
不同灌溉方式对马铃薯光合特性的影响
2015-02-23 02:44王雯,张雄
安康学院学报 2015年4期
王 雯,张 雄
(榆林学院 生命科学学院,陕西 榆林 719000)
马铃属为茄科茄属一年生草本块茎植物,具有耐寒、耐贫瘠、适应性强、生长周期短、营养成分全等特点,联合国粮农组织已将马铃薯列为世界第四大主粮。据报道,2015年中国将启动马铃薯主粮化发展战略,力争通过几年时间,使马铃薯的种植面积、产量及主粮化产品在粮食产品总消费量中的比重显著提升。榆林沙区光照充足,土质疏松,排灌方便,是全国马铃薯五大优生区和高产区之一,该地区常年马铃薯种植面积达17.3万公顷左右,约占粮食作物种植总面积的1/3,是全国马铃薯生产第三大市。毋庸置疑,马铃薯主粮化战略将为榆林沙区马铃薯产业的健康快速发展提供新的契机。
榆林沙区属典型的西北干旱地区,农业水资源严重短缺,且利用效率低下,而马铃薯整个生育期需水较多且对水分变化较为敏感,水分亏缺是导致马铃薯产量不稳和品质下降的主要因素[1]。同时,光合作用是影响马铃薯产量和品质的生理基础,也是马铃薯高产优质的重要因素,已有研究表明,马铃薯叶片的净光合速率、蒸腾速率以及气孔导度均随水分胁迫程度增加而降低[2-3]。然而,不同灌溉方式对马铃薯光合作用有何影响?马铃薯光响应曲线及其参数值有何变化?截止到目前,相关研究报道尚较为少见。鉴于此,我们开展了不同灌溉方式对马铃薯光合特性影响的研究。这项研究对于探明不同灌溉方式下马铃薯的光合生理特性及其调控机制,制定科学合理的马铃薯节水灌溉制度,实现马铃薯的高产、稳产,都具有重要意义。
1 材料与方法
1.1 试验地概况
试验地位于榆林现代农业科技示范区,地处榆林市牛家梁镇榆卜界村 (109°43′E,38°23′N),该区域属干旱半干旱大陆性季风气候,年平均降水量371mm,蒸发量1900mm,年日照时数2900 h,年总辐射量606.7×107J·m2,年均气温8.6℃,10℃积温为3000~3300℃,无霜期167 d。该区域光照充沛,地势平坦,地下水位较高,便于灌溉,土壤为风沙土,肥力水平中等。供试土壤PH值为8.1,有机质含量为7.85g·kg-1,全氮含量为0.36g·kg-1,碱解氮含量为 48.90mg·kg-1,有效磷含量为 13.95mg·kg-1,速效钾含量为 87mg·kg-1。
1.2 试验设计
试验设漫灌(CK)、沟灌(GA,每条沟均匀灌水)、交替隔沟灌(JG,上次灌水沟下次不灌而邻沟灌)、露地滴灌(DG)和膜下滴灌(MG)5个处理。小区面积3m×7m,随机区组排列,重复3次,为了避免不同处理间的相互影响,每个处理间和同一处理间都相隔1m。MG种植模式为1膜1带1行,施底肥后起垄覆膜播种,垄底宽60 cm,垄顶宽40 cm,垄间距40 cm,垄高15 cm,马铃薯行距为60+40 cm,株距30 cm,垄起好后,铺设滴灌带(采用内镶贴片式滴灌带,滴头间距30 cm,滴头流量1.38 L/h),播种后用幅宽80 cm,厚度0.008mm的地膜覆盖;DG种植模式为1带1行不覆膜;GA和JG种植模式为起垄覆膜不铺设滴灌带;CK种植模式为不起垄不覆膜不铺设滴灌带,DG、GA、JG和CK的株、行距与MG相同。漫灌的灌溉量和灌溉次数按照当地农户传统灌溉方式确定。膜下滴灌、露地滴灌、交替隔沟灌和沟灌每次灌溉量分别为漫灌的25%、25%、44%和56%,每次灌溉时,用水表测定各小区的容积灌溉量(m3)。在整个生育期,膜下滴灌、露地滴灌、交替隔沟灌、沟灌和漫灌的灌溉次数分别为11、11、8、7和6次,各处理的灌溉量见表1。
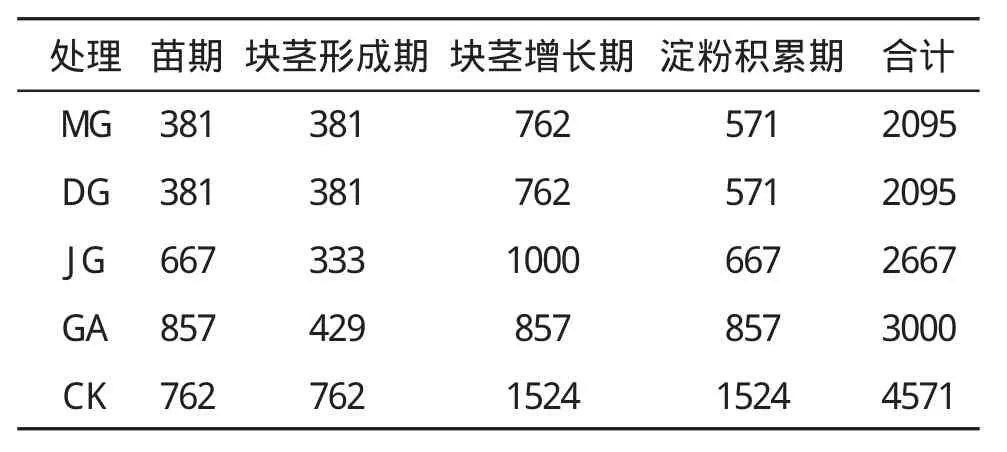
表1 不同灌溉方式的灌溉量(单位:m3·hm-2)
于2014年5月25日播种并展开田间管理试验,供试品种为当地主栽品种紫花白,播种深度8~10cm,66750株·hm-2,底肥施尿素 120 kg·hm-2,磷酸二铵 225 kg·hm-2,硫酸钾 225 kg·hm-2,混匀后在耕地时撒施地表,深翻入土壤。在块茎形成至块茎增长期随灌水追肥3次,滴灌处理每次50 kg·hm-2,其他处理每次 150 kg·hm-2,其余田间管理方式相同,9月28日收获。
1.3 测定项目及方法
试验测定指标包括土壤含水量、光合指标及光响应曲线等。指标具体测量方法如下:
(1)土壤含水量。采用土钻取样烘干法测定20、40、60、80、100 cm土层的含水量。
(2)光合指标。采用LI-6400便携式光合测定仪 (LI-COR,Inc,Lincoln,NE,USA),分别于2014年7月4日、7月26日和8月24日光照充足且相对稳定的时间段9:00~11:00测定光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等光合指标。每小区分别选取3株长势相同的马铃薯,在自然光照下测定其倒3叶的顶小叶的光合指标,每株重复测定3次,结果取其平均值。
(3)光响应曲线。采用LI-6400光合测定仪测定进行不同灌溉方式的马铃薯叶片光响应曲线(Pn-PAR曲线)。测定时,将LI-6400光合测定仪的气体流速设定为500μmol·s-1,参比CO2浓度设置为400μmol·mol-1,叶室温度设定为30℃,将光合有效辐射(PAR)强度设定为2000、1800、1600、1400、1200、1000、800、600、400、200、100、60、20、10、0μmol·m-2·s-1共 15个梯度,每个梯度下平衡时间为120s[4-5],每小区选择3株,根据测定结果重复1~2次。
1.4 数据处理
采用Ye等[6]提出的直角双曲线修正模型(式(1))对光响应曲线进行拟合,并根据模型估算最大净光合速率、表观光量子效率、暗呼吸速率、光饱和点、光补偿点等光合参数。
式(1)中,I为光合有效辐射(μmol·m-2·s-1),α为表观光量子效率(即光响应曲线上I=0时的斜率),β和γ为修正系数(m2·s·μmol-1),Ic为光补偿点(μmol·m-2·s-1)。
式(2)中,Isat为作物叶片的光饱和点光强(μmol·m-2·s-1)。
式(3)中,Rd为作物的暗呼吸速率(μmol·m-2·s-1)。
式(4)中,Pmax为最大净光合速率。
运用SPSS19.0软件进行光响应曲线拟合、光合参数计算及单因素方差分析(LSD,p<0.05),采用Origin 8.5软件绘图。
2 结果与分析
2.1 不同灌溉方式对马铃薯光合作用的影响
2.1.1 马铃薯光合指标
由图1可见,在马铃薯生长旺盛期,不同处理的马铃薯叶片光合指标均表现为MG>DG>JG>GA>CK,且MG处理显著高于CK。在块茎形成期、块茎增长期和淀粉积累期,经MG处理的马铃薯叶片净光合速率分别较CK高65.2%、64.6%和63.8%,蒸腾速率分别较CK高35.0%、18.3%和24.3%,气孔导度分别较CK高45.5%、46.8%和38.6%,胞间CO2浓度分别较CK高40.5%、32.0%、37.9%,增幅最大。并且,MG处理的马铃薯叶片净光合速率较DG高5.1%~18.1%,蒸腾速率较DG高0.8%~7.8%,气孔导度较DG高4.5%~12.0%,胞间CO2浓度较DG高3.8%~8.8%。本研究结果表明,膜下滴灌处理的马铃薯植株光合能力均高于其他灌溉方式。
2.1.2 Pn-Ci以及Pn-Gs的相互关系
回归分析结果显示(见图2),在马铃薯生长旺盛期,各处理的净光合速率与胞间CO2浓度(Pn-Ci)之间以及净光合速率与气孔导度(Pn-Gs)之间均表现为显著的线性正相关关系(R2>0.89,p<0.05)。在块茎增长期,Pn-Ci的相关性最高(p=0.006,R2=0.9411)。在块茎形成期,Pn-Gs的相关性最高(p=0.001,R2=0.9879)。
2.2 不同灌溉方式下马铃薯光响应曲线比较
2.2.1 光响应曲线
由图3可见,在马铃薯生长旺盛期,不同灌溉方式下的净光合速率在0~200μmol·m-2·s-1光强下呈直线上升趋势,当光强超过400μmol·m-2·s-1时,其上升速度明显减缓,MG处理的马铃薯叶片净光合速率在1400μmol·m-2·s-1光强处达到最大值(块茎形成期、块茎增长期和淀粉积累期分别为22.92,26.24,24.36μmol·m-2·s-1),而DG、JG、GA和CK处理的马铃薯叶片分别在1200、1200、1000、1000μmol·m-2·s-1光强处达到最大值,此后随着光强继续增加,不同处理的净光合速率均出现下降,其中MG处理的马铃薯叶片净光合速率下降幅度低于其他处理(MG、DG、JG、GA和CK处理的降幅分别为7.2%~8.6%、13.5%~17.8%、15.3%~24.9%、18.0%~21.3%和21.4%~27.4%)。
2.2.2 光响应曲线参数值
利用直角双曲线修正模型计算的光响应曲线参数结果显示,在马铃薯生长旺盛期,不同处理的表观光量子效率(α)、最大净光合速率(Pmax)、光饱和点(Isat)均表现为MG>DG>JG>GA>CK(见表2)。在各生育期,MG处理的α值显著高于GA和CK(p<0.05),MG处理的Pmax值均显著高于JG、GA和CK(p<0.05),在淀粉积累期,MG处理的α值和Pmax值分别较CK增加60.5%和70.1%,增幅最大。在各生育期,MG处理的Isat值均显著高于其他处理(p<0.05),在块茎形成期,MG处理的Isat值较CK增加40.3%,增幅最大。而不同处理的光补偿点(Ic)和暗呼吸速率(Rd)的变化趋势却与前三个参数相反,均表现为MG 表2 不同灌溉条件下马铃薯光响应曲线参数值 本研究表明,在马铃薯生长旺盛期,膜下滴灌处理的马铃薯叶片Pn值、Gs值、Tr值和Ci值均高于其他处理,且显著高于漫灌(图1)。尽管露地滴灌的灌量、灌次和膜下滴灌相同,但是露地滴灌处理的马铃薯叶片光合指标仍低于膜下滴灌。已有研究资料显示,与传统的漫灌和沟灌相比,膜下滴灌具有明显优势。一方面覆膜种植不仅可降低土壤水分蒸发,减少株间蒸发和养分损耗,其保墒效果显著[3,7];另一方面,滴灌能够按照作物生长和需水规律,实施适时、适量的精确灌溉,使作物根区土壤水分含量适宜、土壤物理结构良好,植株间保持适宜的温度和湿度,有效避免在夏季炎热天气中出现的高温伤薯现象,因而马铃薯植株长势稳健,能够保持较高的光合性能。 作物净光合速率下降主要有两方面因素:其一是气孔因素限制,即叶片气孔导度下降,CO2吸收速率减小,导致胞间CO2浓度降低;二是非气孔因素限制,即叶肉细胞光合作用强度下降,致使叶肉细胞对CO2的利用能力减弱,导致胞间CO2浓度降低[8-9]。因此,分析作物净光合速率变化的原因要综合考虑气孔导度和胞间CO2浓度两方面因素[10]。当作物净光合速率下降时,如果胞间CO2浓度和气孔导度同时下降,说明净光合速率的降低主要是由于气孔因素限制;如果气孔导度下降,而胞间CO2浓度反而上升,则表明净光合速率下降的主要原因是非气孔因素限制,即叶肉细胞光合能力降低[11]。本研究中,各处理的Pn-Ci以及Pn-Gs均表现显著的正相关关系(R2>0.89,p<0.05)(图2),这表明JG、GA和CK处理的马铃薯叶片净光合速率显著低于MG(p<0.05)(图1)的主要原因是其气孔导度下降引起的胞间CO2浓度降低,应归因于气孔因素限制。这是由于在马铃薯生长旺盛期,JG、GA和CK处理的土壤含水量为田间持水量的50%~64%,处于轻度水分胁迫,经JG、GA和CK处理的马铃薯叶肉细胞的光合机构虽未受到损伤,但植株仍需通过降低叶片的气孔导度来防止植株过度蒸腾失水,这一过程也导致叶片净光合速率下降。而膜下滴灌处理的土壤含水量为田间持水量的67%~74%,适宜的土壤含水量能使气孔导度保持较高水平,蒸腾速率增加有利于植株内水分运输和光合产物的合成,从而使净光合速率也明显增加。这表明在马铃薯生长盛期,膜下滴灌保持适宜的土壤含水量,能使马铃薯光合能力明显增强。 表观光量子效率(α)是衡量植物对弱光的利用能力的重要指标。相关研究显示,对于长势良好的作物,α值一般在0.040~0.070之间[12-14]。本研究中膜下滴灌处理的α值在0.062~0.076之间,均高于其他处理,并显著高于沟灌和漫灌处理(表2),这表明膜下滴灌处理的马铃薯对弱光的利用能力较强(p<0.05)。光饱和点和光的高低是衡量作物对强光和弱光利用能力的重要指标[15]。在马铃薯生长旺盛期,不同处理的光饱和点和光补偿点的变化范围分别为991.516~1436.604和26.901~53.251μmol·m-2·s-1(表 2),这与何长征等[2]、凌永胜等[16]以及黄承建等[17]的研究结果基本一致。膜下滴灌处理的光饱和点均显著高于其他处理(p<0.05),其光补偿点也高于其他处理,且显著高于漫灌(p<0.05)(表2),这说明与其他处理相比,膜下滴灌处理使马铃薯对光环境的适应能力明显增强。暗呼吸速率高低是反映作物光合产物消耗大小的重要指标[18-19]。与其他处理相比,膜下滴灌处理的最大净光合速率最高而暗呼吸速率最低,表明该处理的光合产物消耗较低(表2)。在淀粉积累期,膜下滴灌处理的最大净光合速率较漫灌增加60.5%,而其暗呼吸速率较漫灌减小44.9%,表明膜下滴灌处理能促进光合产物向块茎运转,有利于块茎增长和淀粉积累。此外,本研究利用直角双曲线修正模型计算的最大净光合速率(表2)和实际测得的最大净光合速率(图3)相差较小(变化幅度为0.7%~4.7%),这表明利用直角双曲线修正模型能较好地反映不同灌溉方式下马铃薯的光合特性的变化。 在马铃薯生长旺盛期,膜下滴灌处理的马铃薯叶片的Pn、Tr、Gs和Ci值均高于其他处理,且显著高于漫灌(p<0.05);各处理的Pn-Ci以及Pn-Gs均表现为显著的正相关关系(R2>0.89,p<0.05);膜下滴灌处理的α、Pmax、Isat值均高于其他处理,且显著高于沟灌和漫灌(p<0.05)。而其Ic和Rd值均低于其他处理,且Ic值显著低于漫灌(p<0.05)。因此在榆林沙区,膜下滴灌处理的马铃薯光合特性优于其他灌溉方式。 [1]王婷,海梅荣,罗海琴,等.水分胁迫对马铃薯光合生理特性和产量的影响[J].云南农业大学学报,2010,25(5):737-742. [2]何长征,刘明月,宋勇,等.马铃薯叶片光合特性研究[J].湖南农业大学学报:自然科学版,2005,31(5):518-520. [3]张恒嘉,李晶.绿洲膜下滴灌调亏马铃薯光合生理特性与水分利用[J].农业机械学报,2013,44(10):143-151. [4]YEZIPIAO,YUQIANG,KANGHUA JING.Evaluation of photosynthetic electron flow using simultaneous measurements of gas exchange and chlorophyll fluorescence under photorespiratory conditions[J].Photosynthetica,2012(50):472-476. [5]YE ZIPIAO.Nonlinear optical absorption of photosynthetic pigment molecules in leaves[J].Photosynthesis Research,2012,112:31-37. [6]YE ZIPIAO,DAVID JS,PIOTR R,etal.Amechanistic model for the photosynthesis–light response based on the photosynthetic electron transport of photosystem II in C3and C4species[J].New Phytologist,2013,199(1):110-120. [7]王志军,谢宗铭,田又升,等.膜下滴灌水稻黄化叶片光合特性及叶绿素荧光参数分析[J].西北农业学报,2015,24(25):59-65. [8]FARQUHAR G D,SHARKEY TD.Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33(6):317-345. [9]THOMASD S.Photosynthesis in intact leaves of C3plants:Physics,physiology and rate limitations[J].The Botanical Review,1985,55(1):53-105. [10]景茂,曹福亮,汪贵斌,等.土壤水分含量对银杏光合特性的影响[J].南京林业大学学报:自然科学版,2005,29(4):83-86. [11]陈根云,陈娟,许大全.关于净光合速率和胞间CO2浓度关系的思考[J].植物生理学通讯,2010,46(1):64-66. [12]XUCHAO FAN,YIN YONGGAO,CAIRUIGUO,etal.Responses of photosynthetic characteristics and antioxidative metabolism in winterwheat to post-anthesis shading[J].Photosynthetica,2013,51(1):139-150. [13]郎莹,张光灿,张征坤,等.不同土壤水分下山杏光合作用光响应过程及其模拟[J].生态学报,2011,31(16):4499-4508. [14]焦念元,宁堂原,杨萌珂,等.玉米花生间作对玉米光合特性及产量形成的影响[J].生态学报,2013,33(14):4324-4330. [15]梁娟,郭泽宇,叶漪.不同土壤水分条件对七叶一枝花光合特性及有效成分皂苷含量的影响[J].植物生理学报,2014,50(1):56-60. [16]凌永胜,李锦泉,林金秀,等.马铃薯泉云3号和泉引1号叶片光合特性研究[J].福建农业学报,2014,23(3):209-213. [17]黄承建,赵思毅,王龙昌,等.马铃薯/玉米套作对马铃薯品种光合特性及产量的影响[J].作物学报,2013,39(2):330-342. [18]康华靖,李红,权伟.四种作物光下暗呼吸速率降低的原因[J].植物生态学报,2014,38(10):1110-1116. [19]王智明,张峰举,许兴,等.3种油葵光合光响应曲线模型拟合及其光合参数与产量的关系[J].江苏农业学报,2014,30(3):491-496.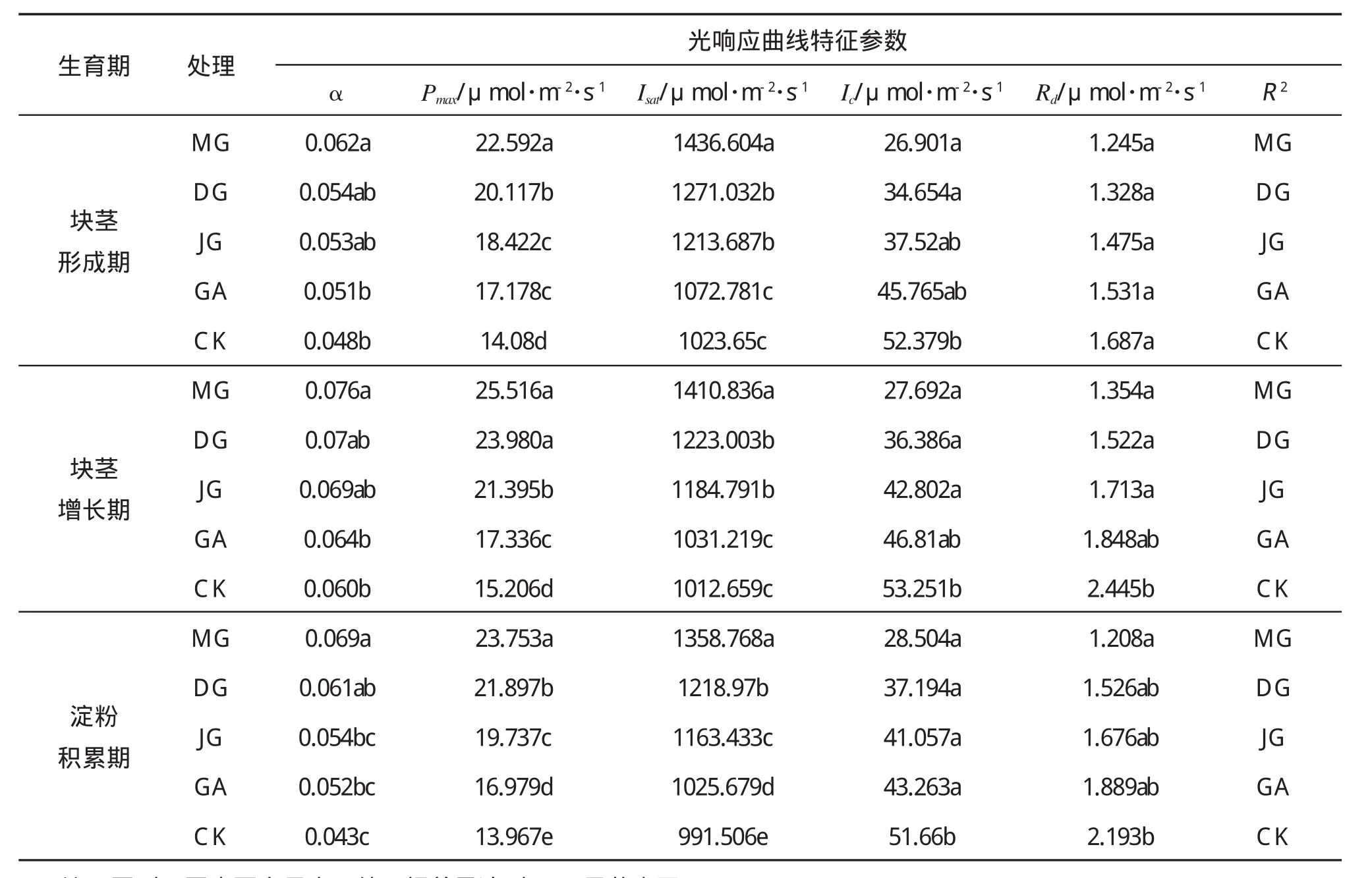
3 讨论
4 结论
猜你喜欢
生态学报(2022年18期)2022-10-13
航空材料学报(2022年4期)2022-08-04
东北林业大学学报(2022年4期)2022-05-16
贵州农业科学(2022年4期)2022-05-06
农业工程学报(2022年4期)2022-04-24
农业工程学报(2022年3期)2022-04-16
干旱地区农业研究(2022年1期)2022-01-28
新疆农业科技(2021年2期)2021-07-03
农业与技术(2020年12期)2020-07-04
科技创新与品牌(2019年12期)2019-02-06
