
生理和病理状况对小鼠肝脏SDF2L1基因表达的影响
2015-07-31 23:45王晓娟常永生
基础医学与临床 2015年12期
王晓娟,朱 斌,常永生*
(1.中国医学科学院 基础医学研究所 医学分子生物学国家重点实验室, 北京 100005; 2.首都医科大学附属北京世纪坛医院 普通外科 北京大学第九临床医学院, 北京 100038)
研究论文
生理和病理状况对小鼠肝脏SDF2L1基因表达的影响
王晓娟1,朱 斌2*,常永生1*
(1.中国医学科学院 基础医学研究所 医学分子生物学国家重点实验室, 北京 100005; 2.首都医科大学附属北京世纪坛医院 普通外科 北京大学第九临床医学院, 北京 100038)
目的 确定小鼠肝脏SDF2L1基因在不同生理和病理条件下的表达情况,并构建SDF2L表达质粒,为进一步研究SDF2L1基因的功能奠定基础。方法Real-time PCR确定SDF2L1基因在糖尿病条件下(db/db和ob/ob小鼠分别为糖尿病和肥胖模型小鼠)或不同营养状态的表达情况;设计并合成SDF2L1基因的PCR引物,以野生型C57BL/6J小鼠肝脏cDNA为模板,扩增SDF2L1编码区,PCR产物测序正确后连接到pcDNA4/myc-His表达载体,鉴定插入片段序列正确后,将构建的质粒转染293A细胞,Western blot检测SDF2L1的表达。 结果糖尿病条件下或饥饿状态下小鼠肝脏中SDF2L1基因表达明显下调(P<0.01);纯化质粒的相对分子质量为5.8 kb,酶切鉴定结果符合目的条带(666 bp)大小,插入的寡核苷酸序列与野生型SDF2L1基因序列完全相符,表达的融合蛋白大小为26 ku。结论SDF2L1基因可能参与机体糖脂代谢;成功构建SDF2L表达质粒。
质粒;SDF2L1;基因;表达
糖尿病是一种复杂的代谢性疾病,主要分为1型糖尿病(type 1 diabetes mellitus,T1DM) 和2型糖尿病(type 2 diabetes mellitus,T2DM),其中T2DM是以脂肪、骨骼肌、肝脏等胰岛靶组织胰岛素抵抗或β细胞分泌胰岛素相对不足为主要特征[1]。目前,T2DM的发病机制并不完全清楚,环境因素和遗传因素都影响T2DM的发生和发展。有研究表明内质网应激可以促进胰岛素抵抗和β细胞凋亡,同时也出现了一些基于改善内质网应激进行T2DM治疗的策略[2]。
SDF2L1蛋白的C端有一个内质网保留样模体,有研究表明SDF2L1定位于内质网,是一种新的内质网应激诱导基因[3- 4]。SDF2L1在T2DM发生发展过程中的具体作用还未有报道。本研究构建了含有SDF2L1和His标签融合的真核表达载体,旨在为今后T2DM的治疗提供线索。
1 材料与方法
1.1 材料
SDF2L1扩增序列引物(Invitrogen公司):5′-CG GGGTACCGCCACCATGTGGGGCGCGAGCCGG-3′(正向)和5′-CCGCTCGAGGAGTTCATCGTGACCCGT-3′(反向),两端分别添加KpnⅠ和XhoⅠ酶切位点。KpnⅠ和XhoⅠ酶、T4 DNA连接酶、Ex Taq DNA聚合酶、dNTPs (Takara公司)。T-easy质粒、实时定量PCR试剂盒(Promega公司)。Lipofectamine2000、细胞培养基DMEM、Trizol(Invitrogen公司)。pcDNA4/myc-His、E.coliDH5α感受态细菌以及人胚肾细胞系293A由北京协和医学院基础医学研究所实验室保存。反转录试剂盒(ABI公司)。胎牛血清、青霉素、链霉素(Hyclone公司)。6周龄的雄性C57BL/6J小鼠,体质量18~22 g [SPF级,中国人民解放军军事医学科学院实验动物中心,许可证号为SCXK-(军)2012-0004]。8周龄的雄性db/db、ob/ob 小鼠,体质量36~42 g(SPF级,南京大学模式动物研究中心)。小鼠饲养和管理按照NIH和中国医学科学院的实验动物标准执行。
1.2 小鼠禁食和禁食后重新喂食
野生型C57BL/6J小鼠分别进行正常喂食,禁食24 h,禁食24 h后重新喂食12 h,禁食48 h,禁食48 h后重新喂食12 h,禁食过程中给予正常饮水,待处理后眼眶取血,处死小鼠获肝组织,进行代谢分析测定。
1.3 总RNA提取和real-time PCR
Trizol提取小鼠肝脏细胞总RNA,紫外分光光度计定量,取3 μg 总RNA,加入随机引物进行反转录成cDNA。以cDNA为模板,在CFX- 96仪器(Bio-Rad公司)上进行扩增反应。条件为:95 ℃ 10 min、95 ℃ 20 s、58 ℃ 20 s、72 ℃ 30 s(捕捉荧光值),循环50次,并作熔解曲线。基因的定量PCR引物序列见表1。样本结果以目的基因与36B4的比值做相对定量分析,数据分析采用比较CT法,重复样本(n=3)取平均值。
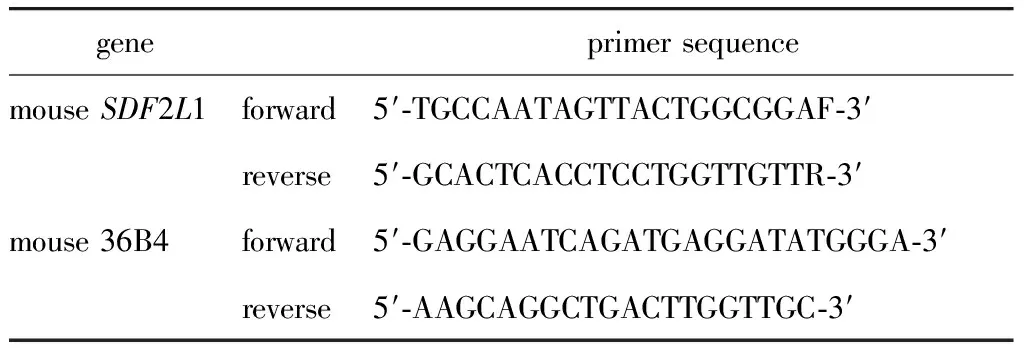
表1 实时定量PCR的引物序列
1.4SDF2L-1基因克隆与鉴定
提取C57BL/6J小鼠肝脏总RNA,反转录为cDNA。根据野生型SDF2L1基因序列设计PCR引物,分别在上下游引物中引入KpnⅠ和XhoⅠ酶切位点,以C57BL/6J小鼠肝脏cDNA为模板,进行PCR反应,切胶回收相应大小的产物片段,纯化PCR产物。
1.5 质粒连接、转化与鉴定
用限制性核酸内切酶分别消化PCR产物和质粒pcDNA4/myc-His,回收目的片段,用T4连接酶连接后转化感受态菌,抽提质粒。重组质粒经双酶切及基因测序鉴定,命名为pcDNA4/myc-His-SDF2L1。
1.6 293A细胞培养及pcDNA4/myc-His-SDF2L1转染
采用10%胎牛血清的 DMEM 培养基培养 293A细胞,培养基中添加100 U/mL青霉素和0.1 mg/mL链霉素。细胞在37 ℃、 5% CO2的孵箱中培养, 实验时取对数期细胞。实验前1 d,取对数期的293A细胞,铺6孔板,按照Invitrogen公司Lipofectamine2000转染试剂盒说明书配制转染试剂和DNA混合液,按每孔4 μL Lipofectamine2000试剂、2 μg重组质粒,混匀后加入6孔板中,置37 ℃、5% CO2孵箱中培养48 h,实验在同等条件下重复3次。
1.7 Western blot检测SDF2L1蛋白表达
蛋白样品以SDS-PAGE(分离胶浓度为10%)分离;电泳结束后将凝胶中的蛋白样品电转至PVDF膜,电转2 h,5%牛奶封闭1 h;PVDF膜置于一抗(anti-His:1∶1 000)溶液中,4 ℃摇床过夜;TBST洗膜4次,每次10 min,PVDF膜与辣根过氧化物酶标记的二抗(1∶7 500)室温作用1 h,TBST洗PVDF膜4次,每次10 min;将ECL液体以0.1 mL/cm2膜的用量滴加在PVDF膜上,室温3 min。迅速吸去多余的ECL液体,用保鲜膜包好PVDF膜,暗室曝光。
1.8 统计学分析
2 结果
2.1 SDF2L1表达水平受病理生理状态及营养状态调节
SDF2L1在db/db、ob/ob小鼠中表达下调8~9倍。SDF2L1在禁食24和48 h表达量分别下降5~6倍和7~8倍(图1)。
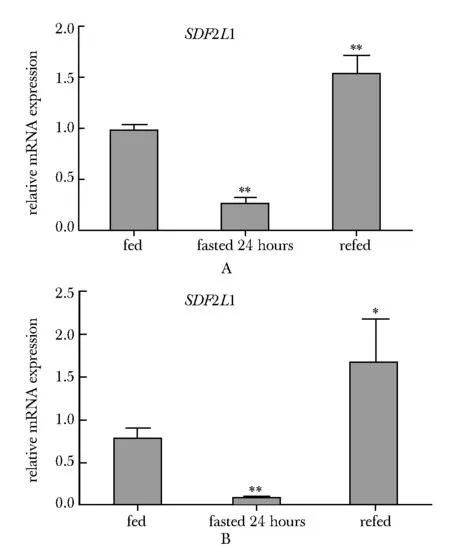
A.quantitative real-time PCR analysis showing hepatic SDF2L1 mRNA levels in eight-week-old male C57BL/6J mice under fed, 24 hours fasted and 24 hours fasted refed 12 hours conditions; B.quantitative real-time PCR analysis showing hepatic SDF2L1 mRNA levels in eight-week-old male C57BL/6J mice under fed, 48 hours fasted and 48 hours fasted refed 12 hours conditions; *P<0.05, **P<0.01 compared with fed图1 SDF2L1基因在T2DM小鼠肝脏中表达受机体营养状态调节Fig 1 Hepatic SDF2L1 gene expression is regulated by nutritional status
2.2SDF2L1基因克隆
PCR所得片段与预期一致(图2),所获得的序列与UCSC中注册的SDF2L1序列相同。
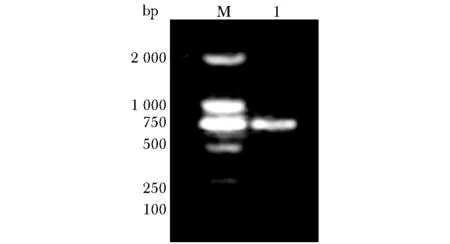
M.DL2000 DNA marker; 1.PCR products of SDF2L1图2 小鼠肝脏cDNA中SDF2L1基因PCR扩增Fig 2 PCR amplification of SDF2L1 gene from mouse liver cDNA
2.3 重组载体酶切鉴定结果
限制性内切酶双酶切鉴定重组质粒pcDNA4/myc-His-SDF2L1,进行琼脂糖凝胶电泳,结果进一步显示基因插入正确(图3)。
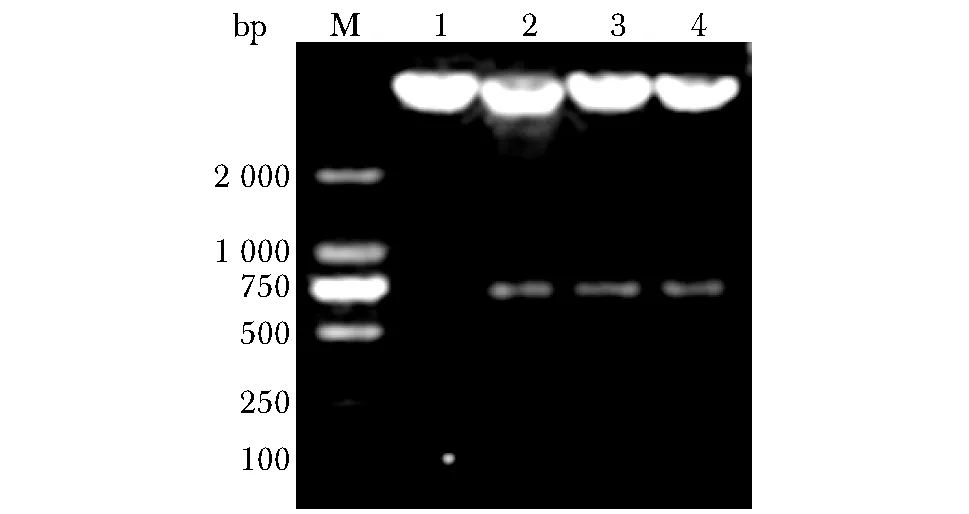
M.DL2000 DNA marker; 1.Kpn Ⅰand Xho Ⅰdigestion of pcDNA4/myc-His; 2~4.Kpn Ⅰand Xho Ⅰdigestion of positive clone with pcDNA4/myc-His-SDF2L1图3 pcDNA4/myc-His-SDF2L1重组子双酶切鉴定Fig 3 Verification of positive clone with pcDNA4/myc- His-SDF2L1 by restriction endonuclease (RE) digestion
2.4 重组质粒转染293A细胞后SDF2L1的表达
与对照组(con、空载体pcDNA4)相比, pcDNA4/myc-His-SDF2L1质粒转染的细胞中有一个26 ku特异性条带,而在对照组则没有条带(图4)。
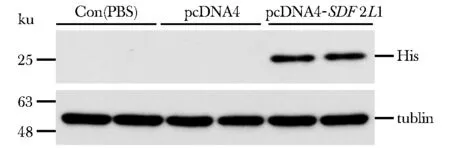
图4 SDF2L1质粒构建验证Fig 4 The identification of SDF2L1 plasmid
3 讨论
肝脏的主要功能是调控葡萄糖和其他新陈代谢原料的供应, 血糖水平的维持依赖葡萄糖的产生和
周边组织神曲葡萄糖之间的平衡。葡萄糖的产生主要通过肝脏,其次是肾脏,并受胰岛素信号通路的密切调控。
内质网是蛋白质生物合成的重要场所,新合成的蛋白质转移到内质网上,与相关的伴侣蛋白结合帮助其进行翻译后修饰,如糖基化修饰以及二硫键的形成[5],正确折叠的蛋白质输送到高尔基体,进一步对其加工、储存和分泌等,而部分错误折叠的蛋白质,则要经过内质网介导降解系统(endoplasmic reticulum associated degradation, ERAD)或转移出内质网进行泛素化降解[6]。未折叠或错误折叠蛋白质在内质网的聚集可以引发内质网应激,近年来的研究显示,内质网应激与糖尿病关系密切[7]。
基质细胞衍生因子(stromal cell derived factors, SDFs)家族是一类由基质细胞产生的一组分子[8]。SDF1和SDF4是化学趋化因子;SDF2和SDF5的结构和功能均未确定。在人类和小鼠,SDF2L1是SDF2的同源基因,它们的蛋白序列有78%相似,其中68%完全相同[9]。有研究分析了SDF2L1基因结构,小鼠SDF2L1基因组DNA包含3个外显子,分子大小为2.2 kb;人类SDF2L1基因组DNA也包含3个外显子,分子大小为2.0 kb[4]。小鼠和人的核苷酸序列位于染色体相同的位置[10]。最新研究表明,SDF2L1在胰腺β细胞中错误折叠胰岛素原的降解过程中起重要作用[3],提示SDF2L1可能在糖尿病的发生发展过程中起重要功能。
本研究成功构建了带有His标签的SDF2L1真核表达质粒,为进一步研究SDF2L1蛋白的功能及研制预防或治疗糖尿病的药物奠定基础。
[1] Kahn SE, Hull RL, Utzschneider KM. Mechanisms linking obesity to insulin resistance and type 2 diabetes[J]. Nature, 2006, 444:840- 846.
[2] Ozcan U, Cao Q, Yilmaz E,etal. Endoplasmic reticulum stress links obesity, insulin action, and typ2 diabetes[J]. Science, 2004, 306: 457- 461.
[3] Akansha T, Irmgard S, Liling Z,etal. SDF2L1 interacts with the ER-associated degradation machinery and retards the degradation of mutant proinsulin in pancreatic β-cells[J]. J Cell Sci, 2013, 126:1962- 1968.
[4] Saburo F, Masaharu S, Yuji M,etal. Murine and human SDF2L1 is an endoplasmic reticulum stress-inducible gene and encodes a new member of the Pmt/rt protein family[J]. Biochem Biophys Res Commun, 2001, 280:407- 414.
[5] Braakman I, Bulleid NJ. Protein folding and modification in the mammalian endoplasmic reticulum[J]. Annu Rev Biochem, 2011, 80:71- 99.
[6] Vembaf SS, Brodsky JL. One step at a time: endoplasmic reticulum associated degradation[J]. Nat Rev Mol Cell Biol, 2008, 9:944- 957.
[7] Crunkhorn S. Type 2 diabetes: ER stress modulator reverses diabetes[J].Nat Rev Dru Discov, 2015, 14: 528- 536.
[8] Kang H, Esxudero-Esparza A, Douglas-Jones A,etal. Transcript analyses of stromal cell derived fastors9 (SDFs): SDF- 2,SDF- 4 and SDF- 5 reveal a different pattern of expression and prognostic association in human breast cancer[J]. Int J Oncol, 2009, 35:205- 211.
[9] Lorenzon-Ojea AR, Caldeira W, Ribeiro AF,etal. Stromal cell derived facor (sdf2): A novel protein expressed in mouse[J]. Int J Biochem Cell Biol, 2014, 53:262- 270.
[10] Dunham I, Shimizu N, Roe BA,etal. The DNA sequence of human chromosome 22[J]. Nature, 1999, 402:489- 495.
新闻点击
婚姻状态影响女性心脏病死亡风险
据美国国家科学院院报(PNAS)网站(2014- 03- 27)报道,婚姻状态与心脏病死亡的风险有关联,过去曾针对这样的问题在男性身上进行过研究,结果发现已婚男性比未婚男性的心脏病死亡风险降低。日前英国剑桥大学的研究团队想了解这样的结论是否在女性身上也适用,进行一项研究,最后证明结果相同。
研究资料来自英国百万妇女研究中超过130万名50岁以上女性的数据,这些女性在1996—2001年加入该研究,研究团队再从中挑选734 626名女性因心脏病入院与死亡的数据进行分析。
分析结果显示,有伴侣的女性发展心脏病的风险与已婚女性相同,但是死于心脏病的风险却减少28%。研究表示,风险较低的原因可能是女性因为受到伴侣的鼓励与支持,会比较愿意寻求治疗,向医生反应疾病症状,但对医嘱的顺从性却与未婚女性没有差别。
该研究刊登于《生物医学中心期刊》(BMCMedicine)。
女性出生体质量过轻长大容易不孕
据美国国家科学院院报(PNAS)网站(2014- 04- 02)报道,随着医疗进步,越来越多出生体质量过轻的儿童能幸存下来并健康长大成人,但最近一项瑞典的研究指出,这可能也会代表不孕慢慢增加,研究发现,这些出生体质量过轻的女孩长大后发生不孕的概率将是一般女孩的2倍。
研究分析1 293位女性的资料,这些女性都是在1973年后出生于瑞典,并在2005—2010年接受不孕治疗的患者。资料分析包括这些女性的不孕原因、她们出生时的大小、参与研究时的年龄与体质指数。根据数据显示,不孕原因复杂,女方不孕占38.5%、男方占27%、双方都不孕占7%,其余为不明原因。出生体质量方面,约有10%的女性出生体质量过轻或体型过小。
但研究样本数太少、且仅针对瑞典所做,可能无法适用于其他国家,未来应针对同样的问题有更多研究去证实两者间确切的关联性。
该研究刊登于新一期BMJOPEN期刊。
Effects of different physiological and pathological conditions on mouse hepaticSDF2L1 gene expression
WANG Xiao-juan1, ZHU Bin2*, CHANG Yong-sheng1*
(1.National Key Laboratory of Medical Molecular Biology, Institute of Basic Medical Sciences, CAMS, Beijing 100005; 2.Dept. of General Surgery, Beijing Shijitan Hospital, Capital Medical University, Peking University Ninth School of Clinical Medicine,Beijing 100038, China)
Objective To determine the expression ofSDF2L1 in mouse liver under different physiological and pathophysiological conditions and to performSDF2L1 overexpression by using eukaryotic expression system for further function research. Methods The expression ofSDF2L1 in mouse liver under different conditions were determined by Real-time PCR. Primers were designed commercially. Total RNA was isolated from the wild type C57BL/6J mouse liver and reverse transcribed to cDNA. The coding sequence ofSDF2L1 was amplified by PCR. The product of PCR was purified and inserted into pcDNA4/myc-His vector and then transformed intoE.colicompetent cells. Expression of recombinant was transfected into 293A cells and identified by SDS-PAGE. Results The fasting or pathological condition lead to decreased expression levels ofSDF2L1. The molecular weight of purified plasmid was 5.8 kb. The product was 666 bp and its sequence was identical to that in wide typeSDF2L1. The recombinant protein was 26 ku.ConclusionsSDF2L1 may play a significant role in the regulation of hepatic glucose and lipid metabolism.
plasmid;SDF2L1; gene; expression
2015- 09- 22
2015- 10- 26
国家自然科学基金(81471049)
1001-6325(2015)12-1601-05
Q591
A
*通信作者(corresponding author):binbinzhu99@sohu.com; changy@ibms.pumc.edu.cn
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
解放军医学杂志(2021年12期)2022-01-18
现代临床医学(2021年1期)2021-01-26
食品科学(2018年10期)2018-05-23
罕少疾病杂志(2017年2期)2017-02-23
安徽医科大学学报(2016年12期)2017-01-15
广西林业科学(2016年3期)2016-03-16
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年33期)2015-03-01
