
HBX蛋白反式激活基因XTP4抑制HepG2细胞凋亡
2015-07-31 23:45:34时朝辉张梦然王丽丽刘顺爱梁跃东
基础医学与临床 2015年12期
时朝辉,张梦然,王丽丽,刘顺爱,梁 璞,吴 君,成 军*,梁跃东*
(1.贵州医科大学, 贵州 贵阳 550004; 2.首都医科大学附属北京地坛医院 传染病研究所, 北京 100015)
研究论文
HBX蛋白反式激活基因XTP4抑制HepG2细胞凋亡
时朝辉1,张梦然2,王丽丽1,刘顺爱2,梁 璞2,吴 君1,成 军2*,梁跃东1*
(1.贵州医科大学, 贵州 贵阳 550004; 2.首都医科大学附属北京地坛医院 传染病研究所, 北京 100015)
目的探讨乙型肝炎病毒X蛋白(HBXAg)反式激活基因4(XTP4)对肝母细胞瘤细胞HepG2细胞凋亡的影响。方法构建XTP4基因的真核表达质粒pcDNA3.1/myc-His(-)A-XTP4(pXTP4)及化学合成XTP4基因的干扰RNA(siXTP4)后,分别将XTP4基因的真核表达质粒、干扰RNA及各自的阴性对照(NC、siNC)瞬时转染HepG2细胞中,48 h后进行以下实验:用Western blot验证XTP4在蛋白水平的过表达和干扰表达;流式细胞术检测细胞凋亡;Western blot 检测Bcl- 2、Bax蛋白水平并计算Bcl- 2/Bax比值;化学发光法检测胱冬肽酶- 3(caspase- 3)的活性。结果成功构建了XTP4的真核表达质粒pXTP4、XTP4在蛋白水平过表达和干扰表达。与各自的阴性对照组相比,过表达XTP4的HepG2细胞中,Annexin V+细胞数显著减少(P<0.05),Bcl- 2/Bax比值增高(P<0.05),胱冬肽酶- 3活性下降(P<0.05);而干扰XTP4表达的HepG2细胞中,Annexin V+细胞数显著增加(P<0.05),Bcl- 2/Bax比值降低(P<0.05),胱冬肽酶- 3活性上升(P<0.05)。 结论XTP4具有抑制HepG2细胞凋亡的作用,可能是通过增加Bcl- 2/Bax的比值实现。
XTP4基因;HepG2细胞;细胞凋亡
乙型肝炎病毒(hepatitis B virus,HBV)感染呈世界性流行,全球有3.5亿人为慢性HBV感染者,每年约有100万人死于HBV感染所致疾病[1],据2006年全国乙肝流行病学调查表明,中国1~59岁一般人群乙型肝炎病毒表面抗原(HBsAg)携带率为7.18%[2]。乙型肝炎病毒X蛋白(HBxAg)是由HBV最小的开放读码框X基因编码的病毒蛋白,因其与所有的已知蛋白没有同源性,故命名为X蛋白,X基因片段是HBV最主要整合片段,X蛋白也是恶性转化肝细胞中主要表达的病毒蛋白[3]。研究表明,HBxAg是一种多功能的调节蛋白,具有广泛的反式激活功能,其通过反式激活作用,调控肝细胞某些基因的表达,影响细胞凋亡、分化、增殖等生物学功能,在HBV致病、致癌过程中发挥重要作用,但其具体机制尚不完全明了[4]。本课题组于2004年应用抑制性消减杂交技术筛选并克隆了HBXAg反式激活靶基因4(XTP4)[5],本研究旨在探讨XTP4对HepG2细胞凋亡的影响,以明确XTP4是否通过对细胞凋亡的影响参与HBV相关肝病的发病与进展。
1 材料与方法
1.1 实验材料
1.1.1 细胞、质粒和siRNA:HepG2细胞、pcDNA3.1/myc-His(-)A质粒(均本实验室保存);siRNA(上海吉玛制药技术有限公司合成),阴性对照(siNC)为吉玛公司提供的与目的序列无同源性的通用阴性对照。序列如下:siNC:sense 5′-UUC UCCGAACGUGUCACGUTT- 3′,antisense 5′-ACGUG ACACGUUCGGAGAATT- 3′;siXTP4:sense 5′-UCU GGGUUCCUGCUCUGUUTT- 3′,antisense 5′-AACAG AGCAGGAACCCAGATT- 3′。
1.1.2 试剂:DMEM细胞培养基和胎牛血清(Gibco公司);jet PRIMETM DNA&siRNA转染试剂(Polyplus公司);细胞凋亡检测试剂AnnexinV-FITC和7-AAD(Biolegend公司);胱冬肽酶- 3发光检测试剂盒(Promega公司)。One Step RNA PCR试剂盒和pGEM-Teasy载体等[宝生物工程(大连)有限公司]。兔抗人XTP4抗体(Abcame公司);抗-myc标签、抗-Bcl- 2、抗-Bax、抗-GAPDH、山羊抗兔IgG-HRP和山羊抗鼠IgG-HRP(Santa公司)。
1.2 方法
1.2.1 真核表达质粒的构建:提取HepG2细胞中的RNA,反转录为cDNA,以此为模板PCR扩增XTP4基因全长片段;PCR引物根据 NCBI中提交的 XTP4 的基因序列设计,进行PCR反应后琼脂糖凝胶电泳鉴定扩增结果并进行切胶纯化回收。将回收产物连接TA载体进行序列测定,经XhoⅠ和EcoRⅤ双酶切后将目的基因连接到真核表达载体pcDNA3.1/myc-His(-)A中,构建的表达载体进行酶切和测序鉴定。
1.2.2 细胞培养及重组质粒转染:用含10%胎牛血清的DMEM培养液,于5% CO2孵箱中培养HepG2细胞;待细胞铺满培养瓶底后,将细胞传代种植于6孔板中,待细胞达60%~70%汇合度时进行重组质粒和干扰RNA的转染,并设立各自的阴性对照。
1.2.3 Western blot验证XTP4蛋白的过表达和干扰表达:质粒转染48 h后提取细胞总蛋白,进行 Western blot,一抗为抗-myc标签、兔抗人XTP4抗体、抗-GAPDH,二抗为山羊抗鼠IgG-HRP、山羊抗兔IgG-HRP。
1.2.4 Annexin V检测细胞凋亡:转染48 h后收集细胞,按照Annexin V/7-AAD检测试剂说明书处理细胞后利用流式细胞仪检测细胞凋亡。Annexin V-7-AAD-为正常细胞;Annexin V+7-AAD-为早期凋亡细胞;Annexin V+7-AAD+为中晚期凋亡细胞。
1.2.5 Western blot检测Bcl- 2、Bax蛋白表达:转染48 h后提取细胞总蛋白,按照常规方法进行Western blot检测。一抗为抗-Bcl- 2、抗-Bax、抗-GAPDH,二抗为山羊抗兔IgG-HRP、山羊抗鼠IgG-HRP。应用 IMAGEPROPLUS 软件进行灰度分析。
1.2.6 胱冬肽酶- 3(caspase- 3)的活性检测:将HepG2细胞以每孔1×104个细胞接种于96孔板中,转染后48 h,按照胱冬肽酶- 3化学发光试剂盒的操作流程处理细胞后检测细胞内胱冬肽酶- 3的活性。
1.3 统计学分析

2 结果
2.1 成功构建XTP4的真核表达载体pcDNA3.1/myc-His(-)A-XTP4
用RT-PCR扩增出XTP4,扩增片段约348 bp,与预期符合(图 1A)。酶切鉴定可见5 500 bp大小的pcDNA3.1/myc-His(-)A线性片段以及348 bp的XTP4基因片段(图 1B)。经测序,序列比对完全一致,表明XTP4基因的真核表达载体构建成功。

A:1.DNA marker 2000, 2.RT-PCR product; B:1.DNA marker 2000, 2.pXTP4 enzyme products图1 XTP4真核表达载体的构建Fig 1 The construction of eukaryotic expression vector of XTP4
2.2 Western blot验证XTP4在蛋白水平的过表达和干扰表达
转染48 h后 Western blot,抗-myc标签、抗-XTP4、抗-GAPDH,验证了 XTP4蛋白的过表达和干扰表达,XTP4蛋白相对分子质量约为12 ku, pcDNA3.1/myc-His(-)A-XTP4(pXTP4)因有myc、His标签,蛋白相对分子质量约为17 ku,与预期相符(图2)。

NC.normal controls;pXTP4.pcDNA3.1/myc-His(-)A-XTP4;siNC.siRNA normal controls;siXTP4.XTP4 small interference RNA图2 Western blot验证XTP4过表达和干扰表达Fig 2 Western blot verified the overexpression and interference expression of XTP4
2.3 XTP4对Annexin V+细胞数的影响
与各自的对照组相比,过表达XTP4后Annexin V+细胞数显著减少,总凋亡率显著减少(P<0.01);干扰XTP4表达后Annexin V+细胞数显著增加,总凋亡率显著增加(P<0.01)(图3)。
2.4 XTP4对Bcl- 2/Bax 比值的影响
与各自的对照组相比,过表达XTP4 组Bcl- 2/Bax的比值升高(P<0.01),干扰XTP4表达组Bcl- 2/Bax的比值降低(P<0.01)(图4)。
2.5 XTP4对胱冬肽酶- 3活性的影响
与各自的对照组相比,过表达XTP4组胱冬肽酶- 3的活性下降(P<0.05),干扰XTP4组胱冬肽酶- 3的活性增加(P<0.05)(图5)。
3 讨论
XTP4最早由美国国立卫生研究院哺乳动物基因集合(MGC)计划通过代表性差异分析技术于2002年发现,并命名为C17orf37/MGC14832[6],在Gene bank 中的别名还有C35、M1EN1、RDX12 和ORB3等。 XTP4蛋白质编码区全长348 bp,位于人类染色体17q12-17q21,肿瘤研究的热点HER2扩增子内,由4个外显子,3个内含子组成。目前已有研究发现XTP4在多种恶性肿瘤中高表达(乳腺癌、前列腺癌、卵巢癌),与乳腺癌的分级、分期明显相关,具有促进肿瘤细胞迁移和侵袭的作用,而且与肿瘤的耐药相关[7- 11]。
本课题组前期应用基因芯片技术和抑制性消减杂交技术筛选了XTP4反式调节基因和差异表达基因[12- 13],提示XTP4可能参与细胞凋亡、细胞增殖与分化、肝细胞纤维化、 肿瘤发生等多种生物学过程,提示XTP4可能在HBV相关肝病的发生中起着重要作用,其机制研究对于进一步了解HBV相关肝病的发病机制具有重要意义。
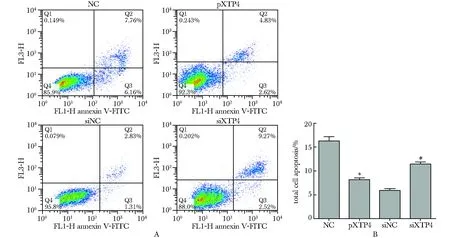
*P<0.01 compared with NC; #P<0.01 compared with siNC
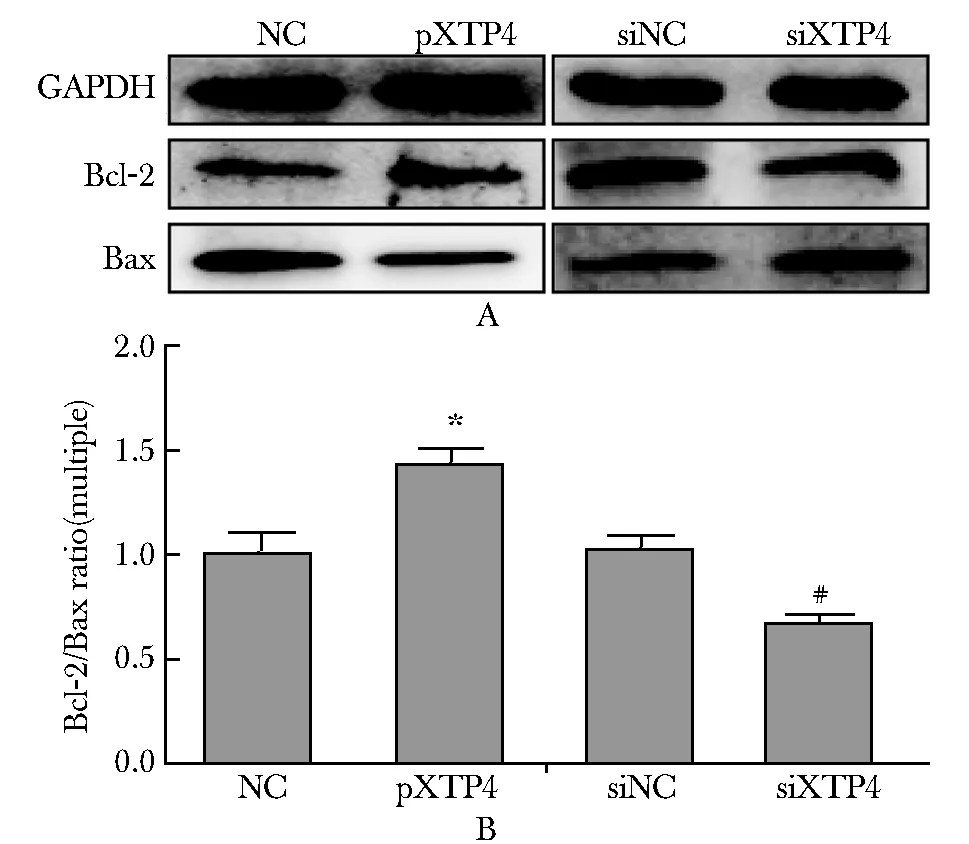
A.Western blot detected the expression of Bcl- 2, Bax protein;B.the ratio of Bcl- 2 to Bax results statistical figure; *P<0.01 compared with MC; #P<0.01 compared with siNC图4 XTP4对Bcl- 2/Bax比值的影响Fig 4 The influence of XTP4 on the ratio of Bcl- 2 to Bax(n=6)
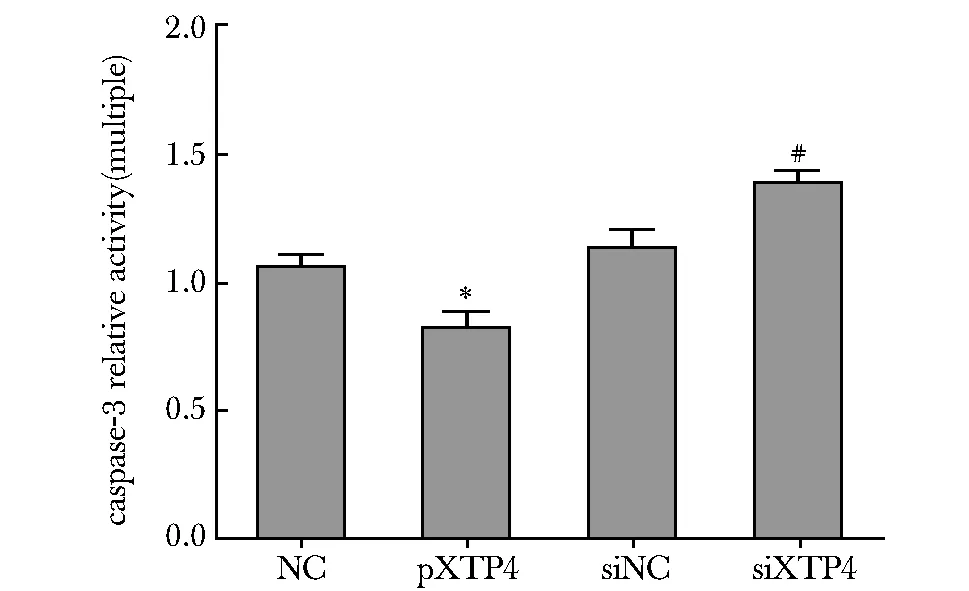
*P<0.01 compared with NC; #P<0.01 compared with siNC图5 XTP4对胱冬肽酶- 3活性的影响Fig 5 The influence of XTP4 on the activities of caspase- 3(n=6)
本研究Annexin V/7AAD结果显示,过表达XTP4可以使HepG2细胞凋亡率显著减少,而干扰XTP4的表达可以使HepG2细胞凋亡率显著增加。表明XTP4的高表达可以抑制HepG2细胞的凋亡,而下调XTP4的表达将加速HepG2细胞的凋亡。
Bcl- 2家族在凋亡的调控中起到关键作用,尤其是Bcl- 2/Bax比值被称作是启动细胞凋亡的分子开关[14]。在各种外界因素刺激下,Bcl- 2/Bax比值发生变化,通过影响凋亡执行分子胱冬肽酶- 3(caspase- 3),最终决定细胞的凋亡与否。通过Western blot检测过表达和干扰XTP4表达后Bcl- 2/Bax比值的变化情况,发现过表达XTP4后Bcl- 2/Bax比值增加,干扰XTP4的表达后Bcl- 2/Bax比值下降。进一步研究证实,过表达XTP4后,胱冬肽酶- 3的活性降低;干扰XTP4表达后,胱冬肽酶- 3的活性增加。表明XTP4的高表达可以上调Bcl- 2/Bax比值,降低胱冬肽酶- 3的活性,抑制HepG2细胞的凋亡,而下调XTP4的表达将下调Bcl- 2/Bax比值,增强胱冬肽酶- 3的活性加速凋亡。
综上所述,XTP4通过增加Bcl- 2/Bax比值,抑制HepG2细胞的凋亡,参与HBV致病过程。然而XTP4对Bcl- 2/Bax比值的调控作用通过哪些调控水平来实现及其确切的细胞内信号传导机制有待进一步研究。XTP4是否通过其他凋亡途径影响肿瘤细胞的凋亡,如死亡受体途径、内质网应激途径等,以及这些途径与上调Bcl- 2/Bax比值间是否存在相互作用有待进一步研究。
[1] Shepard CW,Simard EP,Finelli L,etal. Hepatitis B virus infection:epidemiology and vaccination[J].Epidemiol Rev,2006,28:112- 125.
[2] Cui Y, Jia J. Update on epidemiology of hepatitis B and C in China[J].J Gastroenterol Hepatol, 2013,28 Suppl 1:7- 10.
[3] Feitelson MA, Lee J. Hepatitis B virus integration, fragile sites, and hepatocarcinogenesis[J]. Cancer Lett,2007, 252:157- 170.
[4] Feitelson MA,Duan LX.Hepatitis B xirus X antigen in the pathogenesis of chronic infection and the development of hepatocellular carcinoma[J].AmJPathol,1997,150:1141- 1157.
[5] 刘妍,王春花,成军,等.乙型肝炎病毒X蛋白反式激活基因XTP4的克隆花研究[J].解放军医学杂志,2004:380- 382.
[6] Strausberg RL,Feingold EA,Grouse LH,etal.Generation and initial analysis of more than 15,000 full length human cDNA sequences[J].Proc Natl Acad Sci U S A,2002,99:16899- 16903.
[7] Evans EE,Henn AD,Jonason A,etal.C35(C17orf37) is a novel tumor biomarker abundantly expressed in breast cancer[J].Mol Cancer Ther,2006,5:2919- 2930.
[8] Benusiglio PR,Pharoah PD,Smith PL,etal.HapMap-based study of the 17q12 ERBB2 amplicon in susceptibility to breast cacer[J].Br J Cancer,2006,5:2919- 2930.
[9] Dasgupta S, Wasson LM, Rauniyar N,etal. Novel gene C17orf37 in 17q12 amplicon promotes migration and invasion of prostate cancer cells[J]. Oncogene,2009,28:2860- 2872.
[10] Leung TH, Wong SC, Chan KK,etal. The interaction between C35 and DeltaNp73 promotes chemo-resistance in ovarian cancer cells[J].Br J Cancer,2013,109:965- 975.
[11] Dasqupta S,Wasson LM,Rauniyar N,etal. Novel gene C17orf37 in 17q12 amplicon promotes migration and invasion of prostate cancer cells[J].Oncogene,2009,28:2860- 2872.
[12] 刘妍,成军,纪冬,等. 基因表达谱芯片技术筛选XTP4基因转染细胞差异表达基因[J]. 胃肠病学和肝病学杂志, 2004:209- 213.
[13] 韩萍,李莉,吴勤,等. 抑制性消减杂交技术筛选XTP4蛋白反式调节基因[J]. 解放军医学杂志, 2004:887- 888.
[14] Korsmeyer SJ, Shutter JR, Veis DJ,etal. Bcl- 2/Bax: a rheostat that regulates an anti-oxidant pathway and cell death[J]. Semin Cancer Biol,1993,4:4327- 332.
HBX protein trans-activate geneXTP4 suppresses HepG2 cells apoptosis
SHI Chao-hui1, ZHANG Meng-ran2, WANG Li-li1, LIU Shun-ai2, LIANG Pu2, WU Jun1, CHENG Jun2*, LIANG Yue-dong1*
(1.Guizhou Medical University,Guiyang 550004; 2.Institute of Infectious Diseases,Beijing Ditan Hospital, Capital Medical University,Beijing 100015,China)
Objective To explore the effects of HBX protein trans-activate gene XTP4 on cell apoptosis of HepG2 cells. Methods The plasmid pcDNA3.1/myc-His (-)A-XTP4 (pXTP4) was constructed,XTP4 gene interference RNA(siRNA) was chemically synthesized. And then negative control(NC,siNC) was transiently transfected into HepG2 cells respectively.The following experments were carried out post 48 hours: Western blot was used to for confirming the overexpression and interference expression of XTP4 protein.Flow cytometry was used to observe cell apoptosis.While Bcl- 2、Bax protein were semiquantified by Western blot,and then the ratio of Bcl- 2 to Bax was caculated. The activities of caspase- 3 were detected by caspase- glo 3/7 luminometer respectively.Results The plasmid pXTP4 was successfully constructed,XTP4 protein overexpressed and interferenced. Compared with control group respectively, the number of Annexin V-positive cells was decreased (P<0.05), the ratio of Bcl- 2 to Bax upregulated (P<0.05),the activity of caspase- 3 were reduced in pXTP4(P<0.05).While the number of Annexin V-positive cells was increased (P<0.05),the ratio of Bcl- 2 to Bax downregulated (P<0.05), the activity of caspase- 3 were increased in siXTP4(P<0.05). Conclusions The apoptosis of HepG2 was suppressed by XTP4,and the possible mechanism may be explained by the upregulate of Bcl- 2/Bax.
XTP4 gene; HepG2 cells; apoptosis
2015-07 - 01
2015- 10- 16
国家自然科学基金(81470863);北京市医院管理局重点医学专业发展计划(ZYLX201402)
1001-6325(2015)12-1591-05
R735.7
A
*通信作者(corresponding author):chengjdt@ccmu.edu.cn;lyd302@163.com
猜你喜欢
现代畜牧科技(2021年9期)2021-10-13 06:38:40
昆明医科大学学报(2021年5期)2021-07-22 07:31:58
中国生殖健康(2020年5期)2021-01-18 03:00:06
中国生殖健康(2018年5期)2018-11-06 07:15:56
食品与机械(2018年5期)2018-07-16 01:34:00
中国民族医药杂志(2016年5期)2016-05-09 07:43:50
天津医科大学学报(2015年3期)2015-06-05 12:21:49
新疆大学学报(自然科学版)(中英文)(2014年2期)2014-11-06 07:49:12
现代检验医学杂志(2014年2期)2014-02-02 02:40:34
中国医学科学院学报(2013年6期)2013-03-11 20:26:03
