
表达鸡γ-干扰素重组鸡传染性支气管炎病毒的构建及鉴定
2015-03-09 05:40唐梦君沈欣悦李建梅尤素兰赵宝华戴亚斌
中国预防兽医学报 2015年3期
姜 逸,唐梦君,程 旭,沈欣悦,李建梅,尤素兰,赵宝华,戴亚斌,周 生
(中国农业科学院 家禽研究所,江苏 扬州 225003)
鸡传染性支气管炎病毒(Infectious bronchitis virus,IBV)为冠状病毒科冠状病毒属中的一个代表种,其基因组为不分节段的单股正链RNA,长约27.6 nt,具有典型的5' 帽子和3' Poly A 序列结构。IBV 至少有10 个明显的开放阅读框(ORF),ORF 的定位次序为5'-1a-1b-S-3a-3b-E-M-5a-5b-N-3'[1]。其中1a 和1b 编码病毒的RNA 聚合酶;S、E、M 和N编码病毒的结构蛋白;3a、3b、5a、5b 均编码病毒的非结构蛋白。在IBV 感染细胞内可以检测到3a、3b、5a 和5b,但其功能目前尚不清楚。研究表明3ab 和5ab 编码区的缺失不影响病毒的复制和致病性[2]。
γ-干扰素(IFN-γ)是由多种细胞由特定的诱生剂作用产生的一种具有高度生物活性的可溶性糖蛋白[3]。IFN-γ 可以促进Th1 细胞、CD8+T 细胞以及自然杀伤性细胞等细胞免疫应答,在抗病毒免疫及粘膜免疫中发挥重要作用[4]。鸡IFN-γ(ChIFN-γ)对多种病毒感染均具有较好的抑制作用,如流感病毒、新城疫病毒[5]以及马立克病病毒[6]等。ChIFN-γ 还能够作为一种有效的免疫佐剂[7]。ChIFN-γ 可以通过脂质体[8]、重组病毒载体[9]以及重组质粒DNA 体内接种发挥作用[10]。本研究前期已经建立了IBV H120 疫苗株的反向遗传操作系统[11],并且构建了表达gfp 基因的重组病毒H120-EGFP/5a[12]。在此基础上,本研究通过反向遗传技术构建表达IFN-γ 基因的重组IBV,为开展IBV 的分子致病机理,尤其是新型基因重组疫苗的研究奠定基础。
1 材料和方法
1.1 主要实验材料 用于构建IBV H120 疫苗株全长cDNA 的重组质粒系统pMDTM1~pMDTM10 及H120 株核蛋白基因(N)的真核表达质粒pVAXN 均由中国农业科学院家禽研究所保存;BHK-21 细胞株购自北京鼎国昌盛生物技术有限责任公司;SPF种蛋购自北京梅里亚维通实验动物技术有限公司。
1.2 主要试剂 PrimeSTAR®HS DNA 聚合酶、PrimeScript RT reagent Kit、DNA A-Tailing Kit、pMD18-T、SYBR Premix Ex Taq、Agarose Gel DNA Fragment Recovery Kit、T4 DNA 连接酶、T7 RNA聚合酶等均购自宝生物工程(大连)有限公司;限制性内切酶、m7G(5')ppp(5')G 帽子结构等购自NEB公司;GenJet 转染试剂购自美中清水湾生物科技有限公司;抗His 标签的单克隆抗体(MAb)购自生工生物工程(上海)有限公司;羊抗鼠IgG-HRP 购自南京生兴生物技术有限公司。
1.3 引物的设计及合成 根据GenBank 中登录的ChIFN-γ 基因序列(GQ246226)设计特异性扩增引物,cIFNF:5'-TTGGTCTCAGATGACTTGCCAGACTT-3'(Bsa Ⅰ)和cIFNR:5'-TTGGTCTCAGGTGGCAATTG CATCTCCT-3'(BsaⅠ),预扩增片段长度为515 bp。根据5a 基因所在重组质粒pMDTM9 的序列,设计用于构建pMDTM9-cIFN/5a 的引物,5aF:5'-TAGG TCTCGCACCATCACCATCACCATTAATGGCTGAC TAGTT-3'(BsaⅠ)和5aR:5'-GTTCTGGTCTCGCATC GTCTGTATTTGTTAAG-3'(BsaⅠ),并在ChIFN-γ 基因3' 端引入6×His 标签序列,预扩增片段长度约为5.8 kb。引物由上海生工生物工程技术服务有限公司合成。
1.4 ChIFN-γ 基因的扩增 鸡脾淋巴细胞的分离、诱导培养及总RNA 的提取参照文献[13]。按照反转录试剂盒使用说明合成cDNA,并以其为模板,cIFNF/cIFNR 为引物进行PCR 扩增。将PCR 产物经DNA A-Tailing Kit 按照使用说明处理后,连接至pMD18-T 构建重组质粒pMDcIFN,将其转化JM109,筛选阳性重组子,并进行测序鉴定。
1.5 重组质粒p MDTM9-c IFN/5a的构建 以重组质粒pMDTM9 为模板,5aF/5aR 为引物进行PCR 扩增。将PCR 产物及重组质粒pMDcIFN 分别采用BsaⅠ酶切后,胶回收目的片段并连接,构建重组质粒pMDTM9-cIFN/5a,将其转化JM109 进行测序鉴定。
1.6 表达ChIFN-γ 基因重组基因组全长cDNA的构建 将pMDTM1~pMDTM8、pMDTM9-cIFN/5a及pMDTM10 重组质粒经BsaⅠ酶切,胶回收TM1、TM2、TM3、TM4、TM5、TM6、TM7、TM8、TM9-cIFN/5a 和TM10 片段。根据文献[12]的方法构建表达ChIFN-γ 基因重组基因组全长cDNA。
1.7 重组病毒的拯救及鉴定 参考文献[12]的方法进行重组病毒的拯救。取P1 代含拯救病毒尿囊液,采用RNA 提取试剂盒提取其总RNA,反转录制备cDNA。采用分别位于重组病毒基因组中IFN-γ 基因上游和下游位置的检测引物JC4S(5'-GGAAAAGTCC ACTACGAAGG-3')和 JC4A(5'-GCTCTGCTTGTCCT GCTTTG),RT-PCR 扩增包含完整IFN-γ 基因的片段。拯救病毒的预扩增片段长度为968 bp,而rH120 的扩增片段长度为461 bp,并进行测序鉴定。将拯救的重组病毒命名为rH120-cIFNγ/5a。
1.8 目的基因表达的鉴定 取含有亲本病毒H120和拯救获得的rH120-cIFNγ/5a 尿囊液,经5 倍蛋白上样缓冲液煮沸8 min,制备蛋白样品进行SDSPAGE 电泳;电转至硝酸纤维膜上,采用5 %的脱脂奶封闭,以抗His 标签MAb 为一抗,羊抗鼠IgG-HRP 为二抗(1∶8 000),通过ECL 光学显影进行western blot 鉴定。
1.9 重组病毒的生物学特性测定 将rH120-cIFNγ/5a的P2 代以PBS 100 倍稀释后,接种于6 枚10 日龄的SPF 鸡胚尿囊腔内,0.2 mL/胚,同时2 枚未接种鸡胚作为阴性对照,置37 ℃孵化至144 h,观察鸡胚的病变;将病毒尿囊液经PBS 进行10 倍梯度稀释后,取10-5~10-9稀释度,分别接种5 枚10 日龄SPF 鸡胚,根据Reed-Muench 法测定EID50;将亲本病毒株H120 和rH120-cIFNγ/5a 的尿囊液104倍稀释后,接种于10 枚11 日龄的SPF 鸡胚,接种后间隔不同时间,使用注射器分别对每只胚进行连续取样,抽取尿囊液0.2 mL/胚,通过荧光定量RT-PCR法对样品中的病毒含量进行测定,绘制生长曲线。
1.10 重组病毒的遗传稳定性检测 将rH120-cIFNγ/5a P1 代尿囊液100 倍稀释后,接种于5 枚10 日龄~11 日龄的SPF 鸡胚中进行连续传代至P12 代后,取P1、P3、P6、P9 和P12 代病毒的尿囊液提取总RNA,反转录成cDNA,以JC4S/JC4A 为引物PCR 检测ChIFN-γ 基因的完整性。将P12 代扩增产物测序分析。
2 结果
2.1 ChIFN-γ 基因的扩增及IBV全长cDNA的构建 以白来航鸡脾淋巴细胞的诱导培养物的总RNA制备的cDNA 为模板,cIFNF/cIFNR 为引物,PCR扩增ChIFN-γ 基因,结果显示扩增片段长度约为500 bp,与预期结果相符(图1A)。测序结果表明该片段长度为515 bp,包含完整的ChIFN-γ 基因编码区。将ChIFN-γ 基因插入H120 基因组中非结构蛋白5a 编码区和转录调控序列(TRS)之间,构建含有ChIFN-γ 基因的IBV 基因组全长cDNA(图1B)。在ChIFN-γ 基因终止密码子前引入6×His 标签序列。

图1 A ChIFN-γ 基因的PCR 扩增Fig.1 A Amplification of ChIFN-γ gene by PCR

图1 B 表达ChIFN-γ 基因的rIBV 基因组全长cDNA 的构建Fig.1 B Construction of genomic cDNA of rIBV expressing ChIFN-γ gene
2.2 重组病毒的拯救及鉴定 以含有ChIFN-γ 基因的IBV 基因组全长cDNA 为模板,采用T7 RNA 聚合酶体外转录系统合成病毒基因组RNA,经脂质体转染BHK-21 细胞进行病毒的拯救。采用引物JC4S/JC4A,RT-PCR 扩增包含完整IFN-γ 基因的片段进行拯救的重组病毒鉴定。结果显示,重组病毒的扩增片段长度约1 kb,而H120 亲本病毒的扩增片段长度约0.5 kb,与预期结果相符(图2)。扩增产物的测序结果表明,ChIFN-γ 基因已正确插入到病毒基因组中5a 编码区的上游。

图2 重组病毒的RT-PCR 鉴定Fig.2 Identification of recombinant virus by RT-PCR
2.3 Western blot鉴定目的基因的表达 采用抗His 标签MAb 对rH120-cIFNγ/5a IFN-γ 的表达进行western blot 鉴定。结果显示,rH120-cIFNγ/5a 在约20 ku 处出现特异性条带,并且条带的大小与理论结果相符,而H120 则无此条带(图3)。

图3 Western blot 鉴定目的基因的表达Fig.3 Identification of ChIFN-γ expression by western blot
2.4 对鸡胚的致病性试验结果 将重组病毒接种10 日龄的SPF 鸡胚,至144 h 后观察鸡胚的病变;rH120-cIFNγ/5a 能够引起IBV 感染鸡胚的特征性变化:鸡胚发育受阻,成为侏儒胚(图4)。

图4 重组病毒感染鸡胚的病变Fig.4 Pathogenicity of rIBV to chicken embryos
2.5 重组病毒EID50的测定结果 将H120 和rH120-cIFNγ/5a 分别接种10 日龄SPF 鸡胚,根据Reed-Muench 法,测定H120 和rH120-cIFNγ/5a 的EID50,结果分别为107.5/0.1 mL 和106.83/0.1 mL。
2.6 重组病毒增殖曲线测定结果 将H120 和rH120-cIFNγ/5a 分别接种11 日龄SPF 鸡胚,通过荧光定量RT-PCR 对不同时间段取样中的病毒滴度进行测定。结果显示,两病毒株的病毒滴度均在感染后约30 h 达到峰值,两者配对样本t 检验结果(p<0.05)表明rH120-cIFNγ/5a 生长复制能力受到一定程度的影响(图5),这与EID50的测定结果相符。
2.7 重组病毒的遗传稳定性检测结果 将rH120-cIFNγ/5a 在SPF 鸡胚中进行连续传代,采用RT-PCR检测ChIFN-γ 基因在P1、P3、P6、P9 和P12 的完整性,结果显示,ChIFN-γ 基因在病毒传代至第9代时大部分仍保持完整,但至第12 代时开始出现明显的缺失(图6)。
进一步将P12 代重组病毒的扩增产物进行测序分析。结果显示,不同重组子中ChIFN-γ 基因发生缺失的位点及缺失的程度存在差异,部分重组病毒中ChIFN-γ 基因编码区已经完全缺失(图7)。
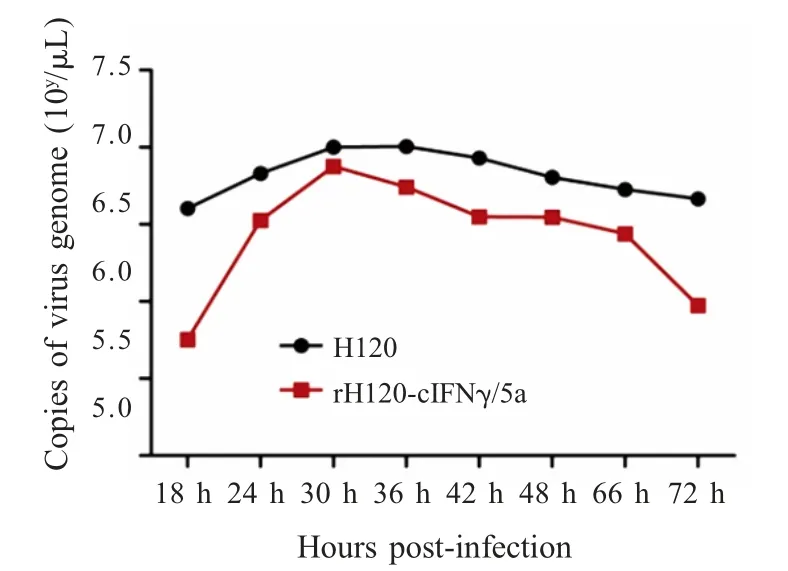
图5 H120 和rH120-cIFNγ/5a 的增殖曲线Fig.5 Growth curve of H120 and rH120-cIFNγ/5a

图6 重组病毒的遗传稳定性分析Fig.6 Analysis of genetic stability of the rIBV

图7 ChIFN-γ 基因缺失位点测序分析Fig.7 Integrity analysis of ChIFN-γ gene by sequencing
3 讨论
已有研究表明IBV 5a 蛋白编码区的缺失并不影响病毒的复制[3],因此本研究基于已建立的IBV H120 疫苗株的反向遗传操作系统,将ChIFN-γ 基因插入H120 病毒基因组中的5a 蛋白编码区上游,构建rH120-cIFNγ/5a。通过western blot 分析,在重组病毒感染鸡胚尿囊液中能够检测到ChIFN-γ 蛋白的表达。rH120-cIFNγ/5a 具有与亲本病毒株H120 相似的生物学特性,能够引起感染鸡胚特征性的变化,但与亲本病毒株相比,其毒价有所下降,因此ChIFN-γ 基因的插入对IBV 的复制产生了影响。
研究表明,不同外源基因在IBV 基因组中的稳定性不同,部分基因在传代过程中容易发生缺失。外源基因在重组冠状病毒中的稳定性受多种因素影响,如基因的大小、序列特性及其在病毒基因组中的位置等。Shen 等选择EGFP、萤火虫荧光素酶基因、eIF3f、SARS 病毒的ORF6 及登革热病毒核心蛋白基因,分别插入IBV 基因组的不同位置拯救获得重组病毒,并对重组病毒中外源基因的表达情况及重组病毒的遗传稳定性进行了研究,结果表明重组病毒的稳定性主要由外源基因本身的特性以及其在病毒基因组中的位置决定[14]。Bentley 等分别采用EGFP 和人源化海肾萤光素酶基因(hRluc)替换IBV基因组中的5a、IR 和3a3b 基因区域,结果也发现重组病毒的遗传稳定性与基因组片段插入的位置有关,并且表达密码子优化hRluc 基因的重组病毒,其遗传稳定性有所提高[15]。本研究将重组病毒rH120-cIFNγ/5a 株在SPF 鸡胚中进行连续传代,检测ChIFN-γ 基因在不同代次的完整性,结果表明ChIFN-γ 基因在病毒传代至第9 代时仍保持完整,但至第12 代时出现部分或完全缺失的现象。以上研究结果表明,研发以IBV 为载体的外源基因表达系统,需要解决重组病毒的遗传稳定性问题。
[1]Brian D A,Baric R S.Coronavirus genome structure and replication[J].Curr Top Microbiol Immunol,2005,287:1-30.
[2]Hodgson T,Britton P,Cavanagh D.Neither the RNA nor the proteins of open reading frames 3a and 3b of the coronavirus infectious bronchitis virus are essential for replication[J].J Virol,2006,80(1):296-305.
[3]Isaacs A,Lindenmann J.Virus interference.I.The interferon[J].Proc R Soc Lond B Biol Sci,1957,147(927):258-267.
[4]Boehm U,Klamp T,Groot M et al.Cellular responses to interferon-gamma[J].Annual Rev Immunol,1997,15:749-795.
[5]Yeh H Y,Winslow B J,Junker D E et al.In vitro effects of recombinant chicken interferon-gamma on immune cells[J].J Interferon Cytokine Res,1999,19(6):687-691.
[6]Zheng Xing,Schat K A.Inhibitory effects of nitric oxide and gamma interferon on in vitro and in vivo replication of Marek's disease virus[J].J Virol,2000,74(8):3605-3612.
[7]Lowenthal J W,O'Neil T E,Broadway M,et al.Coadministration of IFN-gamma enhances antibody responses in chickens[J].J Interferon Cytokine Res,1998,18(8):617-622.
[8]Kedar E,Palgi O,Golod G et al.Delivery of cytokines by liposomes.III.Liposome-encapsulated GM-CSF and TNF-alpha show improved pharmacokinetics and biological activity and reduced toxicity in mice[J].J Immunother,1997,20(3):180-193.
[9]Johnson M A,Pooley C,Lowenthal J W.Delivery of avian cytokines by adenovirus vectors[J].Dev Comp Immunol,2000,24(2-3):343-354.
[10]Haygreen L,Davison F,Kaiser P.DNA vaccines for poultry:the jump from theory to practice[J].Expert Rev Vaccines,2005,4(1):51-62.
[11]周生,戴亚斌,唐梦君,等.鸡传染性支气管炎病毒H120疫苗株全长cDNA 感染性克隆的构建[J].中国家禽,2010,23:22-26.
[12]周生,唐梦君,戴亚斌,等.表达绿色荧光蛋白的重组鸡传染性支气管炎病毒的构建[J].病毒学报,2011,01:11-17.
[13]刘胜旺,刘思国,卢景良.鸡干扰素基因的分子克隆与序列测定[J].中国免疫学杂志,2001,01:45-47.
[14]Shen Hong-yuan,Fang Shou-guo,Chen Bo,et al.Towards construction of viral vectors based on avian coronavirus infectious bronchitis virus for gene delivery and vaccine development[J].J Virol Methods,2009,160(1-2):48-56.
[15]Bentley K,Armesto M,Britton P.Infectious bronchitis virus as a vector for the expression of heterologous genes[J].PLoS One,2013,8(6):e67875.
猜你喜欢
成都医学院学报(2022年4期)2022-08-19
江西农业学报(2021年4期)2021-04-20
中国兽医杂志(2020年4期)2020-09-28
三农资讯半月报(2020年11期)2020-06-21
养猪(2020年1期)2020-02-19
今日农业(2019年11期)2019-08-15
国外畜牧学·猪与禽(2018年11期)2018-05-14
中国介入影像与治疗学(2017年12期)2017-01-14
农村农业农民·B版(2015年9期)2015-10-16
中国当代医药(2015年9期)2015-03-01
