
低血清培养PK-15细胞及其增殖猪圆环病毒2型的研究
2016-10-10 08:01张瑞永吴信明梅建国姚春阳沈志强雷连成
动物医学进展 2016年9期
张瑞永,管 宇,吴信明,梅建国,姚春阳,沈志强,雷连成
(1.吉林大学动物医学学院,吉林长春 130062;2.山东绿都生物科技有限公司,山东滨州 256600)
低血清培养PK-15细胞及其增殖猪圆环病毒2型的研究
张瑞永1,2,管宇2,吴信明2,梅建国2,姚春阳2,沈志强2,雷连成1*
(1.吉林大学动物医学学院,吉林长春 130062;2.山东绿都生物科技有限公司,山东滨州 256600)
依次采用含60、30、20、10 mL/L血清浓度的低血清培养基驯化PK-15细胞,并给驯化好的细胞上接种猪圆环病毒2型(PCV-2),以确定PCV-2在该培养体系下的生长情况。结果用含60、30、20 mL/L血清浓度的低血清培养基各进行3代次驯化,PK-15细胞能完全适应且生长状态良好;当血清浓度降至10 mL/L时,传代1次细胞无法保持良好状态,细胞出现贴壁较差、生长停滞等现象。各血清浓度培养体系中细胞生长曲线测定结果显示,在细胞培养的0、24、48、72、96 h各组细胞密度与常规培养的对照组差别不明显。因此用该低血清培养体系培养PK-15细胞时,血清最低添加量为20 mL/L。用该体系培养的PK-15细胞接种PCV-2后,通过荧光抗体染色测定病毒滴度。结果显示,低血清培养的PCV-2病毒滴度为106.5TCID50/mL,与常规条件培养的PCV-2对照(病毒滴度为106.375TCID50/mL)差别不明显,表明该体系可用于PCV-2的增殖。表明研究建立了PK-15细胞低血清培养PCV-2体系,为PCV-2的相关研究奠定了基础。
PK-15细胞;低血清培养基;猪圆环病毒2型;增殖;间接免疫荧光
随着现代生物制品行业的不断发展,对疫苗制造的安全性和有效性要求不断提升,各生产厂家对效率产能、产品质量提升的需求也越来越迫切。目前多数动物疫苗生产厂家采用PK-15细胞培养PCV-2并用来制造猪圆环病毒疫苗[1-5]。通常用含60 mL/L~100 mL/L的新生牛血清的培养基培养PK-15细胞[6-9], 但血清的使用具有价格昂贵、不同批次间存在质量差异等诸多缺点[10-14]。这些缺点使得血清在细胞大规模培养中的应用受到限制[15-19]。经过广泛研究,已经有低血清培养基产品问世,该培养基可以将血清用量降到较低水平,实现对细胞的低血清培养[20-21],使用低血清培养基具有提高疫苗质量安全性、降低生产综合成本、提高生产效率等优势,对疫苗生产企业具有重要意义。本研究从血清市场及疫苗制造的实际情况出发,经过驯化培养,建立了PK-15细胞低血清培养体系,并应用于猪圆环病毒增殖,以期为传代细胞系低血清驯化培养及圆环病毒在低血清培养细胞上的增殖研究奠定基础。
1 材料与方法
1.1材料
1.1.1细胞、毒株和试剂PK-15细胞、PCV-2 DBN株,由山东绿都生物科技有限公司保存;DMEM培养基,Gibco公司产品;T2(低血清细胞培养液),深圳壹生科公司产品;新生牛血清(BS),康源生物有限公司产品;0.8 g/L胰酶-EDTA,Amresco公司产品;PCV-2单克隆抗体,JBT公司产品;FITC标记的羊抗鼠二抗,KPL公司产品。
1.1.2仪器设备倒置荧光显微镜,CO2培养箱,吸管,洗耳球,96孔细胞培养板,细胞培养瓶,移液器等。
1.2方法
1.2.1PK-15细胞常规传代培养取生长状态良好且铺满T75瓶底的PK-15细胞,弃去培养瓶内培养液。加入5 mL胰酶-EDTA消化液洗细胞1次,更换新鲜消化液3 mL,于37℃恒温培养箱中消化3 min~5 min后,弃去消化液并吸取6 mL含100 mL/L血清的培养液轻轻吹打使之完全消化散开。按1∶3比例传至3个新的细胞培养瓶。于37℃恒温培养箱培养备用。一般经过48 h需要进行下一次传代操作。
1.2.2含60 mL/L BS低血清培养基对PK-15细胞的驯化培养将已长成良好单层的PK-15细胞按1∶3比例传至3个新的细胞培养瓶。选择其中2瓶用含60 mL/L血清的低血清培养基培养,另外1瓶按常规方法加入含60 mL/L血清的DMEM培养基培养作为对照。长满单层后各瓶均以1∶3比例继续连传3代。
1.2.3含30、20、10 mL/L BS 低血清培养基对PK-15细胞的驯化培养将用含60 mL/L BS低血清的培养基驯化的PK-15细胞按常规方法继续传代,培养基中血清降为30 mL/L、20 mL/L和10 mL/L,按1∶3比例继续连传3代。同时增设含60 mL/L血清普通培养基培养的细胞作为对照。其余浓度驯化操作步骤同上。
1.2.4PK-15细胞计数及生长曲线测定分别对正常传代PK-15细胞及用含60、30、20 mL/L BS 低血清培养基驯化的PK-15细胞进行生长曲线测定,在细胞传代后0(接种密度)、24、48、72、96 h用血球计数板按常规方法进行活细胞计数。
1.2.5驯化的PK-15细胞冻存及复苏取对生长良好的用含20 mL/L血清浓度的低血清培养的细胞,经胰酶消化后加入适量冻存液(成分为:800 mL/L 低血清培养基;100 mL/L血清;100 mL/L DMSO)。 用吸管吹打制成细胞悬液(细胞密度约为1×106~3×106/mL) 。加入1 mL细胞于冻存管中,密封后标记冷冻细胞名称和冷冻日期。细胞冻存7 d后,将冻存的PK-15细胞从液氮罐中取出,放于37℃温水中快速摇动,使其融化。在超净工作台上将其移入细胞培养瓶中,加入含20 mL/L BS的低血清培养液,37℃恒温培养。5 h~6 h后更换1次含20 mL/L BS的低血清培养液,继续置37℃恒温培养箱中培养。48 h至细胞长成单层观察细胞状态。
1.2.6PCV-2 增殖试验取低血清培养液(含20 mL/L血清)驯化并生长良好的单层PK-15细胞按1∶3比例传代,传代同时按体积比50 mL/L接入PCV-2种毒。37℃培养24 h后弃掉培养液,更换含20 mL/L血清的低血清培养基,同时添加终浓度为2 mmol/L D-氨基葡萄糖,继续培养48 h后,以2∶1比例传代,传代24 h后添加终浓度为2 mmol/L D-氨基葡萄糖,继续培养48 h后收获病毒。同时设置用含60 mL/L血清培养液培养的PK-15细胞接毒的对照。
1.2.7PCV-2 病毒滴度测定用PK-15细胞悬液将待检的病毒液在灭菌管中作1∶10系列稀释,取10-3~10-75个稀释度分别接种96孔板,每个稀释度6个孔,每孔100 μL,同时设正常细胞对照孔。37℃、体积分数为5%的CO2培养24 h,更换含2 mmol/L D-氨基葡萄糖的DMEM维持液,100 μL/孔。37℃、体积分数为5%的CO2培养48 h,待细胞长满单层。弃去维持液,用0.01 mol/L pH7.4 PBS洗涤3次,800 mL/L冷丙酮4℃固定细胞30 min,100 μL/孔,弃丙酮,用0.01 mol/L pH7.4 PBS洗涤3次,加入1∶300稀释的PCV-2抗体,50 μL/孔,37℃作用1 h;移去孔内液体,用0.01 mol/L pH7.4 PBS洗涤3次;加入1∶100稀释的FITC标记的羊抗鼠二抗,50 μL/孔,37℃作用1 h;移去孔内液体,用0.01 mol/L pH7.4 PBS洗涤3次;荧光显微镜下观察,正常细胞对照孔应无荧光物质着染,而接种病毒的细胞孔细胞胞浆内应有大量绿色荧光物质着染。计算每个稀释度含有PCV-2阳性细胞(细胞为绿色荧光物质着染)的孔数,按Reed-Muench法计算TCID50。
2 结果
2.1PK-15细胞培养结果
将含60 mL/L BS的DMEM培养基培养的PK-15细胞传代48 h后,可长至良好的单层。生长良好的细胞,边缘清晰,轮廓清楚,无悬浮的死细胞,培养液清澈,无混浊现象(图1)。

图1 含60 mL/L BS DMEM培养的PK-15 细胞单层(100×)Fig.1 PK-15 cells in 60 mL/L BS DMEM(100×)
2.2含60 mL/L BS的低血清培养基对PK-15细胞的驯化培养结果
用含60 mL/L BS的低血清培养基培养的PK-15细胞,连传3代后,细胞生长良好,边缘清晰,轮廓清楚,无悬浮的死细胞,培养液清澈,无混浊现象。与对照比较无明显差别(图2)。
2.3含30、20 mL/L BS的低血清培养基对PK-15细胞的驯化培养结果
分别用含30、20 mL/L BS的低血清培养基培养PK-15细胞,连传3代后,细胞均生长良好,边缘清晰,轮廓清楚,无悬浮死细胞,培养液清澈,无混浊现象。与对照无明显差别(图3、图4)。

图2 含60 mL/L BS T2(低血清细胞培养液)驯化培养的 PK-15细胞(100×)Fig.2 PK-15 cells in 60 mL/L BS T2(low serum cell culture medium)(100×)

图3 含30 mL/L BS T2驯化培养的PK-15细胞(100×)Fig.3 PK-15 cells in 30 mL/L BS T2(100 ×)

图4 含20 mL/L BS T2驯化培养的PK-15细胞(100×)Fig.4 PK-15 cells in 20 mL/L BS T2(100 ×)
2.4含10 mL/L BS的低血清培养基对PK-15细胞的驯化培养结果
用含10 mL/L血清浓度的低血清培养基培养PK-15细胞,仅传1代后,细胞生长出现异常,细胞出现脱落拉网现象,接近死亡(图5)。与对照差别明显。继续传代后细胞几乎不再贴壁全部死亡。
2.5细胞计数及生长曲线测定结果
分别对细胞传代后0(接种密度)、24、48、72、96 h进行活细胞计数,正常传代及含有60、30、20 mL/L BS 低血清培养基驯化的PK-15细胞测定生长曲线如图6所示。由结果可知,不同血清含量驯化的细胞在各个时期增殖情况与常规PK-15细胞无明显差别。

图5 含10 mL/L BS T2驯化培养的PK-15细胞(100×)Fig.5 PK-15 cells in 10 mL/L BS T2(100×)
2.6PCV-2滴度测定结果
在驯化好的PK-15细胞上接入PCV-2收获后测得荧光半数感染量为TCID50=10-6.5/mL,对照细胞接入PCV-2收获后测得荧光半数感染量为TCID50=10-6.375/mL。二者差别不明显,PCV-2荧光抗体检测见表1。
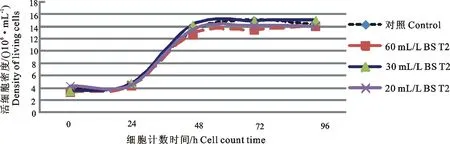
图6 不同血清浓度低血清培养基驯化的PK-15细胞生产曲线Fig.6 Proliferation curve of PK-15 cells in culture containing different concentrations of the sera
3 讨论
3.1血清浓度变化对细胞增殖的影响
低血清培养体系由于减少了血清用量,在初次传代或者降低血清含量的初代细胞会出现少量的贴壁不牢、细胞漂浮等现象[6-7],这些属于正常现象。所以,在驯化过程中需要通过连续多代次传代适应。低血清培养基培养的细胞状态会略微发生变化,表现为细胞扁平化,这可能和培养基更换以及血清含量降低有关,多次传代后可出现转变。本研究根据洪艳等[22]对BKH-21细胞低血清驯化获得的经验将血清降低次序定为60、30、20、10 mL/L,由于采用了专业配方的PK-15适应性低血清培养基,并未出现降低血清浓度后细胞生长缓慢、快速衰老死亡等变化。在降低过程中,含60、30、20 mL/L血清浓度的体系中传代3次,都表现为较快适应,镜检观察无细胞衰老、变形、细胞停滞生长等常见的细胞衰老现象。但当血清含量降至10 mL/L时,细胞出现了立即不适应现象。这说明该培养基中含有10 mL/L浓度的血清时,培养体系无法支撑细胞生长。本试验并没有将血清浓度继续细分,后续试验将继续精细优化以得到该培养基最佳的血清用量。

表1 PCV-2病毒半数感染量检测结果
通过连续的每隔24 h细胞计数,发现血清降低并未对细胞生长过程造成显著影响,计数结果均在常规误差范围内,24 h细胞均表现常规性良好的对数生长现象,48 h后由于细胞平台效应,细胞数量不再增加。所以该培养体系完全可以替代常规细胞培养体系。由于含有10 mL/L血清浓度的培养体系中细胞直接快速衰老死亡,本研究未进行该浓度下细胞计数试验。
3.2培养体系变化对病毒滴度的影响
据李智力等[23]报道,常规培养时血清浓度变化往往会对病毒增殖造成较大影响。但本试验中低血清培养系统中PCV-2接毒试验结果表明,PCV-2在该低血清培养体系中增殖并未受到显著影响。因为未来应用中低血清培养系统的目的就是将血清浓度降到较低水平,所以本试验仅在已驯化好的含20 mL/L血清的低血清培养系统进行了病毒增殖试验,数据显示,通过3次测量结果的平均值,二者数据几乎一致,与对照组并无明显差别。在培养基中添加20 mL/L血清的情况下并不影响病毒的增殖。但猪圆环病毒增殖培养本身是一个相对复杂的过程[24],简单按原始条件进行培养并不能完全代表病毒在该体系的最佳增殖状态。此外,病毒在该培养体系中的稳定性、免疫原性等关键参数是否变化仍需深入研究[25]。
总之,本试验采用低血清培养基对PK-15细胞进行驯化培养。用含20 mL/L血清的低血清培养基培养的细胞生长状态良好,在该体系中按常规方法接入PCV-2,病毒增殖滴度可达106.5TCID50/mL,与常规方法增殖病毒的滴度差异不显著。该体系可以初步应用于PCV-2的增殖培养。本研究建立的PK-15细胞及PCV-2低血清培养体系,为PCV-2的疫苗生产工艺改进及相关应用研究奠定了基础。
[1]闪伊红,王进产,郭丽霞,等.猪圆环病毒2型培养工艺研究[J].中国畜牧兽医文摘,2016,32(3):56-58.
[2]陈文庆,王建超,刘华杰,等.悬浮培养工艺与转瓶培养工艺的比较分析[J].中国兽药杂志,2010,44(10):37-41.
[3]郭玲华,王丽敏,胡修忠,等.猪圆环病毒2型培养工艺研究进展[J].中国畜牧兽医文摘,2015,31(1):37-39.
[4]杨雷,李伟,漆世华,等.微载体灌注培养Marc-145细胞制备猪繁殖与呼吸综合征疫苗[J].中国兽药杂志,2013,47(2):7-10.
[5]王家敏,马桂兰,马伟,等.昆虫细胞无血清培养基研究进展[J].动物医学进展,2016,37(2):101-104.
[6]焦文强,殷相平,柳纪省,等.哺乳动物细胞无血清培养基研究进展[J].动物医学进展,2011,32(2):69-72.
[7]张大鹤,易小萍,张元兴,等.适于重组CHO细胞培养的无血清培养基的制备[J].中国生物制品学杂志,2011,24(10):1152-1156.
[8]付百年,赵睿,王智宇,等.无血清培养基与含血清培养基培养CHO细胞的比较[J].中国生物制品学杂志,2004,17(5):158-160.
[9]Zhang Y X,Yi X P,Zhang L,et al.Animal Cell Culture Engineering[M]. Chem Indus Press,2007,40:956-960.
[10]Su L,Liu R X, Dong W J, et al.The effect of inoculum density on the growth of BHK21 cell in MEM-SLM medium[J].Vet Sci China,2007,37:1035-1037.
[11]Liu R X,Song X Y, Dong W J,et al.Study of BHK-21 cell culture with different serum level to multiply the foot-and-mouth disease virus[J].Vet Sci China(B),2007,37:1033-1034.
[12]Wang H F.Strengthen the management of raw material and auxiliary material ensure the safety of biological product[J].China Pharmac News,2008,55:785-790.
[13]Fiorin O A,Frigo G, Cucchetti E .Liquid chromatographic analysis for amino and imino acids in protein hydrolyzates by post-columnderivatization with D-phthalaldehyde and 3-mercaptopropionic acid[J].J Chromatogr,1988,476:83-92.
[14]Kioukia N,Nienow A W,Emery A N,et al.Physiological an d environmental factors affecting the growth of insect cells and infection with baculovirus[J]. J Biotechnol,1995,38(3):243-251.
[15]Tamama K,Fan V H,Griffith L G,et al.Epidermal growth fact oars a candidate for exvivo expansion of bone marrow-derived mesenchymalstem cells[J].Stem Cells,2006,24(3):686-695.
[16]Nobutaka H,Takahiro M,Mitome M,et al.Lowered glucose suppressed the proliferation and in creased the differentiation of murineneuralstem cellsinvitro[J]. FEBS Let,2004,571(1-3):237-242.
[17]Fujisaki Y,Ichihara T,Sasaki N.Field trials on inactivated porcine parvovirus vaccine for prevention of viral stillbirth among swine[J].National Institute of Animal Health Quarterly,1978,18(3):180-185.
[18]Mengeling W L,Brown T T,Paul P S,et al.Efficacy of an inactivated virus vaccine for prevention of porcine parvovirus induced reproductive failure[J]. Am J Vet Res,1979,40(7): 204-207.
[19]Wrathall A E,Wells D E,Cartwright S F,et al.An inactivated,oil-emulsion vaccine for the prevention of porcine parvovirus induced reproductive failure[J]. Res Vet Sci,1984,36(5): 136-143.
[20]Krakowka S,Ellis J A,Meehan B.Viral wasting syndrome of swine:experimental reproduction of postweaning multisystemic wasting syndrome in gnotobiotic swine by co-infection with circovirus2 and porcine parvovirus[J].Vet Pathol,2000,37(3): 254-263.
[21]Oraveeerakul K,Sooccchoi C,Molitor T W.Detection of porcine parvovirus using no radioactive nucleic acid hybridization[J].Vet Diagn Invest,1990,36(2): 85-91.
[22]洪艳,郭美锦,黄明志,等.幼小仓鼠肾细胞(BHK21)低血清驯化及代谢比较[J].化学与生物工程,2008,8:21-26.
[23]李智力,易小萍,储炬,等.PK-1细胞微载体悬浮培养生产猪细小病毒的工艺研究[J].中国生物工程杂志,2015,35(7):62-67.
[24]祝怀平,阮长耿.血管性血友病因子的研究进展[J].国外医学输血及血液学分册,2002,25(2):121-123.
[25]陈昭烈,肖成祖.动物细胞无血清培养基及其应用[J].生物工程进展,1994,14(5):23-27.
Study on PK-15 Cells Cultivated with Low Serum Culture and Culture Proliferation of Porcine Circovirus Type 2
ZHANG Rui-yong1,2, GUAN Yu2, WU Xin-ming2, MEI Jian-guo2,YAO Chun-yang2, SHEN Zhi-qiang2, LEI Lian-cheng1
(1.College of Veterinary Medicine Jilin University,Changchun,Jilin,130062;2.ShandongLvduBio-SciencesandTechnologyCo.Ltd.,Binzhou,Shandong,256600,China)
In this study,the low serum media containing 6%,3%,2% and 1% sera,respectively,were used to culture PK-15 cells and to proliferate the porcine circovirus type 2.The results showed that PK-15 cells were in good growth state in concentrations of 6%,3% and 2% sera culture medium,respectively,but when the concentration of the serum was decreased to 1%,the cells were in poor attachment and growth.Meanwhile,the densities of PK-15 cells at 0 h,24 h,48 h and 72 h time point in concentrations of 6%,3% and 2% serum culture medium was not significantly different with the control.Thereby,the culture media containing 2% serum were selected for cell cultivation and virus yield,and the virus titers were determined by indirect immunofluorescent assay.The results showed that the PCV-2 titer of the low serum culture was 106.5TCID50/mL,a little bit higher than that of the control (106.375TCID50/mL),but the difference was not significant. In this study,the PK-15 cell culture aging system with low serum culture medium for PCV-2 proliferation was established, which laid the foundation for the research of PCV-2.
PK-15 cell;low serum medium; PCV-2;proliferation;indirect immuno fluorescent assay
2016-04-11
山东省重点研发计划项目(2015GSF121027)
张瑞永(1980-),男,山东临沂人,硕士,主要从事兽用生物制品学工作。*通讯作者
S852.659.2
A
1007-5038(2016)09-0016-05
猜你喜欢
中国乳品工业(2022年4期)2022-05-19
中学生物学(2021年8期)2021-11-02
福建畜牧兽医(2021年5期)2021-10-03
生物技术通讯(2019年1期)2019-02-18
中国果业信息(2019年1期)2019-01-05
天然产物研究与开发(2018年9期)2018-10-08
中国畜牧兽医文摘(2018年6期)2018-07-28
科学与财富(2016年32期)2017-03-04
中国科技纵横(2016年20期)2016-12-28
科学启蒙(2015年8期)2015-08-07
