 h之后,后者LC3Ⅱ蛋白表达水平降低了58.6﹪(P < 0.01)。与对照组Heap1-6 shctrl相比,衣霉素作用12 h后实验组Heap1-6 shLC3 c-Caspase3增加了37.7﹪(P = 0.007),Caspase9增加了37.1﹪(P = 0.023));衣霉素作用24 h后shLC3组c-Caspase3增加了12.6﹪(P = 0.04), Caspase9增加了14.3﹪(P = 0.043)。药物干预12 h和24 h后,Heap1-6 shLC3组比对照组Heap1-6 shcoo2凋亡比例分别增加22.8﹪和18.6﹪。结论 成功建立小鼠LC3基因沉默的Heap1-6稳定表达细胞系,LC3基因沉默促进衣霉素诱导的小鼠肝癌细胞Heap1-6的凋亡。
基因; shRNA; 肝肿瘤; 细胞凋亡
自噬是真核生物体内一种高度保守的动态过程,能应对各种应激反应,例如:低氧、营养物质缺乏等发生时,自噬降解细胞内的蛋白质或者受损细胞器,并且循环利用降解产物[1-2]。然而,自噬在肿瘤发生发展的不同阶段发挥的作用不尽相同,研究表明,肝癌的发生、发展和侵袭转移都与自噬密切相关[3-5]。但是,自噬与肝癌细胞凋亡的关系尚不清楚。因此本研究拟构建慢病毒载体沉默自噬过程中的关键基因LC3(微管相关蛋白1轻链3),抑制自噬体的形成,研究抑制自噬将对肝癌细胞凋亡产生的影响,为肝癌的治疗及药物开发提供新的视角。
材料与方法
一、主要材料与试剂
人胚肾上皮细胞系293T和小鼠肝癌细胞系Heap1-6(中国科学院上海细库);DMEM/F12培养基、DMEM高糖培养基、0.25﹪胰酶-EDTA、胎牛血清(美国Hyclone公司);嘌呤霉素和pLKO.1 TRC克隆载体(美国Sigma公司);限制性内切酶AgeⅠ、EcoRⅠ(美国NEB公司);酶切鉴定所用限制性内切酶EcoRⅠ、NcoⅠ(加拿大Fermentas公司);T4 DNA连接酶(日本TaKaRa公司);快速胶回收试剂盒、质粒中抽试剂盒(德国QIAGEN公司);质粒DNA小量提取试剂盒(日本Bio Flux公司);衣霉素(tunicamycin,美国Enzo Life Sciences公司);BCA蛋白定量试剂盒、100×磷酸酶抑制剂和100×蛋白酶抑制剂(美国Pierce公司);β-actin抗体(美国Sigma公司),cleaved-Caspase3抗体、Caspase9抗体及LC3抗体(美国CST公司);辣根过氧化物酶标记的山羊抗鼠和山羊抗兔二抗(美国Thermo公司);ECL蛋白显色液(美国Advansta公司)。
二、实验方法
1.慢病毒载体的构建:设计并合成一段针对LC3的shRNA序列5'-CCGGCCTGA CCATGTCA ACATGAGTCTCGAGACTCATGTTGACATGGT CAGGTTTTTG-3',溶解稀释成20 µmol/L,95℃退火4 min备用。将pLKO.1-TRC质粒扩增后抽提质粒,应用限制性内切酶AgeⅠ、EcoRⅠ双酶切载体,产物进行DNA电泳并胶回收(回收7 kb条带)。回收DNA片段与已退火的shRNA在T4 DNA连接酶的作用下形成重组质粒。然后将重组质粒导入感受态大肠杆菌,18 ~ 24 h后挑取至少5个克隆作增菌培养,抽提质粒并进行EcoRⅠ、NcoⅠ双酶切鉴定。将鉴定正确的克隆进一步扩增培养,应用无内毒素、RNA酶和DNA酶的质粒中量抽提试剂盒抽提pLKO.1-shLC3质粒备用。不针对任何基因的阴性对照质粒shcoo2按照同样的方法进行操作。
2.细胞培养及转染:293T细胞在DF12培养基中生长密度达到40﹪~ 60﹪时,将10 μg重组质粒、2.5 μg包装质粒pDM2G、7.5 μg包膜质粒psPAX2及62.5 μg PEI同时加入到新鲜的DF12培养基中静置于37 ℃、5﹪ CO2培养箱,次日换液加5 ml DF12完全培养基,48 h、72 h分别收集病毒上清,0.4 μm滤器过滤后于-80 ℃冰箱储存。
3.Heap1-6 shLC3稳定细胞株的建立:将肝癌细胞Heap1-6培养于含10﹪胎牛血清的DMEM高糖培养基中,待增殖至50﹪~ 60﹪时加入8 ml DMEM高糖培养基、80 μl polybrene、800 μl病毒上清,18 h后弃培养液,新鲜完全培养基中加入嘌呤霉素(终浓度为1 mg/L),连续用含有嘌呤霉素的培养基筛选10 d,建立稳定传代的细胞系Heap1-6 shLC3(实验组)和Heap1-6 shctrl(对照组)。
4.Western Blot检测基因沉默后LC3的表达变化:将上述已筛选成功的Heap1-6 shLC3稳定细胞株和Heap1-6细胞株分别接种于含10﹪胎牛血清的DMEM高糖培养基中,贴壁之后用0.8 μg/ml的衣霉素处理12 h,不加药处理的做对照。待细胞生长到合适的密度时,用胰酶消化收集细胞。100 μl裂解液冰上裂解20 min,4℃ 12 000×g离心25 min,取上清作为蛋白样品,用BCA定量试剂盒对蛋白样品进行定量。取25 μg蛋白高温变性之后,进行SDS-PAGE并转至PVDF膜,4﹪BSA封闭1 h,一抗4 ℃孵育过夜。次日1×TBST洗涤3次,二抗孵育1 h,1×TBST和1×PBS各洗膜3次,加入ECL发光液暗室曝光成像,应用Bio Rad公司的Quantity One软件测得各条带的灰度值(3次结果测得平均值),用上样对照β-actin进行校正并分析实验结果。
5.Western Blot 检测LC3及凋亡相关蛋白的表达:将鉴定结果正确的实验组Heap1-6 shLC3稳定细胞株和对照组Heap1-6 shctrl分别接种于DMEM高糖培养基中,加入0.8 μg/ml衣霉素作用0、12、24 h,分别收集细胞制备蛋白样本,然后进行Western Blot,其中一抗分别为和细胞凋亡密切相关的cleaved-Caspase3抗体和Caspase9抗体,其余方法同上。
6.流式细胞术检测细胞凋亡:按上述方法,将实验组Heap1-6 shLC3细胞和对照组Heap1-6 shcoo2细胞应用衣霉素处理不同时间后,胰酶消化收集细胞,预冷PBS洗涤2次,再加300 μl PBS将细胞重悬并轻柔吹散,加入PI(终浓度为50 μg/ml),避光染色5 min,流式细胞仪上机检测并分析实验结果。
三、统计学分析方法
用SPSS17.0软件进行统计分析,Western Blot条带灰度值以±s表示,两组比较采用t 检验,以P < 0.05为差异有统计学意义。
结 果
一、pLKO.1-TRC质粒双酶切及重组质粒鉴定结果
AgeⅠ、EcoRⅠ双酶切pLKO.1-TRC载体,并经1.5﹪琼脂糖DNA电泳后可见分子量大小为7 kb和1.9 kb两条线状双链DNA分子(图1a)。胶回收7 kb条带与已退火的目的片段重组之后的连接产物用EcoRⅠ、NcoⅠ进行双酶切鉴定,可见分子量大小为5 kb和2 kb两条线状双链DNA分子(图1b),实验中挑取的6个重组载体都成功克隆了pLKO.1-shLC3载体。
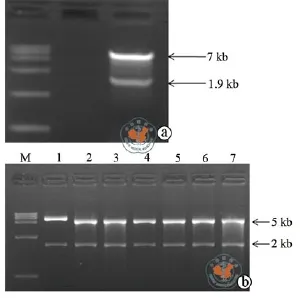
图1 空载体双酶切及重组质粒鉴定结果
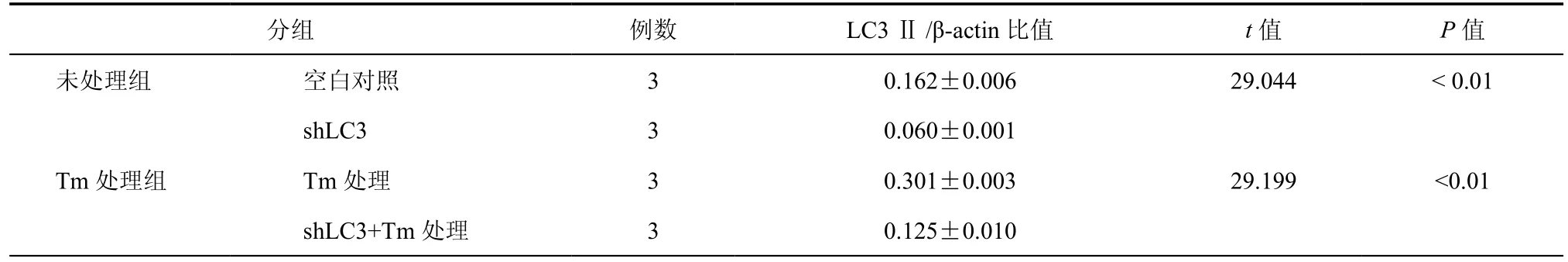
表1 Heap1-6细胞LC3基因沉默后LC3Ⅱ/β-actin结果
2.稳定转染LC3 shRNA细胞株的鉴定
在细胞自噬过程中LC3蛋白被激活,LC3前体裂解成LC3Ⅰ进而结合PE(磷脂酰乙醇胺)形成LC3Ⅱ,因此LC3Ⅱ的高低能准确反映细胞自噬水平的高低。为了鉴定LC3 shRNA 是否成功克隆到Heap1-6细胞基因组当中,应用Western Blot从蛋白水平检测LC3 shRNA对LC3表达的影响。结果显示,未进行衣霉素处理前,基因沉默后Heap1-6细胞的LC3Ⅱ/β-actin蛋白表达量比值为0.060± 0.001,与未经基因沉默的Heap1-6(0.162±0.006)相比,下降了62.9﹪(P = 0.0001,图2,表1)。衣霉素作用12 h后,LC3基因沉默后Heap1-6细胞与未经基因沉默的相比,其LC3Ⅱ/β-actin比值下降了58.6﹪(0.125±0.010和0.301±0.003,P = 0.0003,图2,表1),表明不管衣霉素刺激与否,LC3基因的shRNA序列实现基因沉默的目的,完全符合实验要求。
三、LC3 shRNA对Heap1-6细胞基因沉默后凋亡相关蛋白的影响
应用Western Blot 检测Heap1-6细胞凋亡相关蛋白cleaved-Caspase3(c-Caspase3)和Caspase9的表达改变,结果发现:与对照组(Heap1-6 shctrl)细胞相比,衣霉素作用12 h后实验组(Heap1-6 shLC3)细胞的 c-Caspase3增加了37.7﹪(P = 0.007,t = 5.027,F = 0.030),Caspase9增加了37.1﹪(P = 0.023,t = 3.589,F = 0.002);衣霉素作用24 h后,与对照组相比,实验组c-Caspase3增加了12.6﹪(P = 0.040,t = 2.990,F = 0.209),Caspase9增加了14.3﹪(P = 0.043,t = 1.766,F = 0.263)(表2)。
四、流式细胞术检测LC3 shRNA对Heap1-6细胞凋亡的影响
与未进行药物干预的对照组细胞相比,经0.8 μg/ml衣霉素刺激后,实验组细胞凋亡数量增加,且随着作用时间延长,凋亡细胞逐渐增多(图4a)。衣霉素作用12 h和24 h后,实验组细胞比对照组细胞凋亡比例分别增高22.8﹪和18.6﹪(图4b)。
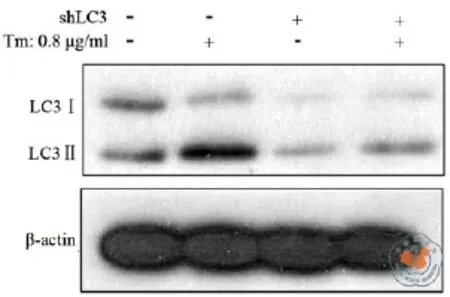
图2 LC3 shRNA对LC3表达的影响
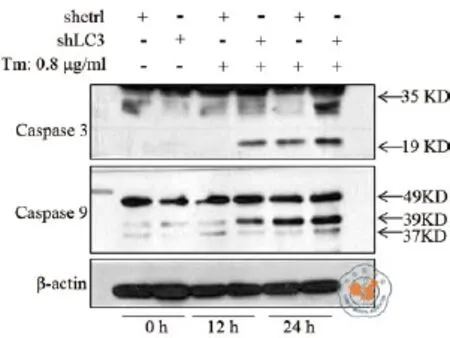
图3 LC3基因沉默对Heap1-6细胞凋亡相关蛋白的表达改变
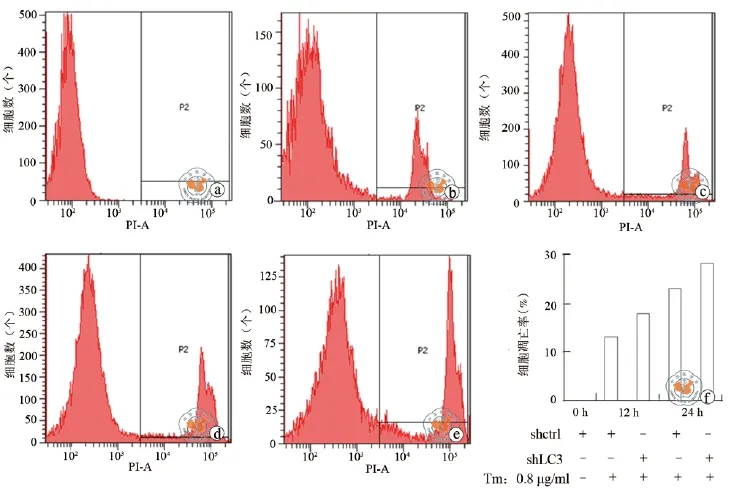
图4 LC3 shRNA对Heap1-6细胞凋亡的影响
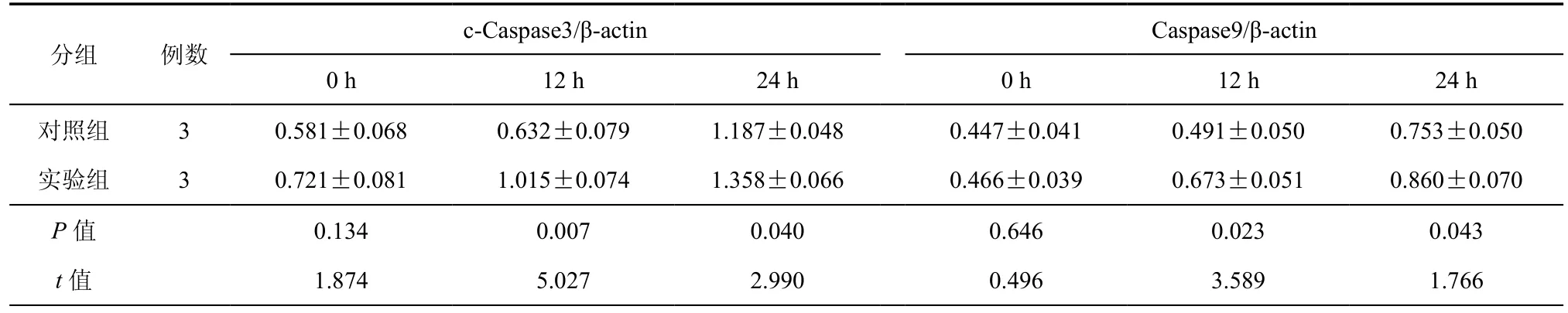
表2 基因沉默后Heap1-6细胞经衣霉素作用不同时间c-Caspase3 /β-actin与Caspase9 /β-actin结果
讨 论
肝细胞癌是全球第6大最常见的恶性肿瘤,在世界范围内居癌症死亡原因的第3位[6]。由于肝癌的隐匿性、手术低切除率和高复发率[7-9],尽管有很多分子标志与组织学特点有助于肝癌的早期诊断,然而许多促进肝癌发展演进的标志尚不十分清楚,因此更多与肝癌相关的分子机制及生物标志的发现对于预测肝癌患者的生存和预后十分重要[10]。
LC3Ⅰ在泛素样酶Atg7和Atg3的催化下,与自噬泡膜表面的磷脂酰乙醇胺结合形成LC3Ⅱ,这一过程对于自噬小体的形成起着关键的作用,可以作为自噬体的标志蛋白反映细胞自噬水平的高低。LC3的高表达与肝癌的转移呈正相关[11],肝癌原位组织中LC3表达显著低于肝转移癌中的LC3[12]。近年来多项研究发现自噬在肿瘤的发生发展中有双向调节作用[13-15]。本研究旨在弄清,LC3这一关键的自噬相关蛋白究竟对肝癌细胞的凋亡有何影响。因此采用RNA干扰技术从基因水平抑制自噬这一过程来观察其对细胞凋亡的作用。
RNA干扰(RNAi)是有效沉默或抑制目标基因表达的过程,短发卡RNA在进入细胞后被Dicer(核糖核酸酶Ⅲ家族成员之一)剪切为21个核苷酸的siRNA或miRNA而发挥干扰效应,通过诱导靶mRNA的降解实现对基因的抑制或者沉默[16-17]。本实验中,Western Blot结果表明,shRNA技术成功抑制了LC3的表达(图2a,b)。利用pLKO.1-TRC慢病毒载体把LC3 shRNA导入Heap1-6细胞,载体中的U6启动子确保shRNA总是能表达并传递到子代细胞中去,从而使LC3基因沉默可被稳定遗传,载体上的氨苄青霉素抗性基因利于质粒的筛选。
衣霉素是内质网应激经典的诱导剂,当作用时间较长引起长时间的内质网应激时会导致细胞凋亡,而内质网应激也可以介导自噬的发生[18-19]。由于自噬与细胞凋亡的关系尚存在争议,因此本实验希望通过构建针对自噬的关键蛋白LC3的慢病毒载体,阻断自噬,研究自噬对于肝癌细胞Heap1-6的影响。实验结果发现,与未处理的Heap1-6细胞相比,0.8 μg/ml的Tm处理12 h 使LC3Ⅱ水平上升了46.1﹪(P < 0.001,图2a,b)表明Tm能促进Heap1-6细胞发生自噬。LC3基因沉默之后,Western Blot结果显示自噬激活标志LC3Ⅱ表达显著降低(图2a,b),而且在Tm作用下,c-Caspase3和Caspase9表达均增高,流式分析也显示在Tm作用相同时间时,LC3基因沉默组比未处理组的细胞凋亡更多,这与Peng等[20]人的研究结果一致,表明通过抑制LC3来阻断自噬体的形成可以对肝癌细胞起到抑制作用。
本研究通过构建慢病毒载体使肝癌细胞中自噬相关蛋白LC3基因沉默,显著增加了衣霉素诱导的肝癌细胞Heap1-6凋亡,从而为肝癌的治疗及药物开发提供新的靶点。
1 吴莉, 施东婧, 李国东.自噬对大鼠原位肝移植术后急性肾损伤的影响[J].中华器官移植杂志, 2015, 36(10):606-610.
2 王绕绕, 宋红丽.自噬在肝移植缺血再灌注损伤中的研究进展[J].中华器官移植杂志, 2015, 36(9):565-568.
3 刘虹, 邵荣光.自噬在肿瘤的发生与发展过程中的调节作用[J].药学学报, 2016, 51(1):23-28.
4 沈艺南, 卢军华, 杨田.肝细胞癌相关分子机制研究新进展[J].肝胆外科杂志, 2014, 22(2):157-160.
5 朱朋飞, 秦建民.细胞自噬在肝细胞癌侵袭转移中的作用及生物学意义[J].世界华人消化杂志, 2015, 23(13):2029-2035.
6 Pan B, Yi J, Song H.MicroRNA-mediated autophagic signaling networks and cancer chemoresistance[J].Cancer Biother Radiopharm, 2013, 28(8):573-578.
7 Siegel R, Naishadham D, Jemal A.Cancer statistics.2013[J].CA Cancer J Clin, 2013, 63(1):11-30.
8 Fan ST, Mau Lo C, Poon RT, et al.Continuous improvement of survival outcomes of resection of hepatocellularcarcinoma:a 20-year experience[J].Ann Surg, 2011, 253(4):745-758.
9 Yang T, Zhang J, Lu JH, et al.A new staging system for resectable hepatocellular carcinoma:comparison with sixexisting staging systems in a large Chinese cohort[J].J Cancer Res Clin Oncol, 2011, 137(5):739-750.
10 Al-Shenawy HA.Expression of Beclin-1, an autophagy-related marker,in chronic hepatitis andhepatocellular carcinoma and its relation with apoptotic markers[J].APMIS, 2016, 124(3):229-237.
11 Wu DH, Jia CC, Chen J, et al.Autophagic LC3B overexpression correlates with malignant progression and predictsa poor prognosis in hepatocellular carcinoma[J].Tumour Biol, 2014, 35(12):12225-12233.
12 Thoen LF, Guimarães EL, Dollé L, et al.A role for autophagy during hepatic stellate cell activation[J].J Hepatol, 2011, 55(6):1353-1360.
13 Lock R, Kenific CM, Leidal AM, et al.Autophagy-dependent production of secreted factors facilitates oncogenicRAS-driven invasion[J].Cancer Discov, 2014, 4(4):466-479.
14 Shi Y, Song Q, Hu D, et al.Oleanolic acid induced autophagic cell death in hepatocellular carcinoma cellsvia PI3K/Akt/mTOR and ROS-dependent pathway[J].Korean J Physiol Pharmacol, 2016, 20(3):237-243.
15 Cerezo M, Lehraiki A, Millet A, et al.Compounds triggering ER stress exert anti-melanoma effects and overcome BRAF inhibitor resistance[J].Cancer Cell, 2016, 29(6):805-819.
16 何洁凝, 田生礼.RNA干扰的研究进展[J].生命科学研究, 2014, 18(3):265-268.
17 周天龙, 王会敏, 周青霞.RNA聚合酶Ⅱ启动子shRNA表达载体构建的研究进展[J].微循环学杂志, 2010, 20(2):47-48,50.
18 B'chir W, Maurin AC, Carraro V, et al.The eIF2α/ATF4 pathway is essential for stress-induced autophagy gene expression[J].Nucleic Acids Res, 2013, 41(16):7683-7699.
19 Shen S, Zhang Y, Wang Z, et al.Bufalin induces the interplay between apoptosis and autophagy in glioma cellsthrough endoplasmic reticulum stress[J].Int J Biol Sci, 2014, 10(2):212-224.
20 Li XF, Chen DP, Ouyang FZ, et al.Increased autophagy sustains the survival and pro-tumourigenic effects ofneutrophils in human hepatocellular carcinoma[J].J Hepatol, 2015, 62(1):131-139.
Effect of silencing LC3 on apoptosis of mouse hepatocellular carcinoma Heap1-6 cells
Ren Bingshuang1,2, Lei Yan1, Kong Xuhui1,2, Wang Shuiliang1, Lu Jun1, Huang Lianghu1.1Fujian Provincial Key Laboratory of Transplant Biology, Fuzhou General Hospital, Fuzhou 350025, China;2Medical College of Xiamen University, Xiamen 361102,China
Corresponding author: Huang Lianghu, Email: hlhtiger@163.com
Objective To establish a stable hepatocellular carcinoma Heap1-6 cell line expressing shRNA against mouse LC3 and to study apoptosis of Heap1-6 cells treated with tunicamycin.Methods shRNA targeting LC3 gene and negative shRNA were designed and synthesized.pLKO.1-TRC-shRNA LC3 vector and negative vector were constructed.After amplification and identification, the recombinant lentivirus vectors were transfected into 293T cells with packaging and envelope plasmids.The supernatant of 293T cells transfected with recombinant vector was collected, and Heap1-6 cells were transfected with plasmids, and treated with puromycin for ten days to acquire a cell line with stable expression of shRNA against mouse LC3.Western blot analysis was used to detect the expression level of LC3Ⅱ, cleaved-caspase3, and caspase9 protein respectively.Apoptotic cells were measured by flow cytometry.Results We successfully constructed pLKO.1-shLC3 lentivirus vector.Before treated by tunicamycin, the level of LC3Ⅱin the Heap1-6 shLC3 cells was decreased by 62.9﹪ compared with that in the WT Heap1-6 cells(P = 0.0001).After being treated by tunicamycin for 12 h, the level of LC3Ⅱ in Heap1-6 shLC3 cells was decreased by 58.6﹪ compared with that in the WT Heap1-6 cells P = 0.0003).Compared with the control group(Heap1-6 cells transfected with negative shRNA vector), the level of cleaved-caspase3 and caspase9 in the shLC3 group was increased by 37.7﹪ and 37.1﹪ respectively under tunicamycin for 12 h(P = 0.007, 0.023).And the level of the same two proteins in the shRNA group was elevated by 12.6﹪ and 14.3﹪ compared with those of Heap1-6 shctrl cells respectively(P = 0.040, 0.043).The ratio of apoptotic cells of the experiment group was increased by 22.8﹪and 18.6﹪compared with that of the control treated with tunicamycin for 12 h and 24 h, respectively.Conclusion LC3 knockdown could promote apoptosis of mouse hepatocellular carcinoma Heap1-6 cell line induced by tunicamycin.
Genes; shRNA; Liver neoplasms; Apoptosis
2016-12-01)
(本文编辑:陈媛媛)
10.3877/cma.j.issn.2095-1221.2017.02.007
国家自然科学基金面上项目(81270431)
350025 福州总医院泌尿外科移植生物学重点实验室1;361102 厦门大学医学院2
黄梁浒, Email:hlhtiger@163.com
任冰霜,雷艳,孔旭辉,等.LC3基因沉默对小鼠肝癌细胞Heap1-6凋亡影响的研究[J/CD].中华细胞与干细胞杂志(电子版),2017,7(2):101-106.
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
华人时刊(2022年9期)2022-09-06
成都医学院学报(2022年4期)2022-08-19
世界最新医学信息文摘(2021年12期)2021-06-09
江西农业学报(2021年4期)2021-04-20
华人时刊(2020年15期)2020-12-14
三农资讯半月报(2020年11期)2020-06-21
四川蚕业(2020年4期)2020-02-10
中国火炬(2013年11期)2013-07-25
中国火炬(2013年10期)2013-07-24
