
抑制MEK/ERK通路对孤独症模型大鼠病症行为的影响*
2015-03-14 08:37戴旭芳秦利燕
重庆医学 2015年36期
戴旭芳,秦利燕
(1.重庆师范大学教育科学学院特殊教育系,重庆400047;2.重庆市特殊儿童心理诊断与教育技术重点实验室,重庆400047;3.第三军医大学西南医院输血科,重庆400038)
孤独症(autism)又称儿童自闭症,是一种儿童脑发育障碍综合征,主要临床特征为交往障碍、语言障碍和(或)重复刻板行为等。发病率为2‰~6‰[1]。当前,中国约有60多万儿童患孤独症,且发病率呈上升趋势[2]。导致孤独症发病的原因复杂,涉及遗传、环境、教育等多种因素,目前尚未阐明[3]。
研究发现,编码 MEK/ERK(extracellular signal-regulated kinase)信号通路相关基因的缺失与孤独症的发病相关,如位于16号染色体上ERK1基因的缺失、22号染色体中ERK2基因或 MAPK(mitogen-aetivated protein kinase)基因的缺失均与孤独症的发病密切相关[4]。进一步研究表明,MEK/ERK信号通路可促进神经细胞的凋亡,而孤独症模型小鼠大脑皮质中 MEK/ERK信号通路过度激活[5],但抑制 MEK/ERK信号通路能否改善孤独症的行为症状目前尚未见报道。因此,本课题以孤独症大鼠为模型,探讨MEK/ERK信号通路的抑制剂U0126(l,4-diamino-2,3-dieyano-l,4-bis[2-aminophenylthio]butadiene)对孤独症大鼠病症的改善作用。
目前,科学家们已建立了多个孤独症动物模型,如病毒感染模型、基因敲除动物模型、VPA处理模型等[6]。在诸多模型中,VPA药物处理模型应用最广泛。因此,本研究主要以VPA药物处理的大鼠作为孤独症动物模型。
1 材料与方法
1.1 材料
1.1.1 实验动物 本实验中的Wistar大鼠购自第三军医大学大坪医院实验动物中心。雄性Wistar大鼠10只 (体质量300~350g),雌性 Wistar大鼠10只(体质量200~250g),于第三军医大学实验动物中心饲养。
1.1.2 药物与试剂 丙戊酸钠(sodium valproate,VPA)、U0126购自Sigma公司;GAPDH、MEK、磷酸化 MEK、ERK、磷酸化ERK抗体为Cell Signaling公司产品;PVDF膜为美国Bio-Rad公司产品;细胞裂解液和BCA蛋白浓度测定试剂盒为江苏碧云天生物技术公司产品;蛋白Marker及免疫组织化学染色试剂购自北京中杉金桥生物技术有限公司。
1.2 方法
1.2.1 动物模型制备 根据Schneider等[6-7]的方法,把雌、雄大鼠合笼,过夜后在雌鼠阴道发现阴栓者标记为妊娠第1天(E1)。所有模型组在E12.5d按600mg/kg剂量腹腔一次性注射VPA,对照组孕鼠腹腔注射等量的生理盐水。对照组孕鼠所产幼鼠作为对照组,而模型组孕鼠所产幼鼠作为孤独症模型组。U0126组及U0126联合VPA组从E13d开始按照每天400μg/kg体质量在饲料中添加U0126处理大鼠直至幼鼠断奶;幼鼠出生后第1天记为P1。
1.2.2 社会交往行为检测 出生后35d,从每组中各选择6只雄性幼鼠进行检测。各只检测鼠间的体重差别小于15g,且分笼饲养。检测场所为60cm×60cm×60cm的透明三室箱。检测前1d,将被检测鼠放入检测场所预适应环境。检测时,1只被测鼠首先被置于中间小室。10min后,在右侧小室放置陌生幼鼠并用铁丝笼子罩住,在左侧小室放置同样的空铁丝笼子。然后打开左、右小室与中间小室的通道,用摄像记录被检测鼠的行为,时间为10min。最后借助Any-Maze软件统计分析被检测鼠嗅铁丝笼子与陌生鼠(社交行为)时间及自梳理(非社交行为)时间。
1.2.3 神经行为学检测 在幼鼠出生后35d进行,被检测幼鼠的选择标准同上。分别从各组选择5只幼鼠进行检测,检测场所为25cm×25cm×38cm的封闭箱体,箱体中央区大小为12cm×12cm。检测前1d,将被检测鼠放入检测场所预适应环境。检测时,将被检测鼠置于检测场所中央,用摄像记录被检测鼠的行为,时长为5min。然后通过Any-Maze软件系统统计分析被检测鼠的直立次数及在中央区活动时间。
1.2.4 Western blot检测 在行为学检测完成之后,将幼鼠处死,并立即分离海马、前额叶皮层和小脑组织。把各组织用组织裂解液匀浆后于冰上放置30min,再以12 000r/min 4℃离心30min。分离上清液,测定蛋白浓度后放置于-80℃冰箱备用。实验时,把含50μg总蛋白的样品行SDS-聚丙烯酰胺凝胶电泳分离后转移至PVDF膜上。封闭后用一抗于4℃孵育过夜,再经PBST洗涤后加二抗室温孵育1h。最后经PBST洗涤后用化学发光液进行显影曝光。
1.3 统计学处理 所有数据采用SPSS 19.0统计软件分析,结果用x±s表示,两组间比较采用t检验,以P<0.05为差异有统计学意义。
2 结 果
2.1 U0126处理显著改善孤独症大鼠的社会交往能力 对各组大鼠的社会交往能力分析表明,与对照组相比,VPA组幼鼠社会交往能力明显下降,表现为不愿接触新事物,不喜欢与陌生幼鼠接触,自我梳理时间较长,与孤独症儿童的行为吻合。U0126处理组幼鼠的社会交往能力与正常对照组幼鼠比较差异无统计学意义(P>0.05)。而U0126联合VPA组幼鼠的社会交往能力与VPA组幼鼠相比则有显著改善(图1,表1)。

图1 各组大鼠的社会交往能力比较

表1 各处理组大鼠的社交活动时间统计分析结果(x±s,min,n=5)
2.2 U0126处理明显改善孤独症大鼠的焦虑行为
2.2.1 U0126处理减少孤独症幼鼠在中央区活动时间 行为检测结果表明,对照组幼鼠在中央区活动时间显著低于非中央区,而VPA处理组幼鼠在中央区与非中央区活动时间无显著差异,这一活动方式与正常对照组幼鼠的活动方式存在明显差异。U0126处理组幼鼠的活动方式与对照组幼鼠相比无显著差异;而U0126联合VPA处理组在中央区活动的时间则与VPA处理组相比显著减少,活动方式明显改变(图2、表2)。

图2 各组大鼠在中央区活动时间分析
2.2.2 U0126处理增加孤独症大鼠站立次数 检测结果表明,与正常对照组幼鼠相比,VPA处理组幼鼠在5min之内的站立次数显著降低,而VPA联合U0126处理组幼鼠的站立次数则比VPA处理组幼鼠明显增加。见图3、表3。
2.3 U0126处理抑制孤独症大鼠海马、前额叶皮层和小脑组织的MEK/ERK信号通路 Western blot结果表明(图4),与正常对照组相比,VPA处理组孤独症幼鼠的海马、前额叶皮层与小脑组织中MEK/ERK信号通路中MEK与ERK蛋白的磷酸化水平显著增强,而U0126处理组及VPA联合U0126处理组中MEK与ERK蛋白的磷酸化水平明显下降,表明U0126处理可抑制孤独症幼鼠前额叶皮层、海马和小脑组织中的MEK/ERK信号通路。

表2 各处理组大鼠的中央区活动时间统计分析结果(x±s,min,n=5)

图3 各组大鼠站立次数分析

表3 各处理组大鼠的站立时间统计分析结果(x±s,min,n=6)
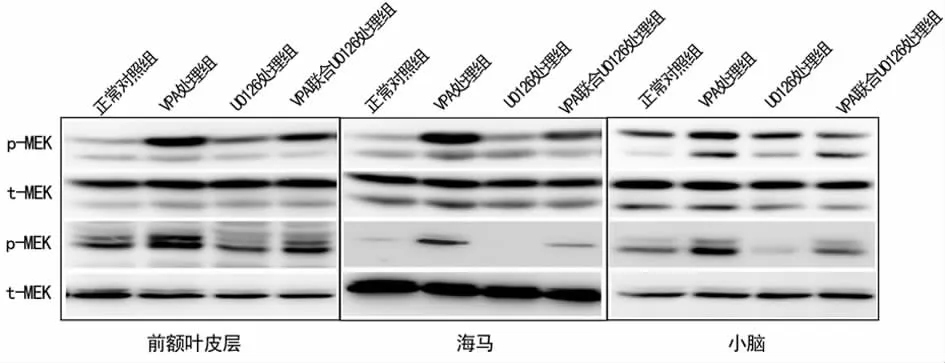
图4 Western blot分析各组大鼠前额叶皮层、海马和小脑组织中MEK和ERK表达水平
3 讨 论
社会交往实验是检测模型动物社会交往能力的有效实验,检测时,空笼子代表一种新事物,而陌生鼠代表新伙伴。通过分析被检测鼠与空笼子及陌生鼠的接触时间、自我梳理时间来判断其社会交往能力的强弱[8]。本研究中VPA组模型幼鼠的社会交往能力与对照组相比明显减弱,符合孤独症症状。在神经行为学检测实验中,被检测鼠在中央区所待时间的长短反映了其神经兴奋性,正常鼠具有避开中央区、在四周角落中探寻的习惯,而孤独症鼠则相对愿意待在中央区。站立次数则反映被检测鼠在新环境中的探索能力。正常鼠通常探索能力强、站立次数多,而孤独症鼠则探索能力减弱、站立次数减少[8]。本研究中,VPA组孤独症模型鼠的站立次数与对照组相比明显减少,表明其探索能力的减弱。VPA处理导致大鼠发生孤独症的相关机制尚不明确。本研究结果表明,VPA处理可以增加大鼠海马、前额叶与小脑组织中ERK与MEK蛋白的磷酸化,从而强化了MEK/ERK信号通路,这与Yin等[5]在BTBR孤独症模型小鼠大脑皮质中的发现相符,表明MEK/ERK信号通路的过度激活可能是孤独症发病的重要机制。
MEK与ERK是丝裂原化蛋白激酶家族(mitogen-aetivated protein kinases,MAPKs)成员,在神经前体细胞的生成、神经脊的发育和形成、突触信号传递以及意识、学习和记忆能力形成过程中发挥重要作用[9]。近年来,大量研究证实 MEK/ERK信号通路异常是导致智力发育异常的重要分子机制。敲除ERK基因后会导致神经干细胞的发育及成熟异常[10],而MEK/ERK信号通路的过度激活则可能导致神经细胞的凋亡[5]。本研究结果也进一步表明,孤独症模型大鼠脑组织中MEK/ERK通路被过度激活,而抑制这一信号通路则有助于改善孤独症的病症行为。
U0126 (l,4-diamino-2,3-dieyano-l,4-bis[2-aminophenylthio]butadiene)是一种高度选择性的MEK抑制剂,可特异地抑制 MEK/ERK通路活性[11]。U0126可阻断氨酸或喜树碱诱导的神经细胞损伤[12],还可减轻谷氨酰胺缺乏引起的神经细胞死亡及脑部外伤引起的海马神经损伤[13]。另外,多巴胺诱导的纹状神经元死亡也与 MEK/ERK通路活化有关[14]。但目前尚没有U0126改善孤独症症状的相关报道。本实验结果表明,U0126处理后大鼠前额叶皮质、海马及小脑组织中MEK与ERK蛋白磷酸化水平显著下降,MEK/ERK信号通路被抑制,模型大鼠的孤独症病症行为得到有效改善。因此,通过对MEK/ERK信号通路的抑制,U0126具有改善孤独症模型大鼠病症行为的作用,这一结果为孤独症的治疗提供了一定的理论与实验依据。
[1]Napoli E,Duefias N,Giulivi C.Potential therapeutic use of the ketogenic diet in autism spectrum disorders[J].Front Pediatr,2014,30(2):69.
[2]陈文雄.孤独症70年:从Kanner到DSM-V[J].临床儿科杂志,2013,31(11):1001-1004.
[3]Matson JL,Jang J.Treating aggression in persons with autism spectrum disorders:A review[J].Res Dev Disabil,2014,35(12):3386-3391.
[4]Pucilowska J,Puzerey PA,Karlo JC,et al.Disrupted ERK signaling during cortical development leads to abnormal progenitor proliferation,neuronal and network excitability and behavior,modeling human neuro-cardio-facial-cutaneous and related syndromes[J].J Neurosci,2012,32(25):8663-8677.
[5]Yin A,Qiu Y,Jia B,et al.Retracted:the developmental pattern of the RAS/RAF/Erk1/2pathway in the BTBR autism mouse model[J].Int J Dev Neurosci,2014,39:2-8.
[6]Schneider T,Przewlocki R.Behavioral alterations in rats prenatally exposed to valproic acid:animal model of autism[J].Neuropsychopharmacology,2005,30(1):80-89.
[7]Banerjee S,Riordan M,Bhat MA.Genetic aspects of autism spectrum disorders:insights from animal models[J].Front Cell Neurosci,2014,8:58.
[8]Shah CR,Forsberg CG,Kang JQ,et al.Letting a typical mouse judge whether mouse social interactions are atypical[J].Autism Res,2013,6(3):212-220.
[9]Tanimura S1,Nomura K,Ozaki K,et al.Prolonged nuclear retention of activated extracellular signal-regulated kinase 1/2is required for hepatocyte growth factor-induced cell motility[J].J Biol Chem,2002,277(31):28256-28264.
[10]Kalkman HO.Potential opposite roles of the extracellular signal-regulated kinase(ERK)pathway in autism spectrum and bipolar disorders[J].Neurosci Biobehav Rev,2012,36(10):2206-2213.
[11]Duan W,Wong WS.Targeting mitogen-activated protein kinases for asthma[J].Curr Drug Targets,2006,7(6):691-698.
[12]Lu KT,Cheng NC,Wu CY,et al.NKCC1-mediated traumatic brain injury-induced brain edema and neuron death via Raf/MEK/MAPK cascade[J].Crit Care Med,2008,36(3):917-922.
[13]de Bernardo S,Canals S,Casarejos MJ,et al.Role of extracellular signal-regulated protein kinase in neuronal cell death induced by glutathione depletion in neuron/glia mesencephalic cultures[J].J Neurochem,2004,91(3):667-682.
[14]Chen J,Rusnak M,Lombroso PJ,et al.Dopamine promotes striatal neuronal apoptotic death via ERK signaling cascades[J].Eur J Neurosci,2009,29(2):287-306.
猜你喜欢
海南医学院学报(2022年18期)2022-10-13
广东药科大学学报(2022年4期)2022-07-22
作文周刊·小学二年级版(2022年20期)2022-05-05
郑州大学学报(医学版)(2021年1期)2021-03-03
安徽医学(2020年12期)2020-02-03
创新作文(小学版)(2019年10期)2019-09-25
中国肿瘤外科杂志(2019年1期)2019-03-22
小学生学习指导(低年级)(2017年5期)2017-05-04
星星·散文诗(2016年23期)2016-12-29
中国实验诊断学(2014年10期)2014-06-01
